
The essential role of architectural noncoding RNA Neat1 in cold-induced beige adipocyte differentiation in mice
- Hikaru Toya1,
- Yuko Okamatsu-Ogura2,
- Saori Yokoi1,
- Misuzu Kurihara1,
- Mari Mito3,
- Shintaro Iwasaki3,
- Tetsuro Hirose4 and
- Shinichi Nakagawa1
- 1RNA Biology Laboratory, Faculty of Pharmaceutical Sciences, Hokkaido University, Sapporo 060-0812, Japan
- 2Laboratory of Biochemistry, Faculty of Veterinary Medicine, Hokkaido University, Sapporo 060-0818, Japan
- 3RNA Systems Biochemistry Laboratory, RIKEN Cluster for Pioneering Research, Saitama 351-0198, Japan
- 4RNA Biofunction Laboratory, Graduate School of Frontier Biosciences, Osaka University, Suita 565-0871, Japan
- Corresponding author: nakagawas{at}pharm.hokudai.ac.jp
Abstract
Neat1 is an architectural RNA that provides the structural basis for nuclear bodies known as paraspeckles. Although the assembly processes by which Neat1 organizes paraspeckle components are well-documented, the physiological functions of Neat1 are not yet fully understood. This is partly because Neat1 knockout (KO) mice, lacking paraspeckles, do not exhibit overt phenotypes under normal laboratory conditions. During our search for conditions that elicit clear phenotypes in Neat1 KO mice, we discovered that the differentiation of beige adipocytes—inducible thermogenic cells that emerge upon cold exposure—is severely impaired in these mutant mice. Neat1_2, the architectural isoform of Neat1, is transiently upregulated during the early stages of beige adipocyte differentiation, coinciding with increased paraspeckle formation. Genes with altered expression during beige adipocyte differentiation typically cluster at specific chromosomal locations, some of which move closer to paraspeckles upon cold exposure. These observations suggest that paraspeckles might coordinate the regulation of these gene clusters by controlling the activity of certain transcriptional condensates that coregulate multiple genes. We propose that our findings highlight a potential role for Neat1 and paraspeckles in modulating chromosomal organization and gene expression, potentially crucial processes for the differentiation of beige adipocytes.
Keywords
- Neat1
- paraspeckles
- cold exposure
- beige adipocytes
- knockout mice
- nondomain biopolymer
- intrinsically disordered regions
INTRODUCTION
In the nuclei of higher eukaryotes, biomolecules such as DNA, RNA, and proteins are not uniformly distributed but are organized into distinct compartments known as nuclear bodies (Hirose et al. 2023; Shan et al. 2023). These nuclear bodies, often referred to as nonmembranous organelles or biomolecular condensates, consist of components that engage in specific molecular processes, typically associating through weak multivalent interactions to form flexible and dynamic structures (Roden and Gladfelter 2021; Uversky 2021). One such mammalian-specific nuclear body, the paraspeckle, was initially identified as a structure enriched with particular RNA-binding proteins (Fox et al. 2002). Subsequent studies have revealed that these structures form along a long noncoding RNA (lncRNA) known as Neat1 (nuclear paraspeckle assembly transcript 1), which serves as a critical architectural component within these bodies (Chen and Carmichael 2009; Clemson et al. 2009; Sasaki et al. 2009; Sunwoo et al. 2009). To date, more than 40 proteins, many harboring intrinsically disordered regions (IDRs), have been identified as constituents of paraspeckles (Naganuma et al. 2012; Fong et al. 2013; Fox et al. 2018). Proteins with IDRs in paraspeckles can undergo phase transitions in vitro, forming reversible hydrogels or liquid droplets, providing a molecular basis for the formation of paraspeckles (Hennig et al. 2015; Fox et al. 2018). Arranged along specific regions of Neat1, these proteins contribute to the characteristic core–shell structure of paraspeckles (Souquere et al. 2010; West et al. 2016). The Neat1 gene produces two isoforms, Neat1_1 and Neat1_2, through differential 3′-end processing from the same promoter (Sasaki et al. 2009), with only the longer isoform, Neat1_2, capable of inducing paraspeckle formation (Sasaki et al. 2009; Naganuma et al. 2012). Neat1_2 contains multiple functional domains involved in various processes, including transcript stabilization, isoform switching, and assembly (Yamazaki et al. 2018). The modular organization of Neat1_2 imparts distinct features to paraspeckles, such as a defined diameter, an internal core–shell structure, and a tendency to merge into elongated, sausage-like formations, explainable by the block copolymer micelles model (Yamazaki et al. 2021).
Although the role of Neat1 in paraspeckle formation is well-established, its physiological functions and the molecular mechanisms involved remain less clear (Fox et al. 2018; Nakagawa et al. 2022). Previous studies have reported that Neat1 knockout (KO) female mice show decreased fertility because of dysfunctional corpus luteum formation, responsible for progesterone production (Nakagawa et al. 2014). This phenotype is not consistently observed across all animals and is present in roughly half of them. Intriguingly, the same homozygous KO mice that have experienced successful pregnancies may subsequently encounter pregnancy failures, suggesting that the low-penetrance phenotype is not attributable to genetic background variations (Nakagawa et al. 2014). Neat1 KO mice display several other phenotypes, including impaired mammary duct formation (Standaert et al. 2014), reduced proliferation of vascular smooth muscle cells postinjury (Ahmed et al. 2018), compromised adaptive behavioral responses to stress (Kukharsky et al. 2020), and impaired response of macrophages to LPS (Azam et al. 2024), among others (Gao et al. 2020). Generally, Neat1 KO mice present little to no overt phenotypes under normal conditions; however, these phenotypes become pronounced when subjected to specific environmental stressors. Additionally, Neat1’s function in promoting or inhibiting cancer progression varies with the experimental model (Adriaens et al. 2016; Mello et al. 2017), supporting the notion that its role is context-dependent.
In response to cold stimulation, mice, like many other mammals, use thermogenic strategies to maintain their body temperature, relying on both shivering and nonshivering thermogenesis. Brown adipose tissue (BAT) plays a central role in nonshivering thermogenesis, characterized by brown adipocytes that contain multilocular lipid droplets and a high mitochondrial density. These mitochondria, through the action of uncoupling protein 1 (Ucp1), dissipate the proton gradient, converting energy typically used for ATP synthesis into heat, thereby playing a crucial role in thermoregulation (Nicholls 2017; Chouchani et al. 2019). In addition to the well-characterized thermogenic role of BAT, another thermogenic cell type, known as beige adipocytes, emerges in response to cold exposure. These cells, induced within subsets of white adipose tissue such as inguinal white adipose tissue (iWAT), share characteristics with brown adipocytes. However, unlike brown adipocytes, beige adipocytes are rarely present in normal mice reared under normal ambient temperature (23°C) but are instead conditionally induced upon cold exposure (Young et al. 1984; Wu et al. 2012). Once exposed to a cold environment, the sensation of cold by the central nervous system triggers the activation of the sympathetic nervous system, which in turn activates BAT by inducing the expression of Ucp1 (Wang and Seale 2016). The activation of sympathetic nerves also induces the differentiation of beige cells in white adipose tissues, including iWAT, contributing to the body's adaptive nonshivering thermogenesis mechanism alongside BAT (Wang and Seale 2016).
To further investigate Neat1’s physiological roles and identify conditions that elicit clear, high-penetrance phenotypes, we subjected the animals to various physiological stressors. We found that Neat1 KO mice exhibited impaired beige adipocyte differentiation in response to cold exposure. During early differentiation, Neat1_2 is transiently upregulated in adipocytes or adipocyte precursor cells, coinciding with enhanced paraspeckle formation in these cells. RNA-seq analysis of Neat1 KO mice indicated that the expression of multiple genes, differentially expressed during beige adipocyte differentiation, was altered in the mutant mice. Notably, these genes were found to frequently cluster at specific genomic loci, which are repositioned closer to paraspeckles in the nucleus upon cold exposure. These spatial rearrangements of target loci suggest a potential regulatory interaction, indicating that paraspeckles might influence certain transcriptional condensates that regulate the expression of these gene clusters during the differentiation of beige adipocytes.
RESULTS
The differentiation of beige adipocytes upon cold exposure is severely impaired in Neat1 KO mice
To explore the physiological environments or conditions where Neat1 becomes essential, we subjected Neat1 KO mice to several stress conditions, including glucose tolerance, starvation, high-fat diet, and cold exposure. Among these, we discovered that cold exposure–induced beige adipocyte differentiation was severely impaired in the mutant mice (Fig. 1). Consequently, we focused on this cell type in subsequent studies. Beige adipocytes are conditional thermogenic cells that emerge upon cold exposure in specific adipose tissues such as iWAT (Young et al. 1984; Wu et al. 2012), contributing to nonshivering thermogenesis alongside constitutive thermogenic cells known as brown adipocytes residing in BAT. To investigate the differentiation of beige adipocytes, we exposed the animals to a cold environment and examined the histology of iWAT through hematoxylin and eosin (H&E) staining on paraffin sections. Under normal ambient temperature (23°C), iWAT primarily consisted of white adipocytes, identified by single cytoplasmic lipid droplets and a small nucleus in both wild-type (WT) and KO mice (Fig. 1A). Upon cold exposure (4°C for 1 week), differentiation of beige adipocytes was induced in the iWAT of WT mice, identified by the formation of small multilocular lipid droplets in the cytoplasm (Fig. 1A). Conversely, beige adipocyte differentiation was severely impaired in Neat1 KO mice, with iWAT remaining predominantly composed of white adipocytes even after cold exposure, although we occasionally observed small patches of cells resembling beige adipocytes (arrowheads in Fig. 1A). To further validate the impaired differentiation of beige adipocytes in the iWAT of Neat1 KO mice, we investigated the expression of Ucp1, a marker protein for beige adipocytes known to uncouple oxidative phosphorylation from ATP production in mitochondria, thereby generating heat (Nicholls 2017; Chouchani et al. 2019). Immunohistochemical analyses on paraffin sections indicated that Ucp1+ beige adipocytes were greatly reduced in the iWAT of Neat1 KO mice compared to their WT counterparts (Fig. 1B), which was also confirmed by western blot analysis (Fig. 1C). We also assessed the mRNA expression of genes essential for the differentiation and/or function of beige adipocytes using quantitative RT-PCR (qRT-PCR), including Elovl3, Cidea, Cox4, Pgc1a, and Eva1. All these genes were upregulated in the iWAT of WT mice upon cold exposure, a response absent in Neat1 KO mice (Fig. 1D).
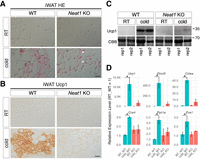
Impaired beige adipocyte differentiation in Neat1 KO mice. (A) Nomarski images of H&E-stained paraffin sections of iWAT derived from WT and Neat1 KO mice reared under normal ambient temperature (RT) and cold temperature (cold) for 1 week. Beige cells are identifiable by numerous small multilocular lipid droplets in the cytoplasm forming mesh-like structures. (B) Immunohistochemical detection of Ucp1 in iWAT derived from WT and Neat1 KO mice reared under normal (RT) and cold temperature (cold) for 1 week. Note that Ucp1 expression is strongly upregulated in WT iWAT upon cold exposure, which is absent in Neat1 KO mice. (C) Western blot analyses of Ucp1 expression in iWAT of WT and Neat1 KO animals. Two biological replicates (rep1 and rep2) were used for each condition. Coomassie brilliant blue (CBB)-stained gels served as a loading control. (D) Expression of mRNAs for beige cell markers Ucp1, Elovl3, Cidea, Cox4, Pgc1a, and Eva1. Note that these markers were upregulated upon cold exposure in WT mice, which is attenuated in Neat1 KO mice. The asterisks indicate statistically significant changes: (*) P < 0.05, bootstrap test, 1000 surrogates, n = 2 for RT and n = 3 for cold. Scale bar, 20 µm.
Brown adipocytes are normally formed in Neat1 KO mice
Given that beige and brown adipocytes share most signaling pathways and transcription factors required for their differentiation (Shapira and Seale 2019), we investigated whether brown adipocyte differentiation was also impaired in Neat1 KO mice. Unexpectedly, we found normal brown adipocytes in the BAT of Neat1 KO mice reared at room temperature, as confirmed by histological H&E staining on paraffin sections (Fig. 2A), Ucp1 immunohistochemistry (Fig. 2B), and western blotting (Fig. 2C). Activation of brown adipocytes upon cold exposure was also noted in Neat1 KO mice, as evidenced by the increased expression of Ucp1 and mRNA expression of marker genes (Fig. 2C,D). These results indicate that the sensation of cold by the central nervous system and the subsequent activation of sympathetic nerves occur normally in Neat1 KO mice, and that Neat1 specifically regulates the differentiation of beige adipocytes, rather than that of brown adipocytes, upon cold exposure.
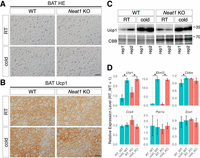
Brown adipocyte differentiation appears fairly normal in Neat1 KO mice. (A) Nomarski images of H&E-stained paraffin sections of BAT derived from WT and Neat1 KO mice reared under normal ambient temperature (RT) and cold temperature (cold) for 1 week. No apparent differences were observed between WT and Neat1 KO BAT. (B) Immunohistochemical detection of Ucp1 in BAT derived from WT and Neat1 KO mice reared under normal (RT) and cold temperature (cold) for 1 week. Note the presence of Ucp1 expression in both conditions. (C) Western blot analyses of Ucp1 expression in BAT of WT and Neat1 KO animals. Two biological replicates (rep1 and rep2) were used for each condition. CBB-stained gels served as a loading control. Scale bar, 20 µm. (D) Expression of mRNAs for beige cell markers Ucp1, Elovl3, Cidea, Cox4, Pgc1a, and Eva1. The asterisks indicate statistically significant changes: (*) P < 0.05, bootstrap test, 1000 surrogates, n = 2 for RT and n = 3 for cold.
Neat1_2 is transiently upregulated during the early stages of beige cell differentiation
To further investigate the role of Neat1 and paraspeckle formation in beige adipocyte differentiation, we evaluated Neat1 expression in iWAT at various time points—0 h, 4 h, 24 h, and 4 days—after cold exposure using a probe that detects both Neat1_1 and Neat1_2 via northern blot analysis (Fig. 3A). As the probe is known to cross-react with unidentified transcripts similar in size to Neat1_2 (asterisk in Fig. 3B; Isobe et al. 2020), we also used a probe that specifically detects Neat1_2 (Fig. 3A). Under standard conditions at room temperature (0 h), robust expression of Neat1_1 was observed in all examined animals, whereas the expression of Neat1_2 varied: Two animals exhibited only weak signals of Neat1_2, and one displayed a detectable, albeit low, level of the long isoform (Fig. 3B). These findings suggest that Neat1_2 expression is typically suppressed in iWAT at room temperature but can be induced by certain physiological stimuli. Upon cold exposure, a marked increase in Neat1_2 was noted in all three animals 4 h poststimulation, followed by a decrease at subsequent time points such as 24 h and 4 days. In contrast, Neat1_1 levels remained relatively constant across all instances (Fig. 3B). The robust induction of Neat1_2 upon cold exposure was not observed in BAT (Fig. 3B). We further confirmed the transient upregulation of Neat1_2 4 h after cold exposure through qRT-PCR (Fig. 3C). Notably, the surge in Neat1_2 precedes the elevation of beige adipocyte marker Ucp1 (Fig. 3C), suggesting that Neat1_2 may regulate the early stages of beige adipocyte differentiation upon cold exposure, well before the emergence of morphologically distinct, functional beige adipocytes.
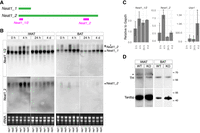
Transient upregulation of Neat1_2 during early stages of beige adipocyte differentiation upon cold exposure. (A) Schematic representation of the positions of the probes that detect both Neat1_1 and Neat1_2 (Neat1_1/2) and solely Neat1_2. (B) Northern blot analyses of Neat1_1 and Neat1_2 expression in iWAT and BAT at 0 h, 4 h, 24 h, and 4 days after cold exposure. rRNA was used as a loading control, visualized using ethidium bromide–stained agarose gels. The asterisk indicates nonspecific signals detected by the Neat1_1/2 probe. (C) qRT-PCR analyses of Neat1_1/2, Neat1_2, and Ucp1 expression in iWATs at various time points post–cold exposure. Note the earlier upregulation of Neat1_2 compared to that of Ucp1. The asterisks indicate statistically significant changes: (*) P < 0.05, bootstrap test compared with “0 h” followed by post hoc Bonferroni correction, 1000 surrogates, n = 3 per group. (D) Western blot analyses of tyrosine hydroxylase (TH) expression in sympathetic ganglia of WT and Neat1 KO (KO) mice. Tardbp was used as a loading control.
Beige adipocyte differentiation is triggered by sympathetic nerve stimulation upon cold exposure. Consequently, we assessed whether sympathetic innervation in iWATs was altered in Neat1 KO mice by evaluating the levels of TH, a recognized marker for sympathetic nerves, using western blot analysis. The TH levels in iWATs of both WT and KO mice were comparable (Fig. 3D), indicating that nerve innervation into iWAT was unchanged in Neat1 KO mice.
Paraspeckle formation is enhanced during the initial stages of beige adipocyte differentiation
To investigate the expression of Neat1 and the formation of paraspeckles in iWAT at the cellular level, we performed in situ hybridization of Neat1 using probes that detect both isoforms and a probe specific to Neat1_2. In control iWAT, Neat1_2 expression was usually observed as small spots, likely representing transcription sites, whereas Neat1_1 signals, identifiable only with the Neat1_1/2 probes, were more diffusely distributed within the nucleoplasm (Fig. 4A). These patterns are reminiscent of previously reported microspeckles, which form nonparaspeckle structures in the absence of Neat1_2 expression (Li et al. 2017). With cold exposure, Neat1_2 signals intensified and formed distinct spots that fully coincided with the Neat1_1/2 probe signals (Fig. 4A). This indicates that Neat1_2 upregulation triggered the formation of discrete paraspeckles during cold exposure, and Neat1_1 transcripts, typically forming microspeckles, were incorporated into paraspeckles. To quantify these observations, we counted the number of Neat1_1/2 and Neat1_2 signals before and after cold exposure and measured the area of these signals in each nucleus. The count of foci detected with Neat1_1/2, most likely representing microspeckles, significantly dropped after cold exposure, accompanied by an increase in focus areas (Fig. 4B). Conversely, the number of foci detected with Neat1_2, corresponding to paraspeckles, increased (Fig. 4B). The areas positive for Neat1_2 also expanded, although the change was not statistically significant. This could be attributed to the formation of paraspeckles in a limited cell population, induced by the stochastic stress exerted on the cells. To further validate that the Neat1_2 foci indeed are paraspeckles, we simultaneously detected Neat1_2 and the established paraspeckle marker protein Nono (Fig. 4C). Previous studies have reported that Nono is diffusely distributed in the nucleoplasm and enriched in paraspeckles in both cultured cell lines and mouse tissues (Fox et al. 2005; Nakagawa et al. 2011). In iWAT, Nono signals were not detected in the nucleoplasm, likely because of the presence of large lipid droplets that hinder the penetration of fixative during sample preparation. However, Nono signals within paraspeckles were clearly observed 4 h after cold stimulation, confirming the augmented formation of paraspeckles upon cold exposure (Fig. 4C).
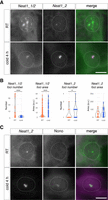
Formation of enlarged paraspeckles in iWAT upon cold exposure. (A) Subnuclear localization of Neat1_1 and Neat1_2 in iWAT. Localization of Neat1_1 becomes restricted to paraspeckles where Neat1_2 accumulates upon cold exposure. (B) Quantification of the number of foci per cell and the area of each focus positive for Neat1_1/2 or Neat1_2. The asterisks indicate statistically significant differences: (*) P < 0.05, (***) P < 0.001, Mann–Whitney U-test. (C) Simultaneous detection of Neat1_2 and the paraspeckle marker protein Nono in adipocytes of iWAT upon cold exposure. Scale bar, 5 µm. Dotted lines represent the shape of each nucleus.
The long isoform of Neat1_2, but not the short isoform Neat1_1, is required for beige adipocyte differentiation
Although the short isoform Neat1_1 is dispensable for paraspeckle formation (Sasaki et al. 2009; Naganuma et al. 2012), it has been suggested to play a role independent of paraspeckles by forming microspeckles, which are diffusely distributed in the nucleoplasm (Li et al. 2017). Because the Neat1 KO mice used in our study lack the expression of both isoforms, we were unable to determine which Neat1 isoform is essential for the formation of beige adipocytes. To address this, we examined Neat1_1-specific KO mice (Neat1 ΔPAS), which lack the polyadenylation signals necessary for the production of Neat1_1 (Isobe et al. 2020). Beige adipocytes were normally induced in the iWAT of Neat1 ΔPAS mice following cold exposure, as evidenced by histochemical analysis and marker gene expression (Fig. 5A–D). These findings suggest that the loss of Neat1_2, rather than Neat1_1, accounts for the impaired differentiation of beige adipocytes upon cold exposure observed in Neat1 KO mice.
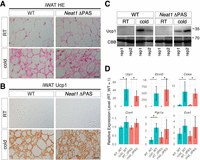
Beige adipocyte differentiation appears normal in Neat1 ΔPAS Mice. (A) Nomarski images of H&E-stained paraffin sections of iWAT derived from WT and Neat1 ΔPAS mice reared under normal ambient temperature (RT) and cold temperature (cold) for 1 week. (B) Immunohistochemical detection of Ucp1 in iWAT derived from WT and Neat1 ΔPAS mice reared under normal (RT) and cold temperature (cold) for 1 week. (C) Western blot analyses of Ucp1 expression in iWAT of WT and Neat1 ΔPAS animals. Two biological replicates (rep1 and rep2) were used for each condition. CBB-stained gels served as a loading control. (D) Expression of mRNAs for beige cell markers Ucp1, Elovl3, Cidea, Cox4, Pgc1a, and Eva1. The asterisks indicate statistically significant changes: (*) P < 0.05, bootstrap test, 1000 surrogates, n = 3 per group. Scale bar, 20 µm.
Neat1 is expressed in a subpopulation of white adipocytes and adipocyte precursor cells
Although iWAT is mostly composed of white adipocytes, it also contains adipocyte precursor cells, endothelial cells, and lymphocytes such as macrophages, T cells, and B cells. To gain insight into the specific cell types expressing Neat1, we reanalyzed public single-cell RNA-seq data sets obtained from iWAT of adult mice 10 weeks old, maintained at room temperature (Fig. 6A; Rajbhandari et al. 2019). Given that the RNA-seq libraries were constructed using oligo-dT primers, only the expression of polyadenylated Neat1_1, but not the RNase P-cleaved Neat1_2, was captured in the data set. Nonetheless, we observed cell type–biased expression of Neat1, with the highest levels in white adipocytes, followed by adipocyte precursor cells (Fig. 6A). Other cell types, including endothelial cells and lymphocytes—macrophages, T cells, and B cells—showed lower levels of Neat1 expression (Fig. 6A). To verify the cell types expressing Neat1, we performed simultaneous in situ hybridization to detect Neat1 and pre-mRNA of a white adipocyte marker Pparg (Tontonoz and Spiegelman 2008) and an adipocyte precursor marker Pi16 (Merrick et al. 2019). The pre-mRNA signals, identified using probes that target intronic sequences, were observed at transcription sites within the nucleus. This allowed for the identification of nuclei specific to these cell types (Fig. 6B). Upon cold exposure, signals for Neat1_1/2 were detected in half of the Pparg-expressing adipocytes (12 out of 22 cells, or 54.5%) and in a smaller subset of the Pi16-expressing adipocyte precursor cells (2 out of 22 cells, or 9%), suggesting that upregulation of Neat1 occurs in a subset of white adipocytes and adipocyte precursors at a defined time point (Fig. 6B).
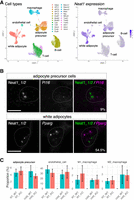
Neat1 expression in subpopulations of white adipocytes and adipocyte precursors in iWAT. (A) UMAP visualization of adult iWAT cell types and associated Neat1 expression from reanalyzed public data sets (GSE133486). Colors denote distinct cell types, including white adipocytes (Adipo), adipocyte precursors (APC1–4), B cells (Bcell1–3), T cells (Cd4 and Cd8), macrophages (Macro1–3), and endothelial cells (Endo), illustrating that Neat1 expression is prevalent among subpopulations of white adipocytes and adipocyte precursor cells. (B) Simultaneous detection of Neat1 and pre-mRNAs of cell type–specific markers using intron-targeted probes. White adipocytes and adipocyte precursor cells were identified by the expression of Pparg and Pi16, respectively. The panels indicate the percentages of these cells expressing Neat1. Scale bar, 5 µm. (C) Proportions of adipocyte precursors, endothelial cells, and M1/M2 macrophages in iWAT of WT and Neat1 KO mice, with no statistically significant differences due to KO of Neat1 observed (Mann–Whitney U-test, n = 6 per group).
The cellular composition of iWAT was largely unaltered in Neat1 KO mice
The differentiation of beige adipocytes might be explained by either a deficiency of beige precursor cells or a reduced capacity for differentiation following cold exposure. Beige adipocytes are known to arise from both adipocyte precursor cells and morphologically identifiable white adipocytes (Shao et al. 2019). Given the absence of noticeable changes in the morphology or number of white adipocytes in the iWAT of Neat1 KO mice (Fig. 1A), we focused on adipocyte precursor cells identifiable by the surface antigen markers CD31−/Sca1+/CD34+/PDGFRα+ (Shin et al. 2017). No statistically significant differences were observed in the counts of adipocyte precursor cells either before or after cold exposure (Fig. 6C). We also assessed the numbers of endothelial cells and macrophages, as these cells secrete factors known to influence the differentiation of beige cells in iWATs (Seki et al. 2016; Wang and Seale 2016; Villarroya et al. 2018). FACS analysis showed that CD31+ endothelial cells were unaffected in the iWAT of Neat1 KO mice (Fig. 6C; Supplemental Fig. S1). Similarly, the numbers of F4/80+/CD11c+ M1 macrophages and F4/80+/CD206+ M2 macrophages, which, respectively, inhibit and promote the differentiation of beige adipocytes, were not altered (Fig. 6C; Supplemental Fig. S1). These results suggest that the cellular composition of iWATs does not change in Neat1 KO mice, both under normal conditions and during the early stages of cold exposure.
Neat1-regulated genes form clusters across multiple genomic loci
To gain further insights into the molecular mechanisms involved, we performed RNA-seq analysis on iWAT derived from WT and Neat1 KO mice at 0, 4, and 24 h after cold exposure, and the count data (Supplemental Tables S1 and S2) were analyzed using RNAseqChef (Etoh and Nakao 2023). Initial analysis using UMAP and hierarchical clustering revealed a significant overlap in the transcriptional profiles among samples collected at 0 and 4 h, regardless of genotype (KO or WT). This suggests that individual biological variability predominates over genotypic or temporal differences during the early time points upon cold exposure (Supplemental Fig. S2A). In contrast, samples collected at 24 h posttreatment exhibited a clear distinction between KO and WT groups (Supplemental Fig. S2A). This divergence was also evident in volcano plot analyses, in which most genes showed similar expression levels at room temperature, but these levels diverged over time post–cold exposure (Fig. 7A). Enrichment analyses following K-means clustering revealed pathways upregulated in WT but not in KO mice 24 h after cold exposure, including oxidative phosphorylation, adipogenesis, cholesterol homeostasis, and fatty acid metabolism. These are consistent with the impaired beige cell differentiation observed in Neat1 KO mice (Supplemental Fig. S2B,C). Pathways specifically upregulated in Neat1 KO mice included myogenesis and the G2M checkpoint, although the physiological significance of these findings remains to be clarified (Supplemental Fig. S2B,C). We then used the iDEP-Genome algorithm (Ge et al. 2018), which scans the genome with 6 Mbp sliding windows and 2 Mbp overlaps, to identify regions exhibiting statistically significant changes in gene expression. Remarkably, genes that changed expression 24 h post–cold exposure in Neat1 KO mice clustered on multiple chromosomes, including the proximal region of Chromosome 19 containing the Neat1 gene (Fig. 7B). Furthermore, some genomic regions enriched in downregulated genes in Neat1 KO mice overlapped with regions enriched for genes upregulated during beige adipocyte differentiation, identified by comparing expression data from WT iWAT at 0 and 24 h post–cold exposure (Fig. 7B,C). This suggests that gene clusters are coregulated during beige adipocyte differentiation, a process that is partially impaired in Neat1 KO mice lacking paraspeckles. To determine if the positions of these gene clusters relative to paraspeckles change upon cold exposure, we simultaneously detected paraspeckles and the gene loci within differentially expressed clusters (Fig. 7D) and measured the proximity of differentially expressed gene loci to the nearest paraspeckles (Fig. 7E). At room temperature, the distances between gene loci and paraspeckles were not significantly different compared to an unrelated control locus 4.5SH, except for genes in the cluster containing Neat1 (Fig. 7E). However, upon cold exposure, five out of six examined gene loci (Tfr2, Gamt, Uqcr11, Gpt, and Fasn) were closer to paraspeckles than control loci 4.5SH (Fig. 7E). The average distance between paraspeckles and these gene loci, which shortened upon cold exposure, was 1.46 ± 0.92 µm, less than the distance to control loci 4.5SH (2.76 ± 1.03 µm) but more than that to genes in the Neat1-containing cluster (0.60 ± 0.56) (Fig. 7E). These findings suggest that although paraspeckles do not directly associate with clusters of differentially expressed genes, they may recruit these genes to specific submicrometer-scale, transcriptionally active compartments that form in the areas surrounding the paraspeckles.
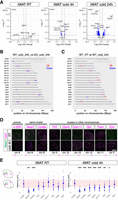
Clustering of differentially expressed genes in iWAT of Neat1 KO mice at specific genomic regions. (A) Volcano plot illustrating differentially expressed genes between WT and Neat1 KO mice at room temperature, and at 4 and 24 h after cold exposure. Blue dots represent genes with statistically significant changes (FDR < 0.05, log2 fold change >1) as calculated by DESeq2. Neat1 showed an exceptionally low log10 (FDR) and log2 fold change, which is force adjusted to 12.0 and 5.0, respectively, in the plots for visualization purposes. (B) Identification of genomic regions enriched in differentially expressed genes in Neat1 KO mice using the “Genome” function of iDEP. The genome was scanned with 6 Mbp windows and 2 Mbp overlap; blue and red bars with end caps indicate regions where average fold changes are statistically higher (red) or lower (blue) compared to other regions. Magenta rectangles highlight genomic regions enriched in genes downregulated in Neat1 KO mice but upregulated upon cold exposure in WT mice. (C) Genomic regions enriched in genes differentially expressed during beige adipocyte differentiation in iWAT. Magenta rectangles again highlight regions with genes downregulated in Neat1 KO mice but upregulated upon cold exposure in WT mice. (D) Simultaneous detection of paraspeckles and genomic loci of clustered genes downregulated in Neat1 KO mice. Paraspeckles were identified by Neat1 expression, and gene loci were detected by DNA FISH. The chromosomal positions of each gene are indicated below with magenta lines. (E) Statistical analysis of 3D distances between genomic loci shown in D and paraspeckles in iWAT at room temperature and 4 h after cold exposure. The shortest distance (d) between paraspeckles and each genomic locus was measured. The asterisks denote gene loci that are significantly closer to paraspeckles compared to the unrelated control gene locus 4.5SH: (***) P < 0.001, (**) P < 0.01, (*) P < 0.05, Dunnett's multiple comparisons test compared with 4.5SH. Pink rectangles indicate the range of distances between paraspeckles and unrelated gene loci (mean ± SD). Dotted lines represent the shape of each nucleus. Scale bar, 5 µm.
DISCUSSION
We have shown that Neat1 KO mice display a marked reduction in beige adipocyte formation when subjected to cold conditions. Although the precise molecular mechanisms leading to aberrant gene expression in the iWAT of Neat1 KO mice remain elusive, we have observed that genes differentially expressed in these mice cluster within specific genomic regions. Previous research has established that genes involved in particular biological pathways or lineage specification tend to be coregulated and form clusters at specific genomic loci (Lee and Sonnhammer 2003; Kosak et al. 2007; Neems et al. 2016; Zinani et al. 2022). These coexpression domains are distinct from topologically associated domains (TADs) (Soler-Oliva et al. 2017), which are more stable across different cell types (Eres and Gilad 2021). Advanced live imaging analyses of transcriptional activity at the single-cell level have revealed that transcription factor clusters can coactivate multiple promoters across a given genomic region (Kawasaki and Fukaya 2023). Notably, the formation of these transcription factor clusters is enhanced by the artificial incorporation of IDRs into trans-acting transcription factors (Kawasaki and Fukaya 2023). The functional significance of IDRs in transcriptional regulation, potentially promoting the assembly of molecular condensates that govern the expression of coregulated genes, has garnered considerable interest (Staby et al. 2017; Tsafou et al. 2018; Brodsky et al. 2020). Given that paraspeckles harbor various proteins equipped with distinct IDRs (Hennig et al. 2015) and that Neat1 associates with transcriptionally active chromosomal loci (West et al. 2014), it is plausible that Neat1 may modulate the formation of such molecular condensates by recruiting IDR-containing proteins, thereby enhancing the formation of a micrometer-scale transcriptionally active area surrounding paraspeckles (Fig. 8). This could account for our observation that differentially expressed genes move toward paraspeckles upon cold exposure. Recent research has also shown that Neat1 is acutely and transiently induced upon LPS stimulation in macrophages, required for the expression of inflammatory cytokines (Azam et al. 2024). In this scenario, transient upregulation of Neat1 and concomitant enlargement of paraspeckles lead to the redistribution of multiple paraspeckle proteins containing distinct IDRs (Azam et al. 2024). Identifying IDR-containing proteins that link paraspeckles with transcriptional regulation in iWAT is crucial to substantiating this hypothesis. Furthermore, examining if gene cluster repositioning is impaired in Neat1 KO mice is essential, either by comparing each locus's position with the adjacent Neat1 gene locus to paraspeckles or through genome-wide chromatin conformation analyses using techniques like Hi-C. However, given that Neat1 is expressed only in a subset of Pparg-expressing white adipocytes and Pi16-expressing adipocyte precursors, such experiments are technically unfeasible because of the lack of exclusive molecular markers necessary to identify cells expected to express Neat1 and undergo rearrangement. In essence, Neat1 serves as a marker for cells poised to express it. However, in Neat1 KO mice, where Neat1 expression is absent, these specific cells cannot be identified.
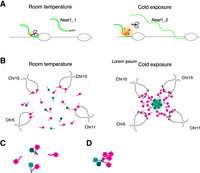
Model of cold-induced formation of paraspeckles and beige adipocyte differentiation. (A) Model for Neat1_2 production upon cold exposure in white adipose tissue. At room temperature, components that induce readthrough of polyadenylation signals (light and dark orange) cannot form a functional assembly, resulting in the production of the short, polyadenylated Neat1_1 isoform. Cold exposure triggers a phase transition in these factors, leading to the readthrough of polyadenylation signals and the production of the long Neat1_2 isoform. (B) Model for the cold-induced movement of gene clusters upregulated during beige cell differentiation. Cold exposure leads to Neat1_2 production and paraspeckle formation, which may initiate the formation of a nuclear compartment enriched in transcriptional activators around paraspeckles. This could occur either by sequestering factors (dark and light green) that typically suppress the assembly of transcriptional activators (magenta) (C) or by providing seeds for the assembly of the factor in the vicinity of paraspeckles (D). Additionally, the decrease in temperature may assist in the formation of such compartments by influencing the phase behavior of transcriptional activators containing IDRs. The formation of the transcriptionally active compartment may subsequently recruit gene clusters to be upregulated during the differentiation of beige adipocytes, resulting in their movement toward paraspeckles.
It has previously been proposed that human NEAT1 is involved in the nuclear retention of mRNAs for nuclear-encoded mitochondrial proteins, thereby controlling the function of mitochondria (Wang et al. 2018). Consistently, we also observed dysregulation of mRNAs of mitochondrial proteins 24 h after cold exposure. However, the shape and number of mitochondria in mouse embryonic fibroblasts prepared from Neat1 KO embryos appear indistinguishable from those in fibroblasts from WT embryos (H Toya and S Nakagawa, unpubl.). This suggests the possibility that the control of mitochondrial mRNA localization might be a species-specific event. Consequently, it remains unclear whether the decreased mRNA expression of mitochondrial proteins in our model is directly caused by the disappearance of paraspeckles or is an indirect effect of impaired beige cell differentiation. Investigating the subcellular distribution of the affected mRNAs via in situ hybridization would be essential to test this hypothesis. However, this approach is technically challenging because of the presence of large lipid droplets in iWATs, which hinder the preparation of frozen sections suitable for in-depth analyses of subcellular mRNA distribution using in situ hybridization. The presence of lipid droplets also perturbs the preparation of subcellular fractions for biochemical analyses. Further technical improvements are therefore essential to overcome these issues and validate this possibility.
Beige adipocytes share various biochemical and physiological properties with brown adipocytes, and key transcription factors such as PPARγ, PRDM16, C/EBPβ, and PGC-1α regulate the differentiation of both thermogenic cell types (Inagaki et al. 2016). Despite the severe impairment in beige cell differentiation observed in Neat1 KO mice, the differentiation of brown adipocytes occurs normally. The robust upregulation of Neat1_2 evident in iWAT is absent in BAT, suggesting that Neat1 and paraspeckle formation may not be essential for brown adipocyte development. The specific requirement of Neat1_2 for beige adipocyte differentiation, as opposed to brown adipocyte differentiation, currently remains unclear. A possible explanation could be the difference in environmental temperatures that nonthermogenic iWAT and thermogenic BAT experience. Prior studies using telemetric transmitters have shown that subcutaneous temperature decreases transiently when mice are exposed to a cold environment (Ishii et al. 1996). Thus, iWAT cells might be subject to lower temperatures upon cold exposure, which could lead to changes in the phase-transition properties of IDR-containing proteins in the nucleus. The induction of Neat1 and the ensuing formation of paraspeckles might modify the phase diagram by sequestering IDR-containing proteins, thereby promoting the formation of functional transcriptional condensates necessary for beige cell differentiation. It would be intriguing to examine the dynamic behavior and kinetics of IDR-containing transcription factors in cells with or without paraspeckles at lower temperatures within a physiological range.
During the differentiation of beige adipocytes, a distinct isoform shift of Neat1 was noted: Neat1_1 is the primary isoform expressed under normal conditions, while differentiation triggers the induction of Neat1_2. This induction coincides with the disappearance of microspeckles, structures identified in cells exclusively expressing Neat1_1 (Li et al. 2017). A similar emergence of the longer isoform has been observed during the differentiation of human embryonic stem cells, where TDP43 has been implicated in promoting the polyadenylation of Neat1, favoring the production of the short isoform, Neat1_1 (Modic et al. 2019). The switch between isoforms has also been reported to be regulated by the opposing actions of CPSF6 and HNRNPK, which, respectively, promote and inhibit the polyadenylation necessary for Neat1_2 production (Naganuma et al. 2012). However, in the context of beige adipocyte differentiation, no significant differences in the localization or expression levels of Tardbp, Cpsf6, and HnrnpK were detected in mice (H Toya, unpubl. observation). To elucidate the mechanisms governing Neat1 isoform switching in vivo, further research is warranted, including analysis of modifications to these regulatory proteins.
Traditionally, the focus has been on domains with distinct structures predicted by primary sequences due to their clear functional roles. However, the primary sequences of lncRNA, including Neat1, are much less conserved compared to classical functional noncoding RNAs such as ribosomal RNAs or UsnRNAs (Mattick et al. 2023). Furthermore, the significance of IDRs and other nondomain structures, often less conserved than globular protein domains, is increasingly acknowledged (Uversky 2021). These “nondomain” biomolecules, whose functions cannot be readily predicted from their primary sequences, play critical roles in cellular processes and achieve this through their inherent flexibility and ability to participate in the formation of molecular condensates via multivalent interactions (Arakawa et al. 2023). Our study contributes to this emerging perspective by illustrating how Neat1, potentially interacting with IDRs and other nondomain structures, plays a pivotal role in beige adipocyte differentiation. This aligns with the understanding that nonconserved sequences can still be functionally significant, highlighting the importance of the reverse-genetic approach to decipher the function of “nondomain” biopolymers.
MATERIALS AND METHODS
Animals
All experiments were approved by the safety division of Hokkaido University (Approval 2021-027). All mice experiments were approved by the Animal Care and Use Committee of Hokkaido University (Approval 20-0031). Neat1 KO mice (Nakagawa et al. 2011) were bred and maintained under a C57BL/6N genetic background. These mice were housed at a stable temperature of 23 ± 1°C, under a consistent 12 h light–dark cycle. For experiments requiring cold exposure, mice were specifically housed at 4°C, maintaining the same light–dark cycle. For the purpose of anesthetization, a combination of medetomidine, midazolam, and butorphanol was intraperitoneally injected at a volume of 10 µL/g of body weight.
Histology analysis
The dissected tissues were methodically dehydrated in a series of ethanol solutions (70%, 95%, 100% ethanol in water) for 1 h each, followed by two 90 min treatments in absolute ethanol, and three 30 min treatments in chloroform at room temperature. Subsequently, the dehydrated tissues were embedded in paraffin. Paraffin sections of 8 µm thickness were collected onto noncoated slide glasses (MATSUNAMI S2111) and stained with hematoxylin (Sigma HHS16) and eosin Y solution (Sigma HT110116) following the manufacturer's recommended protocol.
Immunohistochemistry
Paraffin sections were first deparaffined in a series of xylene and ethanol solutions. After complete dehydration, the sections were treated with 0.1% proteinase K in phosphate-buffered saline (PBS) at room temperature for 5 min for epitope retrieval. This was followed by blocking endogenous peroxidase using 1% H2O2 and 5% serum in Tris-buffered saline (TBS). The samples were then incubated with the primary antibody solution in 5% serum in TBS for 1 h at room temperature. This was followed by incubation with a secondary antibody conjugated to HRP and detected by DAB staining using the ImmPACTDAB Substrate Kit (Peroxidase SK-4105). Antibodies used in this study are shown in Supplemental Table S3.
Western blotting
Tissues were homogenized in PBS and centrifuged at 800g for 20 min at 4°C. The interphase layer was collected to remove any lipids, and an equal volume of 2× SDS sampling buffer (containing 4% SDS, 129 mM Tris-HCl pH 6.8, 20% glycerol, and 5% 2-mercaptoethanol) was added. This mixture was then boiled for 3 min and sonicated. The proteins were separated on 10% acrylamide gels and transferred to PVDF membranes using the Trans-Blot Turbo Transfer System & Trans-Blot Turbo RTA Transfer PVDF Kit (Bio-Rad 1704150J1). The membranes were blocked with 4% milk in TBST for 10 min, then incubated with the primary antibody solution followed by the secondary antibody solution at room temperature for 1 h each. The signal was detected using ECL prime (Cytiva RPN2232).
Northern blotting
RNA samples were separated on 1% agarose, 1× MOPS, 3% formalin gels. Postseparation, the gels were washed in distilled water, 0.05 N NaOH in distilled water, again in distilled water, and then in 20× SSC for 20 min each. The RNA was then transferred to nylon membranes (Merck 11209299001) at room temperature overnight. These membranes were UV cross-linked (1200 × 100 µJ/cm2), prehybridized in DIG Easy Hyb containing 100 ng/mL salmon sperm DNA and yeast t-RNA 100 ng/mL at 68°C for 30 min, and hybridized with the probe in DIG Easy Hyb (Merck 11603558001) at 68°C overnight. After several washes, the membranes were incubated with DIG-AP (Roche 11093274910) antibody in blocking reagent (Roche 11096176001), and detected with CDP-star (Sigma 11685627001) in detection buffer (0.1M Tris-HCl pH9.5, 0.1M NaCl).
RNA extraction and RT-qPCR
RNA extraction was performed using TRIzol reagent (Invitrogen 15596026) as per the manufacturer's protocol. An additional step involved heating the samples at 55°C for 10 min before adding chloroform to enhance Neat1_2 extraction, as previously reported by Chujo et al. (2017). A total of 0.5 µg of RNA was used as a template for reverse transcription, performed using ReverTra Ace qPCR RT Master Mix with gDNA Remover (TOYOBO FSQ-301). The resulting cDNA was diluted five times with distilled water and 1 µL of cDNA was used as a template for RT-qPCR. The RT-qPCR was performed using THUNDERBIRD Next SYBR (TOYOBO QPX-201) with the following PCR conditions: 95°C for 1 min, followed by 40 cycles of 95°C for 15 sec and 60°C for 1 min, and then 95°C for 10 sec, 65°C for 5 sec, and 95°C for 5 sec. The relative expression was calculated using the ΔCT method, and TFIIb was used as an internal control. The primers used in RT-qPCR are listed in Supplemental Table S3.
Probe preparation
Primers for cloning PCR were designed using Primer-BLAST and shown in Supplemental Table S3. The cDNA template for cloning PCR was prepared through reverse transcription using Revertra Ace (TOYOBO TRT-101). The cloning PCR was performed using PrimeSTAR Max DNA Polymerase (Takara R045A), and the resulting PCR products were purified using the Wizard SV Gel and PCR Clean-Up System (Promega A9282). The purified PCR products were then cloned into the pGEM-T Easy Vector (Promega A1360). For preparing the template for in vitro transcription, PCR was performed using M13 primers and the cloning vectors. The resulting PCR products were purified by a column. Labeled probes were synthesized through in vitro transcription using either SP6 (Roche 10810274001) or T7 (Roche 10881775001) polymerase and DIG (Roche 11277073910) or FITC (Roche 11685619910) labeled nucleotides, and then purified using molecular precisionCentriPure MINI Spin Columns Desalt Z-50 (BIOTECH MS-0101-Z100.0-001).
In situ hybridization to detect RNA
Anesthetized mice were perfused with HCMF buffer to remove the blood and subsequently with 4% paraformaldehyde in HCMF. The collected tissues were fixed overnight at 4°C in 4% paraformaldehyde in HCMF. The next day, the tissues were washed with PBS, immersed in 30% sucrose in 0.1 M phosphate buffer overnight at 4°C, and then embedded in OCT compound (Sakura). Frozen sections were cut at 25 µm and collected on PLL-coated slide glasses (MATSUNAMI S7441). The sections were then fixed with 4% paraformaldehyde in HCMF overnight at 4°C and processed as described previously (Nakagawa et al. 2011). The probes used in this study are listed in Supplemental Table S3. Briefly, the samples were treated with 0.2 N HCl, proK (3 µg/mL) in 100 mM Tris pH 8.0, 10 mM EDTA for 7 min at 37°C, quenched with 0.2% glycine in PBS, and fixed with 4% paraformaldehyde in HCMF. They were then acetylated with an acetylation solution (triethanolamine 3 mL, concentrated HCl 0.5 mL, acetic anhydride 0.5 mL, up to 200 mL with DW). After 2 h of treatment with prehybridization solution at 55°C, DIG or FITC-labeled probes were hybridized overnight at 55°C in the hybridization solution. The samples were washed twice in wash buffer, treated with RNaseA buffer to decrease nonspecific signals, and then washed in 2× and 0.2× SSC. Probes were detected by antibodies against DIG or FITC and secondary antibodies conjugated with fluorescence.
Re-analyses of single-cell RNA-seq data set
We used sample annotation data (barcode) and gene annotation data (feature) from GSE133486 (Rajbhandari et al. 2019) registered in the NCBI GEO database, along with gene expression matrix data (expression.matrix). Subsequent analysis was conducted using the Seurat package (Hao et al. 2024) in R for single-cell RNA-seq analysis. Cells were filtered based on the number of expressed genes per cell (200, <2500), and then standardized using LogNormalize. UMAP was used for dimension reduction followed by clustering. Cell type labels were assigned based on labels provided in the metadata. Subsequently, Feature plots were used to plot the expression levels of Neat1 in each cell.
Flow cytometry
Collagenase solution containing 2 mg/mL collagenase (Wako 034-22363) and 2% BSA (Sigma A6003) in DMEM/high glucose was prewarmed at 37°C. Dissected iWAT was perfused with the collagenase solution and incubated for 1 h at 37°C. The digested tissues were then passed through a 200 µm mesh and centrifuged at 1000 rpm for 5 min. Pellets were dissolved in ACK Lysis buffer, rinsed in wash buffer (2% fetal bovine serum [FBS] in PBS), and the cells were incubated with antibodies in wash buffer. The antibodies used in FACS are listed in Supplemental Table S3. Stained cells were washed with wash buffer and centrifuged at 3000 rpm for 5 min twice, then passed through a 40 µm mesh. The cells were counted by a suitable method and analyzed by a compatible system. Cells were filtered using FSC-H and FSC-A plots to remove doublet cells, and were gated based on each fluorescence label. For precursor cell gating, PE-Cy7:CD31+ cells were identified as vascular endothelial cells. Among the remaining PE-Cy7:CD31– cells, PE:Cd34+ and PerCP-Cy5.5:ScaI+ cells were gated. Subsequently, APC:PDGFR+ cells were also gated and counted as precursor cells. For macrophage cell gating, APC:F4/80+ cells were identified as macrophage cells. Within these cells, FITC:Cd11c+ cells were gated as M1 macrophages, and PerCP-Cy5.5:Cd206+ cells were gated as M2 macrophages.
RNA-seq
Total RNA had rRNA removed using mouse/rat riboPOOL (standard RNA-seq DP-P024-000055, siTOOLs Biotech), and libraries were prepared using the TruSeq Stranded Total RNA Library Prep Gold (Illumina). Samples were sequenced on the HiSeqX_Ten platform at Macrogen. Obtained reads were quality-checked using FastQC and mapped to the mouse genome (Mus_musculus.GRCm38.84.gtf) using HISAT2 (Kim et al. 2019). Mapped reads were counted using featureCounts (Liao et al. 2014), and differential expression analysis was performed using DESeq2 (Love et al. 2014). Further genome enrichment analysis was performed using iDEP (Ge et al. 2018). Sequencing data obtained in this study are available at PRJDB17455.
iDEP and RNAseqChef analyses
For the RNAseqChef analysis, the raw count file and metadata file were uploaded to the RNAseqChef website (https://imeg-ku.shinyapps.io/RNAseqChef/) and processed using the Multi DEG pipeline. For UMAP analysis, the number of neighbors was set to 5. Default parameters were used for other analyses. To identify clusters enriched in differentially expressed genes, raw count data were uploaded to the iDEP website (http://bioinformatics.sdstate.edu/idep96/) and analyzed using the Genome pipeline with the following parameters: window size = 6 Mb, steps = 2 Mb, FDR cutoff for window = 1 × 10−4 for the comparison between 24 h WT and 24 h KO samples, and 0.01 for the comparison between 0 h WT and 24 h WT samples.
Simultaneous detection of DNA loci and RNA
For DNA probe preparation, BAC DNAs for the target regions were obtained from the RIKEN BioResource Research Center (BRC). BAC clones used in this study are shown in Supplemental Table S3. BAC DNA probes were synthesized using the Nick Translation Kit (Roche 10976776001) and labeled with fluorescent dUTPs. Following the manufacturer's instructions, BAC DNA, Nick Translation Enzymes, and dNTPs were mixed. The Nick Translation reaction was performed at 15°C for 60 min, stopped by adding stop buffer, and heated at 65°C for 5 min. DNA probes were purified using the QIAquick PCR Purification Kit (QIAGEN 28104), then sonicated. RNA FISH was performed as previously described. To enhance the RNA FISH signal diminished by DNA FISH treatment, tyramide signal amplification (Kiko Tech NEL741001KT) was used. After probe washing, tissues were blocked with TSA Blocking Reagent (Kiko Tech FP1020) at room temperature for 30 min. Probes were detected using HRP-conjugated DIG or FITC antibodies. Following several washes, TSA amplification was conducted on the slide at room temperature for 10 min. To stop the reaction, samples were washed multiple times and fixed with 4% PFA for 15 min. DNA FISH was then performed. DNA probes and Cot1 DNA (50 ng/sample each) were prepared by ethanol precipitation, air-dried briefly, and dissolved in 100% formamide at 50°C for 60 min. 2× Hybridization buffer was added to the probe solution, which was then denatured at 95°C for 5 min and kept at 37°C until use. Tissues on slides were dehydrated in 70% and 100% ethanol, denatured in 70% formamide/2× SSC at 75°C for 5 min, and quickly chilled on ice. After dehydration in ice-cold 70% ethanol and 100% ethanol, slides were dried. Denatured probes were applied to the slides, and hybridization was carried out at 37°C overnight. Samples were washed three times in 2× SSC with 0.01% Triton, once in 2× SSC with 0.01% Triton, and twice in 0.2× SSC with 0.01% Triton at 45°C. Finally, samples were mounted with PVA containing DAPI (1 µg/mL).
Measurement of paraspeckles and DNA loci
The Z-stack images of paraspeckles and DNA foci were loaded into the ImageJ plugin DiAna (Gilles et al. 2017). The images were filtered with a Gaussian blur, and the threshold was set appropriately before segmentation. Other parameters (radius, minimum, and maximum object size) were applied using the default settings. The center-to-center 3D distance between DNA foci and Neat1 foci was measured in the segmented images, and the shortest distance between paraspeckles and DNA foci was used for further analysis.
Statistical analysis
Expression levels of genes essential for the differentiation and/or function of beige adipocytes were compared between cold-exposed and control groups using a nonparametric bootstrap test with 1000 surrogates implemented in the R. The data on Neat1 foci number and area were analyzed using the Mann–Whitney U-test, implemented in GraphPad Prism9 (GraphPad Software). In a time course experiment to investigate changes in gene expression after cold exposure, we compared the relative expression levels to Gapdh without cold exposure with that after 4 h, 24 h, and 4 days after cold exposure using a nonparametric bootstrap test with 1000 surrogates followed by Bonferroni correction implemented in the R. Proportions of adipocyte precursors, endothelial cells, and M1/M2 macrophages in iWAT of WT were compared with those of Neat1 KO mice using Mann–Whitney U-test, implemented in GraphPad Prism9 (GraphPad Software). The data on distances between paraspeckles and genomic loci were analyzed using one-way analysis of variance (ANOVA) followed by Dunnett's post hoc test, implemented in GraphPad Prism9 (GraphPad Software).
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
We thank Dr. Yoshifumi Yamaguchi for helpful discussions on cold responses, Drs. Chitoku Toda and Hiroshi Kitamura for discussions at the initial stage of this study, Drs. Ling-ling Chen and Meng-Ran Wang for discussions on the RNA-seq data set, and Ms. Ikuko Nomura for careful reading of the manuscript. This work was supported by JSPS KAKENHI grant numbers 21H05274 and 21K19246 granted to S.N., Grant-in-Aid for JSPS Fellows granted to T.Y., as well as RIKEN Pioneering Projects “Biology of Intracellular Environments” to S.I. This work was partly supported by the Global Facility Center (GFC), Hokkaido University.
Footnotes
-
Handling editor: Ling-Ling Chen
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079972.124.
- Received January 30, 2024.
- Accepted April 8, 2024.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








