
Spliceosome assembly and regulation: insights from analysis of highly reduced spliceosomes
- Corbin S. Black1,2,
- Thomas A. Whelan3,
- Erin L. Garside4,
- Andrew M. MacMillan4,
- Naomi M. Fast3 and
- Stephen D. Rader1
- 1Department of Chemistry and Biochemistry, University of Northern British Columbia, Prince George, British Columbia, Canada V2N 4Z9
- 2Department of Anatomy and Cell Biology, McGill University, Montréal, Quebec, Canada H3A 0C7
- 3Biodiversity Research Center and Department of Botany, University of British Columbia, Vancouver, British Columbia, Canada V6T 1Z4
- 4Department of Biochemistry, University of Alberta, Edmonton, Alberta, Canada T6G 2H7
- Corresponding authors: rader{at}unbc.ca, amacmill{at}ualberta.ca, nfast{at}mail.ubc.ca
Abstract
Premessenger RNA splicing is catalyzed by the spliceosome, a multimegadalton RNA–protein complex that assembles in a highly regulated process on each intronic substrate. Most studies of splicing and spliceosomes have been carried out in human or S. cerevisiae model systems. There exists, however, a large diversity of spliceosomes, particularly in organisms with reduced genomes, that suggests a means of analyzing the essential elements of spliceosome assembly and regulation. In this review, we characterize changes in spliceosome composition across phyla, describing those that are most frequently observed and highlighting an analysis of the reduced spliceosome of the red alga Cyanidioschyzon merolae. We used homology modeling to predict what effect splicing protein loss would have on the spliceosome, based on currently available cryo-EM structures. We observe strongly correlated loss of proteins that function in the same process, for example, in interacting with the U1 snRNP (which is absent in C. merolae), regulation of Brr2, or coupling transcription and splicing. Based on our observations, we predict splicing in C. merolae to be inefficient, inaccurate, and post-transcriptional, consistent with the apparent trend toward its elimination in this lineage. This work highlights the striking flexibility of the splicing pathway and the spliceosome when viewed in the context of eukaryotic diversity.
Keywords
- Cyanidioschyzon merolae
- homology modeling
- reduced spliceosomes
- spliceosome diversity
- spliceosome structure
INTRODUCTION
Pre-mRNA splicing
Pre-mRNA splicing is an essential step of gene expression in eukaryotes. It consists of the removal of introns and ligation of protein-coding exons via two transesterification reactions. Splicing proceeds via a complex process involving assembly of 80–200 proteins and RNA splicing factors that recognize conserved sequences in each pre-mRNA, principally the 5′ splice site (5′SS), 3′ splice site (3′SS), and branchpoint sequence (BPS). The complex formed by these factors is known as the spliceosome, and assembles anew for each round of splicing (Cheng and Abelson 1987; Hoskins et al. 2011; Cordin and Beggs 2013).
Spliceosome assembly on pre-mRNA substrates in both HeLa and yeast cell extracts has been viewed as proceeding in a stepwise fashion through discrete complexes. Historically, the progression from the prespliceosomal complex A through the spliceosomal B and C complexes was characterized by native gel electrophoresis as well as sedimentation analysis (Konarska and Sharp 1986, 1987; Cheng and Abelson 1987). This assembly pathway included the early or commitment complex (E/CC) as a precursor to the A complex (Seraphin and Rosbash 1989; Das and Reed 1999). Subsequent biochemical, genetic, and mass spectrometric analyses have elaborated this description (for review, see Will and Lührmann 2011), which has now been detailed in multiple high-resolution structural analyses by Cryo-EM as described below.
In Saccharomyces cerevisiae (yeast), the 5′SS is initially recognized by the U1 small nuclear RNA (snRNA) and its associated proteins, which are assembled into a small nuclear ribonucleoprotein particle (snRNP; Cheng and Abelson 1987). Meanwhile, proteins BBP/Msl5 and Mud2 interact with the BPS and polypyrimidine tract (Berglund et al. 1997; Chen and Cheng 2012). At this stage, the assembling particle is referred to as the E complex. BBP/Msl5 is subsequently replaced at the BPS by a stable interaction with the U2 snRNP to form the A complex or prespliceosome (Yan and Ares 1996). The remaining snRNPs, U4, U5, and U6, preassociate in the tri-snRNP and bind to the A complex to yield the precatalytic B complex (Cheng and Abelson 1987). The A complex undergoes a series of rearrangements catalyzed by ATPases Prp28 and Brr2 (Raghunathan and Guthrie 1998a,b) resulting in replacement of U1 snRNA by U6 snRNA at the 5′SS and destabilization and release of the U4 snRNP to yield the activated B complex (Bact; Strauss and Guthrie 1991; Staley and Guthrie 1998). The ATPase Prp2 converts Bact to B*, which carries out the first transesterification reaction to generate the C complex (Liu and Cheng 2012; Ohrt et al. 2012). The ATPase Prp16 displaces the first-step factors Cwc25 and Yju2 (Schwer 2008; Chiang and Cheng 2013; Tseng et al. 2017), allowing the second-step factors Slu7 (Frank and Guthrie 1992) and Prp18 (Brys and Schwer 1996; James et al. 2002) to bind and form the C* complex, which carries out the second catalytic step resulting in the post-catalytic P complex (Chen et al. 2013). Finally, the spliceosome is disassembled through the action of Prp22, a DEAH-box helicase that releases the mRNA (Schwer and Gross 1998; Schwer 2008), and the Ninteteen-Related (NTR) Complex, which disassembles the spliceosome (Chen et al. 2013). The assembly and disassembly steps in metazoan splicing are substantially similar (Will and Lührmann 2011).
For several decades following the discovery of splicing in 1977, central goals of the splicing field were to define the composition of the spliceosome, obtain a general sense of its three-dimensional organization (Chow et al. 1977), and elucidate its mechanism of action (Berget et al. 1977). With the publication of high-resolution cryo-EM structures of the spliceosome in 2015 (Nguyen et al. 2015; Yan et al. 2015), the field has seen a dramatic shift yielding essentially complete views of nearly every step in the splicing cycle including the location of many splicing factors (Agafonov et al. 2016; Abelson 2017; Bai et al. 2017, 2021; Bertram et al. 2017a,b; Zhan et al. 2018a,b; Fica et al. 2019; Zhang et al. 2019; Charenton et al. 2020; Townsend et al. 2020; Wilkinson et al. 2021; Tholen et al. 2022). The focus of splicing research has consequently shifted more toward the mechanism and evolution of splicing.
Our understanding of pre-mRNA splicing comes largely from studies on two organisms: S. cerevisiae and humans. Splicing evolved early in the evolutionary history of eukaryotes (Jurica and Moore 2003; Koonin 2006; Roy and Gilbert 2006) and as a result the general splicing machinery consisting of five snRNAs and their associated proteins appears similar across eukaryotes. However, examination of lesser-studied eukaryotes reveals unexpected plasticity in the splicing process and spliceosome composition. An excellent example of this plasticity is the phenomenon of trans splicing in which a Splice Leader sequence is attached onto all mRNA transcripts via the spliceosome (Günzl 2010). Trans splicing was discovered in trypanosomes (Boothroyd and Cross 1982), and has since been observed in a number of kinetoplastids, nematodes, and other lineages including chordates (Günzl 2010). Splicing in T. brucei mainly occurs through the trans pathway although two canonical, cis-spliced introns have been retained (Mair et al. 2000), demonstrating that the T. brucei splicing machinery is completely capable of cis splicing despite the loss of all other cis-spliced introns.
One mystery of trypanosome splicing is the relatively high expression level of the U1 snRNP. Although in nematodes the U1 snRNP is only required for cis splicing, in trypanosomes it is highly abundant and apparently associated with the spliceosome, even though there are only two introns on which it would function (Günzl 2010). U1's abundance suggests that trypanosomal U1 may have other roles outside of splicing (West 2012), such as human U1 snRNP's role in transcription regulation (Spiluttini et al. 2010; O'Reilly et al. 2013; Vazquez-Arango et al. 2016), or the role of the U2 snRNP in histone mRNA maturation (Friend et al. 2007; So et al. 2019; Zhang et al. 2021).
A variety of other insights into splicing evolution have come from studying organisms beyond humans and S. cerevisiae. Phylogenetic analysis of splicing proteins revealed that early spliceosomes were similarly complex to current plant and animal spliceosomes (Roy and Irimia 2014), and that many spliceosomal proteins are derived from ribosomal proteins and evolved after intron proliferation (Vosseberg et al. 2023). Phylogenetic analysis of introns showed that there had already been substantial intron loss by the time of the Last Eukaryotic Common Ancestor (LECA) (Vosseberg et al. 2022). This points to early proliferation of introns, and hints at nucleogenesis substantially predating LECA (Vosseberg et al. 2022). Comparison of U1-70k proteins in plants showed a proliferation of protein architectures that may be important for diversity of stress responses, and also revealed possible nonsplicing roles of this protein family, for example, in osmotic stress response (Chen et al. 2020). Analysis in five Entamoeba species demonstrated coevolution of introns and spliceosomes, and revealed intron proliferation in an intron-poor lineage (Roy and Bowser 2022). Deletion of splicing factors in the intron-rich yeast Cryptococcus suggested that many splicing proteins conserved in humans have a role in the fidelity of splice site detection (Sales-Lee et al. 2021). Finally, studies in diplomonads and parabasalids provide examples of divergent snRNAs (Hudson et al. 2019). These examples highlight the diversity of splicing systems outside the most commonly studied model organisms.
One potential driver of spliceosomal diversity is genome reduction. Reductions in genome size are accompanied by the loss of noncoding regions, including spliceosomal introns. As has been observed in multiple, distantly related lineages including trypanosomes (Günzl 2010) and giardia (Hudson et al. 2012), loss of introns is mirrored by a reduction in the RNA and protein components of the spliceosome (Jeffares et al. 2006; Grisdale et al. 2013; Hudson et al. 2015). These events—genome reduction and spliceosome simplification—provide us with both mechanistic and evolutionary insights into the spliceosome. In this review, we first examine diverse eukaryotic lineages with a focus on taxa with reduced genomes to identify common patterns of spliceosomal component loss, before turning to more closely examine the red alga Cyanidioschyzon merolae (C. merolae or Cm), which has extreme levels of spliceosome reduction.
Comparing spliceosomes across diverse eukaryotes
Using methods similar to Wong et al. (2021), we searched the proteomes of 40 distantly related eukaryotic taxa (Supplemental Table 1) for 71 spliceosomal proteins found in yeast and humans to assess the completeness of the spliceosome across eukaryotes, and to determine if there are any patterns to spliceosomal reduction. We increased sampling of taxa possessing fewer introns, because we predict they have reduced spliceosomes. In line with the current understanding of genome reduction in eukaryotes, we observe multiple instances of independent spliceosomal reduction, the most extreme examples of which can be seen in the microsporidia (24/71 proteins found for E. cuniculi), Giardia (17/71), the kinetoplastids (38/71 on average), and C. merolae (26/71; Fig. 1). Other lineages with notably reduced spliceosomes include P. cylindrospora (fungi; 41/71) and E. histolytica (amoebozoa; 49/71). Spliceosome reduction has therefore occurred repeatedly across the tree of life, although we have seen no evidence of it in certain groups such as plants and green algae, animals, and stramenopiles.
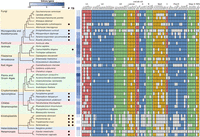
Comparison of spliceosomes across diverse eukaryotes. Eukaryotic relationships follow (Burki et al. 2020) with common taxonomic groupings indicated (left). Parasitic taxa (P) and those that possess trans-splicing (TS) are indicated by dots in columns. Annotated introns/gene shown for each taxon with darker boxes indicating higher intron density. Phytomonas and Perkinsela have no annotated introns. Spliceosome protein presence is indicated with a filled box. Faint colors indicate proteins curated as present. Predicted absence of a protein is indicated by an unfilled box. Cm Nam8 was bioinformatically annotated but could not be identified in mass spectrometry experiments (Reimer et al. 2017).
Considering the life histories of the groups with highly reduced spliceosomes, it is clear that genome reduction correlated with parasitism plays a factor in spliceosomal composition. Microsporidia, kinetoplastids and Giardia are all parasites that have very few introns and have lost substantial numbers of spliceosomal proteins (Fig. 1). However, this pattern is not observed in all parasites. For example, the intracellular apicomplexan parasite that causes malaria, Plasmodium falciparum, does not show the same levels of spliceosome reduction, despite a significantly reduced genome. While Phytomonas sp. and Perkinsela sp. have no annotated spliceosomal introns, the kinetoplastids retain an extremely reduced spliceosome involved in trans-splicing mRNA (Fig. 1).
Some splicing factors are more frequently lost than others, suggesting they are less essential for splicing (Fig. 1). For example, components of the U1 snRNP, the Bact complex, and the Prp19/Nineteen Complex (NTC) are frequently lost. Mud1 is missing in 27 of 40 lineages, and Prp39 is missing from 22. Similarly, the U5 snRNP protein Snu40 is missing in 26 lineages, Snu71 of the A complex is missing in 20, and Spp2 and Cwc25 of the Bact complex are missing in 22 and 21, respectively. In contrast, even in the reduced taxa, the U2 snRNP remains relatively complete, as does the U5 snRNP and the set of second step proteins (Fig. 1).
The four proteins present in all 40 lineages are by inference the most indispensable. These are Hsh155 and Prp9 from the U2 snRNP, and Dib1 and Prp8 from the U5 snRNP. Other highly retained proteins include Cus1, Hsh49, and Rds3 from the U2 snRNP; Brr2 and Snu114 of the U5 snRNP; Snu13 of the U4/U6 snRNP; Msl5 of the A complex; Prp38 and Sub2 of the B complex; Cwc22 of Bact; Cef1, Clf1, Prp46, and Syf1 from the Nineteen Complex; Prp17 of the second step complex; and Ist3 of the RES complex. All of these are present in at least 90% (36) of the lineages surveyed. The absence in some lineages of proteins thought to be essential for splicing, such as Brr2 and Snu114, raises questions about the quality of the sequencing and annotation in some of these cases, but the overall trends are clear.
While we have not surveyed snRNAs here, due to the challenges of unambiguously identifying them in divergent lineages, we are not aware of any organisms that lack the U2, U4, U5, or U6 snRNAs. In contrast, U1 snRNP has been shown to be dispensable in a number of situations (e.g., for trans splicing; Günzl 2010; Stark et al. 2015) and is known to be absent in Cm (Stark et al. 2015). When considering those taxa with reduced spliceosomes, Cm stands out as being the only taxon that is extensively reduced without having a parasitic life history and while still retaining canonical splicing.
An experimentally validated reduced spliceosome
C. merolae is a rhodophyte, a deep-branching unicellular red alga that is thought to have diverged from fungi and humans approximately 1.5 billion years ago (Yoon et al. 2004). Phylogenetic evidence suggests that the ancestral state of the Cm spliceosome and intron complement was much closer to that of fungi such as S. pombe, that is, the genome contained several thousand introns and a larger spliceosome that included the U1 snRNP (Hudson et al. 2015). We have studied pre-mRNA splicing in Cm (Stark et al. 2015; Black et al. 2016; Reimer et al. 2017; Garside et al. 2019), and previously reported that it possesses a unique algal spliceosome lacking the entire U1 snRNP and several other splicing and spliceosome assembly proteins (Table 1; Fig. 2; Stark et al. 2015; Reimer et al. 2017). The Cm spliceosome therefore reflects divergence from other canonical spliceosomes, primarily in the loss of many splicing factors. It is surely not a coincidence that in the Cm lineage all but 38 introns have been eliminated (Wong et al. 2022).
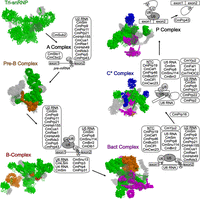
Overview of the Cm splicing proteins present throughout one pre-mRNA splicing cycle. Proteins are colored according to incorporation into the spliceosome, whereas proteins absent in Cm are shown in transparent gray. The color scheme is as follows: Tri-snRNP (green, PDB 3JCR); Pre-B complex (orange, PDB 6AH0); B-complex (red, PDB 5O9Z); Bact complex (purple, PDB 5GM6); C* complex (blue, PDB 5MQF); P complex (no additional proteins, PDB 5YLZ).
Proteins present in cryo-EM spliceosomal structures missing in C. merolae
Despite their limited numbers, Cm introns appear unremarkable. Intron lengths range from 70 to 1309 nts, branchpoint to 3′ splice site distances range from 10 to 63 nt, and the 5′ and 3′ splice sites have canonical consensus sequences (GUAAGU and AG, respectively; Matsuzaki et al. 2004; Ogasawara et al. 2004). The branchpoint sequence is slightly divergent, ACUAACC, with a compensatory change in the U6 snRNA in which the canonical ACAGAGA sequence is changed to ACUGAGA (Stark et al. 2015). In contrast to S. cerevisiae, there is no obvious commonality among the intronic host genes. While two are ribosomal protein genes, and two others are also related to translation, the remainder have a wide variety of functions including protein folding, primary metabolism, and transport. No intron property yet tested correlates with the level of splicing across intron-containing genes, which is strikingly low (Schärfen et al. 2022).
Based on the presently known inventory of splicing-related factors, we propose the assembly of the Cm spliceosome to be novel. For instance, while the branchpoint sequence (BPS) is predicted to be recognized by the branchpoint bridging protein CmMsl5 (BBP, SF1) and CmMud2 (U2 auxiliary factor U2AF65), 5′ splice site (5′SS) recognition remains a mystery due to the absence of the U1 snRNP. Notably, CmMsl5 is missing the SPSP and PPxY motifs, which have been shown to interact with Prp40 and other U1 snRNP proteins absent in Cm (Chang et al. 2012). The SPSP and PPxY motifs are within a coiled-coil domain and regulate the splicing of a subset of human pre-mRNA transcripts (Lipp et al. 2015). Like yeast, Cm is also missing U2AF35, which forms a dimer with U2AF65 in S. pombe (Huang et al. 2002) and humans (Zuo and Maniatis 1996). The absence of U2AF35 is interesting because it might have a cotranscriptional function and hints at splicing and transcription being uncoupled in Cm (Wu and Fu 2015). The absence of Lea1 and Msl1, discussed later, is further possible evidence of uncoupled splicing (Gunderson and Johnson 2009). The absence of the U1 snRNP would also preclude the existence of early spliceosomal states such as the E and A complexes, at least as they are known in other organisms. If spliceosome assembly in Cm proceeds analogously to assembly in yeast, recruitment of the U2 snRNP to the pre-mRNA followed by incorporation of the tri-snRNP would presumably result in the precatalytic B complex.
To complement our experimental data, and to compare the Cm spliceosome with that from yeast and humans, we reviewed structures of spliceosomal complexes from throughout the splicing cycle. We gathered all available sequences from known Cm splicing proteins, all of which have been validated by affinity purification followed by mass spectrometry (AP-MS; Reimer et al. 2017; MR Stark, SD Rader, M Oeffinger, et al., unpubl.), and performed pairwise sequence alignments against human and yeast orthologs. Interestingly, many splicing proteins that are highly conserved between yeast (Hossain and Johnson 2014), humans, and even Galdieria sulphuraria (Qiu et al. 2018), a red alga found to cohabit extreme environments with Cm, show substantial divergence in Cm (Matsuzaki et al. 2004; Stark et al. 2015). Table 2 lists those Cm proteins that lack domains conserved in their orthologs, while Table 3 lists Cm proteins with domains that are not found in their yeast or human homologs. Supplemental Figure 1 shows multiple sequence alignments of key proteins mentioned below. Using the pairwise scores generated from the Needleman–Wunsch algorithm, we annotated the respective proteins in structures from human and yeast spliceosomes (see Table 4 for PDB accession numbers of structures used; Madeira et al. 2019) to highlight presence, absence, and degree of conservation. These data about absent proteins allowed us to model each step of the splicing pathway using cryo-EM structures, and compare Cm to both human and yeast models. Based on the resulting models, we assessed the literature to predict what functions might be conserved, lost, or unique to Cm splicing proteins. These comparisons have allowed us to propose hypotheses about spliceosome structures and mechanisms under the assumption that there are no Cm-specific proteins or snRNAs that have taken on the roles of absent splicing factors. Extensive AP-MS analysis has not yet identified novel factors (Reimer et al. 2017; MR Stark, SD Rader, M Oeffinger, et al., unpubl.).
Conserved domains in complex spliceosomes absent in C. merolaea
Predicted novel domains in C. merolae splicing factorsa
Representative spliceosomal structures
ANALYSIS OF HOMOLOGY MODELS
5′ splice site recognition: a novel role for U5 snRNA?
It is possible that spliceosome assembly in Cm begins with the B complex (Zhan et al. 2018a,b; Fig. 3). There are currently no data to support the presence in Cm of any component of the U1 snRNP or proteins whose functions are related to U1 snRNP such as the DExD/H-box ATPase Prp28 (Stark et al. 2015). In other organisms, Prp28 displaces the U1 snRNP from the 5′SS (Mathew et al. 2008), which allows U6 snRNA in the tri-snRNP to base pair at the 5′SS (Lesser and Guthrie 1993; MacRae et al. 2018). It is unknown how the Cm tri-snRNP is recruited to the spliceosome prior to B complex formation. We previously proposed that 5′SS recognition occurs during tri-snRNP integration, whereby either U6 snRNA or U5 snRNA interacts with the 5′SS (Stark et al. 2015). Evidence for the Cm U6 snRNA:5′SS direct interaction is found in U6 snRNA's extended complementarity to 5′ splice sites: six of seven positions are able to form standard Watson–Crick base pairs (Fig. 4A), which is comparable to the canonical U1 snRNA:5′SS interaction (Crispino and Sharp 1995; Maroney et al. 2000) and longer than the canonical three-base-pair U6 snRNA interaction (Sawa and Abelson 1992; Wassarman and Steitz 1992). Interestingly, the 5′ end of Cm U5 snRNA (5′-GUCUGC) is complementary to all annotated Cm 5′ splice sites (Fig. 4B; Stark et al. 2015). Furthermore, U5 snRNA has previously been found to interact with the 5′SS during B complex formation in humans (Sontheimer and Steitz 1993; Boesler et al. 2015). We previously noted that the 450-nt Cm U5 snRNA is predicted to contain two more stem–loops than yeast or human U5 snRNA (Fig. 4B; Stark et al. 2015). If U5 snRNA is indeed responsible for initial 5′SS recognition, the additional domains of U5 snRNA may play a role in this step.
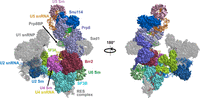
The human pre-B complex (PDB 6AH0). The U1 snRNP and all other proteins absent in Cm are shown in transparent gray.
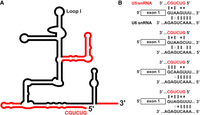
(A) Proposed secondary structure of the Cm U5 snRNA. Cm U5 (dark red) is overlaid on yeast U5 snRNA (black). Model was generated from the MFold RNA secondary structure prediction web server (Zuker 2003). (B) Proposed model for interaction between snRNAs and all sequence variants of 5′ splice sites in Cm. The 5′ splice site of the pre-mRNA is indicated (middle); complementary region of U5 snRNA is shown above, and that of U6 snRNA is shown below. Watson–Crick base pairs, G–U pairs, and G–A base pairs are indicated with symbols accordingly. Interactions with U5 and U6 snRNAs are proposed to occur sequentially.
There is ample precedent for redundancy in intron-recognition pathways from studies of splicing in HeLa cell extracts. High concentrations of SR proteins, and specifically SC35, can compensate for inactivation of the U1 snRNP after its depletion (Crispino et al. 1994) or blocking with an antisense oligonucleotide (Tarn and Steitz 1994), albeit only for some pre-mRNA transcripts. The transcript dependence may indicate that unknown properties of the pre-mRNA facilitate U1-independent splicing. Notably, increasing base-pairing between U6 and the 5′SS, as we see in Cm, increases the amount of U1-independent splicing in extract supplemented with SC35 (Crispino and Sharp 1995). The prior demonstration of U6-mediated suppression of U1 loss is consistent with the possibility that, in the absence of U1, intron recognition proceeds directly via U6 snRNA binding in Cm. There is a candidate SC35 homolog in Cm, but its expression level is not particularly high. Interestingly, U2AF65 can also be rendered unnecessary in HeLa extract by addition of SC35, but only in the presence of the U1 snRNP (MacMillan et al. 1997). These results point to inherent flexibility in intron recognition mediated by independent pathways, and suggest that U2, U5, or U6 snRNAs are all candidates for early roles in intron recognition.
CmBrr2 and predicted stability/regulation of the Cm tri-snRNP
As mentioned, the U4, U5, and U6 snRNPs join the assembling spliceosome as the preassembled tri-snRNP. The presence or absence and degree of similarity of Cm proteins compared to their human orthologs is revealed in the cryo-EM structure of the human tri-snRNP (Agafonov et al. 2016). Proteins that are absent in Cm are observed to be mainly peripheral in this structure, while the center of the tri-snRNP, mostly comprising Prp8 and three snRNAs, is conserved (Fig. 5). Cm lacks U5-40K, Prp6, Sad1, and the U1-associated DEAD-box helicase Prp28, which was previously discussed (Fig. 5). The absence of Prp6 is surprising, because it is required for tri-snRNP accumulation (Galisson and Legrain 1993) and bridges the U4/U6 di-snRNP and U5 snRNP in yeast and human spliceosomes (Fig. 5). U5-40K forms a trimer with Prp8 and Snu114 during assembly of the human U5 snRNP (Achsel et al. 1998), and later interacts with Prp17 in the precatalytic spliceosome (Sapra et al. 2008). It is conserved between animals and S. pombe (Cwf17), but is not present in yeast (Fabrizio et al. 2009) or Cm spliceosomes (Stark et al. 2015). The absence of Sad1 is correlated with the loss of its interacting proteins as well as other Brr2 helicase regulatory elements (Fig. 5, inset and Supplemental Figs. 1A, 2B).
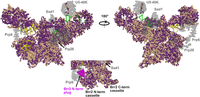
Surface representation of the Cm U4/U6.U5 tri-snRNP homology modeled against the human structure (PDB 3JCR). All protein sequences were annotated by conservation with their Cm homologs. Identical and similar amino acids are colored purple while dissimilar amino acids are colored wheat. Proteins absent in Cm are shown in transparent gray. The RNA is colored as follows: U6 (dark red); U4 (yellow); and U5 (green). (Inset) Surface representation of the amino- and carboxy-terminal cassettes of Cm Brr2 homology modeled against the human structure (PDB 3JCR). The amino-terminal plug domain of Brr2 that is absent in Cm is shown in magenta.
The Cm tri-snRNP could be destabilized by lack of Brr2 regulation
The Ski2-like RNA helicase Brr2 catalyzes unwinding of the U4/U6 snRNA duplex, causing the release of U4 snRNP and progression from the B complex to Bact (Mozaffari-Jovin et al. 2014). Brr2 activity is regulated by a network of proteins (Nguyen et al. 2013) including Prp8, Sad1, Snu114, Snu13, and its own amino-terminal region (Mozaffari-Jovin et al. 2013; Absmeier et al. 2015). Precise control of Brr2 activity is necessary for properly regulated splicing; unregulated helicase activity can result in disassembly of the tri-snRNP or ectopic activation of the spliceosome (Absmeier et al. 2015).
The Cm spliceosome lacks Sad1, which normally prevents premature dissociation of the tri-snRNP by regulating Brr2 (Huang et al. 2014). Specifically, Sad1 tethers Brr2 in an inactive conformation, preventing the central domain of U4 snRNA from loading into Brr2 (Agafonov et al. 2016). This inhibitory activity is necessary as Sad1 depletion results in premature tri-snRNP disassembly by Brr2 (Huang et al. 2014). Sad1 interacts with Prp31, Prp8, Snu114, and the PWI domain of Brr2 in the human tri-snRNP (Nguyen et al. 2016). Cm Brr2 lacks amino-terminal residues for the PWI domain, and Cm Snu114 has poor homology with its human ortholog at its interface with Sad1 (Supplemental Figs. 1A, 2B).
The amino-terminal region of Cm Brr2 is 300 residues shorter than many of its homologs (Supplemental Figs. 1A, 2B). It lacks the “plug” domain (Fig. 5, inset), which is necessary for preventing premature tri-snRNP dissociation as well as for efficient splicing (Absmeier et al. 2015). The amino-terminal cassette of Brr2 features a central RNA binding tunnel formed by the RecA-like, WH (winged-helix), and HB (helical bundle) domains (Absmeier et al. 2015) that binds a single-stranded region of U4 snRNA. The plug domain of the amino-terminal region interacts with the central tunnel's RecA-2 and HB domains to obstruct binding of the U4/U6 snRNA duplex and thereby prevents unwinding (Wollenhaupt et al. 2018).
In addition to the Brr2 auto-inhibition that prevents premature U4/U6 snRNA unwinding, the carboxy-terminal tail of the Prp8 Jab1 domain interacts with the Brr2 RecA-2 and HB domains and prevents their opening to accommodate U4 snRNA in the central domain; it also competes for tunnel occupation with single-stranded RNA regions (Absmeier et al. 2017). Together these two mechanisms help to ensure Brr2 activation is strictly controlled. With its absent plug domain, CmBrr2 appears to have lost a regulatory element of Brr2 activity (Fig. 5, inset) and the CmJab1 carboxy-terminal tail is also absent (Supplemental Fig. 2A). The Prp8 RNase H-like (RH) domain, however, also inhibits Brr2 activity by competing for U4 and U6 snRNA binding with Brr2 and thereby blocking Brr2 from loading its substrate (Mozaffari-Jovin et al. 2013). This domain is conserved in Cm and presumably inhibits Brr2 activity prior to activation. In the absence of the Brr2 auto-inhibitory plug, the Prp8 Jab1 tail, and Sad1, we suggest that U4 snRNA is preloaded in the Brr2 RNA binding tunnel as seen in the yeast tri-snRNP.
Cm lacks the U6 snRNP-associated protein Prp24, which promotes reannealing of the U4 and U6 snRNAs during tri-snRNP recycling in yeast (Raghunathan and Guthrie 1998a; Didychuk et al. 2016) and humans (Medenbach et al. 2004). It has been suggested that the U4 and U6 snRNAs can anneal via a Prp24-independent pathway if they are not completely dissociated by Brr2 (Huang et al. 2014). It is possible that Cm U4 and U6 snRNPs have evolved such a Prp24-independent annealing pathway related to the extra domain in U4 snRNA or the higher temperature at which Cm grows. For example, U4's extra domain could mediate interactions with the U6 snRNP that stabilize U4–U6 interaction (but not necessarily base-pairing) that prevents complete dissociation.
C. merolae B complex
The precatalytic spliceosome is formed upon displacement of the U1 snRNP from the 5′SS by U6 snRNA resulting in the departure of U1 and U4 snRNPs and the formation of the precatalytic B complex. Cm homology modeling reveals many peripheral proteins missing between Cm and other spliceosomes (Fig. 6). Besides the absent proteins already discussed, the Cm B complex lacks U2 snRNP-associated proteins Lea1, Msl1, SF3B6, Ysf3, and the B complex-specific proteins FBP21, PPIH, Snu23, Snu66, Smu1, and Spp381 (Fig. 6). Similar to U5-40K, the peptidyl-prolyl isomerase PPIH, Brr2-regulator FBP21 and alternative splicing factor Smu1 are conserved among metazoans, but are not present in yeast (Agafonov et al. 2011; Henning et al. 2017; Keiper et al. 2019). In humans, Smu1 and RED are required for spliceosome activation when the distance between the 5′ splice site and branchpoint sequence is short (50 nt), and for splicing efficiency in general (Keiper et al. 2019). In Cm, almost all introns have more than 50 nt between the 5′SS and the BPS.

Surface representation of the Cm B complex homology modeled against the human structure (PDB 5O9Z). All protein sequences were annotated by conservation with their Cm homologs. Identical and similar amino acids are colored purple while dissimilar amino acids are colored wheat. Proteins absent in Cm are shown in transparent gray. The RNA is colored as follows: U6 (dark red); U4 (yellow); U5 (green); U2 (blue); and pre-mRNA (cyan).
The heterodimer Lea1/Msl1 has been implicated in cotranscriptional splicing of a subset of yeast genes (Gunderson and Johnson 2009). Specifically, histone acetylation of actively transcribed genes by the histone acetyltransferase (HAT) Gcn5 promotes the recruitment of Lea1/Msl1, and in turn the U2 snRNP (Gunderson and Johnson 2009). This raises the question of whether or not pre-mRNA splicing occurs cotranscriptionally in Cm. While there is no evidence for Lea1/Msl1 in Cm (Stark et al. 2015), the HAT Gcn5 may exist, as it was the Reciprocal Best Hit for Hypothetical Protein CMA025C. With both U2AF35 and the Lea1/Msl1 heterodimer missing, we predict that splicing and transcription are uncoupled in C. merolae.
The tri-snRNP protein Snu66 has a conserved “HIND” element, and its interaction with the ubiquitin-like protein Hub1 is necessary for alternative splicing and for recognition of noncanonical 5′SS in humans and yeast (Wilkinson et al. 2004). With preliminary RNA sequencing data, we have not found evidence of alternative splicing of the single Cm gene that contains more than one intron, nor have we found evidence for noncanonical splice sites (Stark et al. 2015). These observations are consistent with the absence of Snu66.
When looking at features associated with the RNA core, the Cm B complex is relatively conserved (Fig. 7). Cm Hsh155 and the LSm ring that are proximal to the proposed branch duplex show strong sequence conservation (Fig. 7). In addition, the Prp8 amino-terminal domain is conserved, where the U6 snRNA ACAGAGA helix tethers the 5′SS prior to the first transesterification (Fig. 7, bottom right).
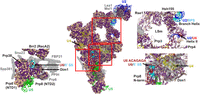
Overview of the Cm B complex homology model (PDB 5O9Z). All protein sequences were annotated by conservation with their Cm homologs. Identical and similar amino acids are colored purple while dissimilar amino acids are colored wheat. Right insets show regions of high conservation, which are also positions of helices formed by U2 snRNA and the BPS (upper right), as well as U6 snRNA and the 5′ splice site (lower right). (Left inset) Surface representation of Prp38 and associated proteins in the Cm B complex (PDB 5O9Z). Cm Prp38 lacks Prp38 complex-associated proteins FBP21, Spp381 and Snu23. Proteins absent in Cm are shown in transparent gray. The RNA is colored as follows: U6 (dark red); U4 (yellow); U5 (green); U2 (blue); and pre-mRNA (cyan).
Snu23 and Spp381, absent in Cm, are part of the Prp38 complex that stabilizes the catalytic RNA prior to the branching reaction (Fig. 7, left inset). Prp38 is an essential splicing protein necessary for formation of the activated Bact complex in humans (Schütze et al. 2016), yeast (Xie et al. 1998), and fruit flies (Andersen and Tapon 2008). Snu23, Spp381, and Prp38 form a trimer that is tri-snRNP-specific in yeast and B complex-specific in animals, which suggests these proteins coevolved (Ulrich and Wahl 2017). The interface between Prp38 and the Prp8 amino-terminal domain reveals those interacting surfaces are well conserved, the Spp381 interacting surface is partially conserved, and the Snu23 surface is not conserved (Fig. 7, left inset). The Prp38-interacting interface of the Prp8 amino-terminal domain is conserved in Cm highlighting the importance of Prp38 in stabilizing the U6 snRNA ACAGAGA helix formed with the 5′SS during the activation step in Cm (Fig. 7, left inset; Supplemental Fig. 1O; Plaschka et al. 2017).
C. merolae Bact complex
In yeast and humans, transition from the B complex to the activated Bact complex requires departure of the U1 and U4 snRNPs, recruitment of the NTC and related proteins, and formation of the U6 snRNA intramolecular stem–loop (ISL) and U2/U6 Helix I. These structures, together with loop I of U5 snRNA, form the RNA center of the spliceosome that coordinates catalytic Mg2+ ions responsible for the actual chemistry of splicing. The formation of the active site occurs in the activated spliceosome or Bact complex. This complex is not yet chemically competent, because the BPS is too distant from the 5′SS for splicing chemistry to occur (Rauhut et al. 2016; Yan et al. 2016).
Cm lacks 16 of the 38 proteins of the yeast Bact complex (Table 1; Fig. 8). The majority of the proteins missing in the Cm Bact complex are non-snRNP or otherwise associated with the NTC-, RES-, or Bact complexes (Fig. 8; Bao et al. 2017a). Notably, the Cm NTC is missing Prp17 and the Prp17 yeast synthetic lethal proteins Syf1 and Syf2, Ecm2, Snt309 (SPF27), Prp45, Cwc2, Cwc25, and peptidyl-prolyl isomerases PPIL1 and PPIE (Fig. 8). Syf1, but not Syf2, is required for cell cycle progression and splicing activity in yeast (Russell et al. 2006). In Cm, Syf1, Syf2, and their interacting partner Prp17 are all absent, which suggests these proteins function cohesively and perhaps coevolved (Russell et al. 2006). In yeast, Syf1, Syf2, Ecm2, Cwc2, and Prp17 are connected to each other in a chain-like fashion, as well as to the core of the Bact complex (Fig. 8). These missing proteins surround helix I of the yeast U2/U6 snRNA duplex, which is therefore presumably exposed in Cm (Fig. 8, right). In contrast, the Prp8 N-domain that coordinates the branch helix formed by the 3′ end of U2 snRNA and the intron branchpoint sequence is highly conserved (Fig. 8, right). Cm lacks Cwc2, which contacts the U6 snRNA internal stem–loop (ISL) and is involved in making the active site catalytically competent (Rasche et al. 2012). The absence of the cotranscriptional splicing factor Prp45 (Hálová et al. 2017) is reminiscent of the loss of the Lea1/Msl1 heterodimer, because they are both associated with histone modification-dependent recruitment of splicing machinery to actively transcribed genes (Gunderson and Johnson 2009; Leung et al. 2019). Prp45, and subsequently the NTC, is cotranscriptionally recruited to the precatalytic B complex by the chromodomain protein Eaf3 (Leung et al. 2019). Eaf3 interacts with both the Set2-trimethylated histone H3K36 as well as Prp45 (Leung et al. 2019).

Surface representation of the Cm Bact complex homology modeled against the Sc structure (PDB 5GM6). All protein sequences were annotated by conservation with their Cm homologs. Identical and similar amino acids are colored purple while dissimilar amino acids are colored wheat. Proteins absent in Cm are shown in transparent gray. The RNA is colored as follows: U6 (dark red); U5 (green); U2 (blue); and pre-mRNA (cyan).
Several proteins that interact with Prp45 in the Bact complex and subsequent structures are also absent in Cm including Ecm2 and Syf2 of the NTC and the RES complex (Fig. 8, right; Yan et al. 2016). The RES complex (Bud13, Pml1, and Snu17) is dispensable for splicing, but RES-depleted yeast have inefficient Bact complex assembly and splicing activity (Bao et al. 2017b). The activated Cm spliceosome also lacks the NTC-associated heat resistant protein Cwc25, which, together with Yju2, stabilizes the U2 snRNA/branchpoint helix during the branching reaction (Chiang and Cheng 2013; Zhan et al. 2018a). While Cwc25 and Yju2 are both essential in yeast, only Yju2 has been identified in Cm (Supplemental Fig. 1R).
The simplified Cm spliceosome offers a glimpse into essential spliceosomal components (Stark et al. 2015). Cm lacks the cotranscriptional pre-mRNA splicing regulation offered by either Lea1/Msl1 (Gunderson and Johnson 2009) or Prp45 (Hálová et al. 2017; Leung et al. 2019), consistent with its inefficient splicing (Stark et al. 2015; Schärfen et al. 2022). We assume the first transesterification is inefficient in Cm based on the splicing factors that are absent in the activated Cm spliceosome, particularly Prp17 and NTC proteins.
Absent Ysf3 protein and destabilization of SF3B in C. merolae
The SF3B complex in the U2 snRNP is responsible for catalytic latency, i.e., for preventing premature nucleophilic attack of the chemically reactive hydroxyl of the branchpoint adenosine (McPheeters and Muhlenkamp 2003). Comparison of the homology with the electrostatic surface potential of the SF3B complex suggests that the important RNA-binding surfaces (electropositive) are conserved in Cm (Fig. 9A,B). Six proteins fold into the compact SF3B structure (Rse1, Hsh155, Cus1, Rds3, Hsh49, and Ysf3). Ysf3 (SF3B5) is a wedge-like protein that is missing in the Cm spliceosome (Fig. 9A). In yeast, Ysf3 is essential for U2 snRNP entry during spliceosomal assembly and splicing (Wang et al. 2005). The SF3B complex can assemble without Ysf3, but loss of Ysf3 causes reduction in Cus1 and a decrease in stability of the SF3B complex (Wang et al. 2005). Ysf3 absence may result in slower splicing, but may also obviate the need for proteins that would otherwise be required to help remove the SF3B complex such as the Prp2 cofactors Cwc22 and Spp2 (Silverman et al. 2004; Yeh et al. 2011), which are indeed absent in Cm.
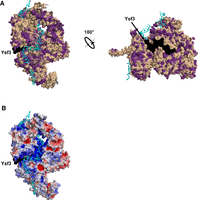
The SF3B complex from the Sc Bact complex (PDB 5GM6), showing both (A) Cm homology model and (B) electrostatic surface representations. Ysf3, shown in black, is absent in the Cm SF3B complex. In A, all protein sequences were annotated by conservation with their Cm homologs. Identical and similar amino acids are colored purple while dissimilar amino acids are colored wheat. mRNA is colored cyan. In B, the protein surface is colored from blue (electropositive) to red (electronegative) with mRNA as in A.
Spp2/GPKOW is a G-patch protein that activates the DEAH-box helicase Prp2/DHX16 (Robert-Paganin et al. 2015). There are five known G-patch proteins that activate DEAH-box helicases in both yeast and humans: Cgm1/GPATCH11, Gno1/PINX1, Pfa1/NKRF, Spp382/TFIP11, and Spp2/GPKOW (Robert-Paganin et al. 2015; Studer et al. 2020). Of those five, Spp2/GPKOW activates Prp2/DHX16 (Warkocki et al. 2015), while the other four all activate Prp43/DHX15 (Hamann et al. 2019; Bohnsack et al. 2021). Mass spectrometry and bioinformatics suggest there do not appear to be any DEAH-box helicase-activating G-patch proteins present in Cm. The absence of G-patch activators of DEAH-box helicases suggests that Cm lacks regulation and efficiency during ATP-dependent conformational changes.
In humans, the U2 protein p14/SF3B6, a component of the SF3B complex, associates with the BPS at early stages of spliceosome assembly and directly contacts the BPS adenosine (MacMillan et al. 1994; Schellenberg et al. 2006; Tholen et al. 2022); this protein is also missing in Cm. It is also notably absent in yeast (Dziembowski et al. 2004) despite the presence of the six other protein components of SF3B, but is conserved in other fungi including the fission yeast S. pombe. Overall, the presence of SF3B6 appears to correlate with degeneracy of the intron BPS (the BPS in both Cm and yeast is highly conserved while that in S. pombe is of intermediate complexity compared to the highly degenerate human BPS). Thus, SF3B6 may play a role in the selection of the BPS as a splicing regulatory mechanism.
C. merolae Prp8 lacks structural motifs necessary for splicing efficiency
The human and yeast Prp8 RNase H domain appears to act as a spliceosomal throttle. It is hypothesized to toggle assembly between the “closed” and “open” catalytic states associated with hyper-accurate/slow and inaccurate/fast splicing, respectively (Schellenberg et al. 2013; Mayerle et al. 2017). It contains a 17 amino acid insertion between two adjacent β-strands termed the “Toggle loop” motif (Schellenberg et al. 2013; Mayerle et al. 2017). This loop motif is not conserved in Cm; absence of the 17 amino acid insertion was suggested bioinformatically and confirmed crystallographically (Garside et al. 2019). In place of the Toggle loop, the Cm Prp8 RH domain contains a well-ordered type II β-turn that locks the RH domain in the “Closed loop” conformation (Garside et al. 2019). The Closed loop conformation of the Cm Prp8 RH domain, combined with the absence of the Toggle loop and other regulatory proteins previously discussed, leads us to predict the Cm spliceosome is slow during assembly, transitional, and catalytic stages.
Similar to the observation with Sad1, the loss of the RH Toggle loop motif is associated with the loss of proteins that interact with it such as Prp6 in the tri-snRNP. Along with the RNase H domain Toggle loop, Cm Prp8 is also predicted to be missing the “Switch loop” of its linker domain (Supplemental Fig. 1J). In the structure of the yeast Bact complex, the Switch loop interacts with Cwc21, which is also missing in Cm (Jia and Sun 2018; Supplemental Fig. 1J). The absence of these two features from the Cm Bact complex active site is predicted to translate to fewer and weaker interactions with the 5′SS prior to catalysis. Thus, splicing accuracy in Cm is also predicted to be lower than in highly regulated spliceosomes. The absence of these two features would also mean that a canonical exon binding channel would not exist in Cm (Figs. 10, 11).
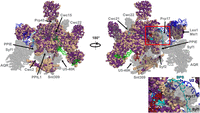
Surface representation of the Cm C* complex homology modeled against the human structure (PDB 5MQF). All protein sequences were annotated by conservation with their Cm homologs. Identical and similar amino acids are colored purple while dissimilar amino acids are colored wheat. (Inset) Several proteins are missing or are poorly conserved within the Cm 3′SS/BPS region, highlighting the importance of CmPrp8 in the second transesterification. Proteins that are absent in Cm are shown in transparent gray. The RNA is colored as follows: U6 (dark red); U5 (green); U2 (blue); and pre-mRNA (cyan).
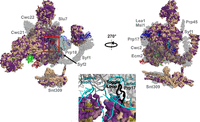
Surface representation of the Cm P complex homology modeled against the Sc structure (PDB 5YLZ). All protein sequences were annotated by conservation with their Cm homologs. Identical and similar amino acids are colored purple while dissimilar amino acids are colored wheat. (Inset) the intron lariat junction from the Sc P complex active site (PDB 5YLZ). The 1585 loop (present in CmPrp8) and Toggle loop (missing in CmPrp8) are represented in cartoon, while the respective linker and RNase H domains of CmPrp8 are represented as surfaces. Proteins absent in Cm are shown in transparent gray. The RNA is colored as follows: U6 (dark red); U5 (green); U2 (blue); and pre-mRNA (cyan).
Despite the lack of protein coordination, we predict there are novel, compensatory RNA–RNA interactions that contribute to splicing in Cm. Cm U5 snRNA is 450 nt in length, which is much larger than both the 214 nt yeast U5 snRNA and the 116 nt human equivalent. Secondary structure prediction of Cm U5 snRNA suggests several additional stems and loops are present (Fig. 4). It is possible that Cm U5 snRNA stabilizes the 5′ exon during the first transesterification in a role otherwise filled by the Prp8 Switch loop (Stark et al. 2015).
C. merolae C* complex
The catalytically activated C complex (C*) is formed after the ATP-driven DEAH-box helicase activity of Prp16 results in dissociation of first step factors and rearrangement of the branched intron for the second transesterification (Chiang and Cheng 2013). Cm lacks a third of the proteins (15) present in the cryo-EM structure of the human C* complex (Fig. 10).
The majority of proteins missing from the C* complex—such as Cwc2, Cwc21, Cwc22, Lea1, Msl1, U5-40K, Prp45, Snt309, Syf1, and Syf2—are not associated with a specific snRNP and have been discussed previously (Fig. 10). Some of these proteins are functionally relevant earlier in the Bact complex and the first transesterification reaction, including the post-first step branched complex. Each of these missing proteins plays an important role in structures of human or yeast C*. Msl1 and Lea1 are part of a bridge between stem IV of U2 snRNA and proteins interacting with the active site (Price et al. 1998; Galej et al 2016). The splicing factor Cwc21 binds and stabilizes the 5′ exon along with portions of Prp8 (Bertram et al. 2017a; Fica et al. 2017). The NTC protein Syf2 binds to Helix II of the annealed U2/U6 snRNA as well as to Prp45 and Clf1 (Bertram et al. 2017b). Thus, the catalytic site of the spliceosome is enveloped by PRP45/NTC components as well as elements of Prp8 (Galej et al. 2016). The Prp18–Slu7 complex that joins the spliceosome for the second transesterification is also absent in Cm, as is Cwc22 that would contact Slu7 at this stage (Fica et al. 2017; Zhang et al. 2017). Prp18, Slu7, and Prp22 are required for efficient positioning of the 3′SS at the catalytic center, and displace first step factors Cwc25 and Yju2 in yeast (Schwer 2008). The absence of Msl1, Lea1, and Prp45 in Cm suggests that stabilization of spliceosomal assembly intermediates may involve the participation of more complex RNA structures or elements of proximal, conserved core proteins.
Based on the above analysis, the transition from the C complex to C* would presumably be less complex in Cm; Yju2 would either depart or undergo a conformational rearrangement upon Prp22 entry, but there would be no exchange between Cwc25, Prp18, or Slu7. This last factor is unnecessary in exon ligation events where the distance between the BPS and 3′SS is 9 nucleotides or fewer (Frank and Guthrie 1992; Brys and Schwer 1996; James et al. 2002); 18 of the 38 Cm introns contain 20 nucleotides or more between the BPS and 3′SS, so the absence of Slu7 is surprising. It has been reported that splicing occurs at a low efficiency in yeast extracts depleted of Cwc25, Prp18, and Slu7 (Tseng et al. 2017). Prp17 is also absent from the Cm C* complex and might cause the U2 snRNA/branchpoint helix to be relatively less stable (Fig. 10). The majority of proteins at the catalytic center that contact snRNAs, such as Cdc5 and Crn, are present in the Cm C* spliceosome (Fig. 10). Interestingly, the amino acid conservation between these proteins is not high; perhaps their structures remain conserved while their sequence-specific RNA recognition does not (Fig. 10).
C. merolae P complex
Recent structures of yeast P complexes have provided several snapshots immediately following the transesterification reaction that results in exon ligation (Bai et al. 2017; Liu et al. 2017; Wilkinson et al. 2017; Fica et al. 2019). The yeast P complex contains approximately 40 proteins (Fica et al. 2019) of which only 15 have orthologs that have been identified in Cm (Fig. 11). Of the 16 proteins that stabilize RNA elements at the P complex active site, Cm is missing 10 (Table 1).
A recurring theme in the analysis of the Cm spliceosome is that most of the proteins absent in the Cm P complex are peripheral to the RNA helices or stabilize the active site (Fig. 11). We hypothesize that the proteins present in the Cm P complex represent an essential catalytic core of proteins and RNA necessary for exon ligation. These include Prp8, Snu114, Cef1, Clf1, Bud31, and Prp46. With the exception of Bud31, these are all among the phylogenetically most highly retained splicing factors. Many of the missing proteins in the C* complex are of course absent in the Cm P complex, such as the intron-binding NTR protein Cwc2 or the NTC component Syf2, which stabilizes the U2/U6 snRNA Helix II (Rasche et al. 2012; Bertram et al. 2017b). Cwc21 and Cwc22, also absent, stabilize the 5′ exon during and after ligation (Yeh et al. 2011; Chen et al. 2013; Bai et al. 2017; Bertram et al. 2017b). The absence of Prp17 and the Prp8 Toggle loop motif could have substantial effects on the stability of the spliceosome active site in the C* and P complexes (Fig. 11 inset; Sapra et al. 2008; Mayerle et al. 2017). Some of the missing proteins modulate the efficiency of splicing as described for Prp18 and Slu7.
DISCUSSION
Our analysis of yeast and human spliceosome structures through a C. merolae lens reveals an interesting trend, namely the increased number of missing proteins as a function of step in the splicing cycle (Fig. 2). Most of the proteins present in the catalytic core of the spliceosome are present in Cm with the exception of Prp17 (Figs. 10, 11). An in-depth look at the catalytic centers of these spliceosomes shows that as assembly and catalysis progress in Cm, more proteins and structural motifs are missing at each step. Interestingly, some of these absent proteins and domains, such as the Prp18–Slu7 heterodimer, or the Toggle loop of the Prp8 RNase H domain, are proposed to act as throttles that normally allow the spliceosome to shift between two states: slow and accurate versus fast and inaccurate. The absence of such domains is consistent with the low fraction of spliced transcripts observed in Cm (Stark et al. 2015; Schärfen et al. 2022), although we have not observed evidence of inaccuracy: while some cryptic splice site use is detected by Illumina and PacBio sequencing, there do not appear to be more of these in Cm than in other organisms (Schärfen et al. 2022).
We have observed an absence of proteins or motifs that are coupled: Sad1 and the Brr2 plug domain during tri-snRNP incorporation (Fig. 5, inset); Cwc21 and the Prp8 RH Toggle loop during spliceosome activation; and Cwc22 and Slu7 in the C* complex. In the ligation reaction, Prp17 and the Prp8 RH Toggle loop interact with each other to stabilize the 3′SS and are both missing in Cm (Fig. 11). The Prp8 RH Toggle loop motif, together with the 1585 loop/α-finger motif of the Prp8 linker domain, sandwiches the intron lariat/active site in the yeast P complex (Fig. 11). The most surprising protein absences from the Cm spliceosome are those from the active site, such as Cwc21 and Prp17, as well as cotranscriptional regulatory factors Lea1/Msl1 and Prp45.
Pairwise and multiple sequence alignments of the Cm proteins suggest many proteins are either missing canonical domains or contain novel ones. Prp19 is dramatically truncated, lacking the amino-terminal RING domains and the carboxy-terminal WD40 motifs (Supplemental Fig. 2D). The Toggle loop motif of the Prp8 RNase H domain is another example (Fig. 11; Supplemental Figs. 1J, 2A; Mayerle et al. 2017). Conversely, CmMud2 has a large amino-terminal extension compared to its human and yeast orthologs and is ∼800 aa longer (Supplemental Fig. 1F), and the U5 snRNA is nearly twice as large as its homologs in other organisms (Fig. 4).
Homology-based annotation of human and yeast structures revealed that the reduced C. merolae spliceosome lacks protein–RNA and protein–protein interactions that together suggest less coordination of RNA elements and regulation of protein activity during the splicing cycle. We suggest Cm splicing—and by analogy that of other organisms with reduced spliceosomes—to be inefficient, inaccurate, and post-transcriptional. The inefficiency of Cm splicing contrasts strongly with splicing efficiency in other organisms including yeast (Stark et al. 2015; Schärfen et al. 2022). Furthermore, Cm lacks the DEAD-box RNA helicase Prp28, which is not only associated with release of U1 snRNP, but also with proofreading the 5′SS and splicing fidelity (Price et al. 2013). These properties of Cm splicing may be more similar to an ancestral (pre-LECA) spliceosomal ribozyme—with few associated proteins—than to the highly elaborated, protein-rich spliceosomes present in animals and plants.
The discovery of a spliceosome lacking the U1 snRNP was surprising. The comparative analysis performed here will be useful to the extent that it stimulates questions about the precise role of each splicing factor and the evolutionary pressures that have led to the great divergence in splicing systems seen across organisms. Much mechanistic work is still required before we understand the many layers of splicing regulation and splice site selection.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
We acknowledge Kevin Huolt for initial development of algorithms used to color structures by sequence conservation. We thank Dr. Martha Stark for helpful feedback on the manuscript. This work was supported by Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery grants to S.D.R., N.M.F., and A.M.M.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079273.122.
-
Freely available online through the RNA Open Access option.
This article, published in RNA, is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵








