
Thermodynamic control of mismatch discrimination for extensive splicing regulation of PKM pre-mRNA
- Department of Nucleic Acids Bioengineering, Institute of Bioorganic Chemistry, Polish Academy of Sciences, 61-704 Poznan, Poland
- Corresponding authors: nbartys{at}ibch.poznan.pl, apa{at}ibch.poznan.pl
-
Handling editor: Michelle Hastings
Abstract
In this article, we present an approach to maximizing the splicing regulatory properties of splice-switching oligonucleotide (SSO) designed to regulate alternative splicing of pyruvate kinase M (PKM) pre-mRNA. The studied SSO interacts with the regulatory element in exon 10 of PKM pre-mRNA and contributes to a significant reduction of PKM2 level with a simultaneous increase of the PKM1 isoform. This SSO forms a duplex not only with the regulatory fragment of exon 10 but also with a similar RNA fragment of intron 9. The impact of this unspecific interaction on SSO regulatory properties, as well as the functional role of the intron 9 fragment, remains debatable. Herein, two types of modified nucleotides, unlocked nucleic acids (UNAs) and locked nucleic acids (LNAs), were used to study mismatch discrimination within duplexes involving modified SSOs. Our studies showed that LNAs increased mismatch discrimination, resulting in stronger regulatory properties of SSO. On the other hand, UNA reduced mismatch discrimination, decreasing the potentially therapeutic properties of SSO. The results indicate that specific interactions with exon 10 are more favorable for therapeutic applications than simultaneous hybridization with intron 9 and exon 10. The results also suggest the lack of a regulatory role for the intron 9 target site in alternative splicing of PKM pre-mRNA.
Keywords
INTRODUCTION
Splice-switching antisense oligonucleotides (SSOs) are 15–30 nt long molecules designed to interact with pre-mRNA through complementary base-pairing and alter pre-mRNA splicing (Bauman et al. 2009). SSOs function by base-pairing with regions of an RNA that is required for splicing. This hybridization can prevent the binding of splicing factors to their target sites, leading to altered splice sites recognition and a change in the alternative splicing pattern of target pre-mRNA. Additionally, SSO hybridization might affect the RNA secondary structure, which in turn can alter the accessibility of splice sites or splice regulatory elements for binding by splicing factors (Peacey et al. 2012; Singh et al. 2015; Havens and Hastings 2016). One of the challenges in the design of SSO is identifying the key pre-mRNA fragments (i.e., splice sites, cryptic splice sites, splicing regulatory elements) responsible for the splicing regulation of target gene. The potential of SSOs was largely realized following the commercial success of nusinersen, a groundbreaking SSO that is used to treat spinal muscular atrophy (SMA) patients (Chiriboga 2017). Moreover, four other SSO therapeutics targeted toward exon 45, exon 51, and exon 53 have been approved for dystrophin gene alternative splicing regulation (Egli and Manoharan 2023). There is an increasing number of studies on SSOs applications in the treatment of various diseases caused by dysregulation of an alternative splicing process, including cancer (Ceccarello et al. 2021; Balachandran et al. 2023; Chen et al. 2024).
Pyruvate kinase M (PKM) is an enzyme involved in the conversion of phosphoenolpyruvate to pyruvate in the glycolysis process. In the PKM pre-mRNA, there are two mutually exclusive exons (9 and 10) that are selected during the alternative splicing process to produce two different isoforms, PKM1 and PKM2, with exons 9 and 10, respectively (Fig. 1A; Takenaka et al. 1991). The PKM2 isoform is thought to contribute to cancer development and progression (Hsu and Hung 2018). Therefore, there are attempts to redirect the splicing of the PKM gene in cancer cells to reduce the expression of the PKM2 isoform and to increase the level of PKM1 (Wang et al. 2012; Ma et al. 2022). In 2012, the Krainer research group published the sequence of an SSO that can redirect splicing of PKM pre-mRNA with high efficiency (Wang et al. 2012). The oligonucleotide hybridizes to a strong regulatory element located in exon 10 of PKM pre-mRNA (Fig. 1B). The authors demonstrated the proapoptotic influence of the SSO, which was related to the reduction of PKM2 level. However, analysis of the PKM pre-mRNA sequence revealed that in intron 9, an RNA fragment with a sequence very similar to the exon 10 regulatory motif is present (Fig. 1B). Consequently, the 15 nt long SSO forms a full complementary duplex with the exon 10 fragment, but also might hybridize with intron 9, forming a duplex containing four mismatches (Fig. 2A,B). The similarity of these sequences suggests that both can promote regulatory function. The authors presented the theory that the effectiveness of the SSO is related to simultaneous hybridization with exon 10 and intron 9, although the regulatory effect mainly derives from the exon 10 interaction, with the effect partially supported by hybridization to intron 9 (Fig. 1B).
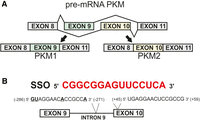
(A) Mutually exclusive alternative splicing of PKM gene. (B) PKM pre-mRNA sequences with potential SSO-binding sites.
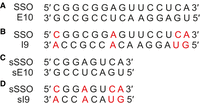
The sequence of duplexes formed by SSO with exon 10 (A) or intron 9 (B) fragments. Duplexes formed by sSSO applied in thermodynamic studies constitute shortened models of SSO interactions with exon 10 (C) and intron 9 (D). The red font indicates mismatch positions in SSO/I9 and sSSO/sI9 duplexes.
One challenge in antisense oligonucleotide (ASO) design is the off-target effect caused by unintended hybridization of the
ASO with other RNA fragments of high sequence similarity. Off-target interaction is a serious problem in therapeutic approaches
and causes toxicity of the drug, simultaneously decreasing its specificity of action (Yoshida et al. 2019; Scharner et al. 2020). To achieve the regulatory effect caused by steric-blocking oligonucleotides like SSOs, the RNA fragment with regulatory
function must be a target of hybridization. Therefore, the off-target effects caused by SSOs are less common than those caused
by other types of oligonucleotides that act via degradation mechanism. Nonetheless, the unspecific SSO hybridization limits
effective SSO concentration and weakens therapeutic effect. The sequence similarity of the fragments within exon 10 and intron
9 of PKM pre-mRNA suggests a high probability of off-target interactions leading to SSO activity reduction. In our studies, we aimed
to increase the therapeutic potential of SSO and simultaneously verify the hypothesis about the regulatory function of intron
9. For this purpose, we analyzed the thermodynamic parameters of duplexes formed by the hybridization of SSO with exon 10
and intron 9 fragments. It is expected that hybridization with complementary fragments is more energetically favorable and
that such a duplex is more stable than a mismatched structure. This tendency is manifested by lower Gibbs free energy (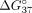 kcal/mol) of a complementary duplex. The presence of mismatches in a duplex with intron 9 results in decreased duplex stability,
less favorable
kcal/mol) of a complementary duplex. The presence of mismatches in a duplex with intron 9 results in decreased duplex stability,
less favorable 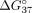 , and, at the same time, a lower probability of hybridization. The preference of an oligonucleotide to hybridize with a complementary
fragment instead of a mismatched one is named mismatch discrimination.
, and, at the same time, a lower probability of hybridization. The preference of an oligonucleotide to hybridize with a complementary
fragment instead of a mismatched one is named mismatch discrimination.
Application of various modified nucleotides opens the possibility of manipulating the thermodynamic stability of duplexes influencing the hybridization preferences of SSOs. In our studies, we used two well-known chemical modifications of nucleotides: unlocked nucleic acids (UNAs) and locked nucleic acids (LNAs) (Fig. 3). UNA nucleotides lack the bond between C2′ and C3′ atoms in the ribose residue (Fig. 3B). The characteristic impact of UNA on duplexes is the reduction of their thermodynamic stability (Langkjær et al. 2009; Pasternak and Wengel 2010, 2011). The LNA nucleotides have an additional methylene bridge between the C4′ and the O2′ atoms of the ribose residue (Fig. 3A). Unlike UNA residues, LNAs increase the thermodynamic stability of duplexes (You et al. 2006; Campbell and Wengel 2011; Owczarzy et al. 2011). Our efforts were focused on their application at oligonucleotide positions that form mismatches with intron 9. The manipulation of mismatch discrimination might influence the preferred oligonucleotide hybridization and predictably change the regulatory effect. We expected that the reduction of mismatch discrimination might facilitate understanding of the previously postulated regulatory role of the intron 9 fragment as well as optimization of the efficiency of a potentially therapeutic SSO. The thermodynamic parameters of unmodified and modified duplexes were determined using the UV melting method. Due to the high stability of the complementary 15 bp long duplex that hinders obtaining reliable thermodynamic data, we synthesized shorter 8-nt long variants of SSO (sSSO) (Fig. 2C,D). Importantly, all mismatches formed between the model oligonucleotides and shortened I9 fragment (sI9) are of the same type, at the same positions, and neighbored by the same base pairs as in the 15 bp long duplex (Fig. 2C,D). All oligonucleotides were composed of a 2′methoxyethyl nucleoside residues (2′MOE) and phosphorothioate backbone.
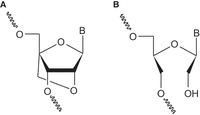
The structures of modified nucleotide residues, LNA (A) and UNA (B).
RESULTS AND DISCUSSION
The influence of LNA and UNA residues on duplex thermodynamic stability
Oligonucleotide residues that form mismatches with the intron 9 fragment were modified with UNA or LNA modifications. Each
modified residue was introduced separately, giving the set of eight modified oligonucleotides (sSSO1–sSSO8, Table 1). Thermodynamic parameters were defined for the hybridization of each modified oligonucleotide with shortened exon 10 (sE10)
and intron 9 (sI9) fragments. To describe the change in thermodynamic stability, we calculated 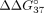 according to the following equation:
according to the following equation: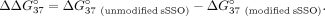
The oligonucleotides applied in thermodynamic studies
Performed thermodynamic studies allowed assessment of the influence of modified nucleotides on the thermodynamic stability
of sSSO/sE10 and sSSO/sI9 duplexes. A comparative analysis of the enthalpy and entropy values determined by the two calculation
methods shows that all duplexes unfolded according to the two-state model (Supplemental Tables 1 and 2). Figure 4 shows the free energy change (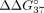 ) between the unmodified duplex sSSO/sE10 or sSSO/sI9 and the modified duplexes of the same type, whereas Tables 2 and 3 present the thermodynamic data. Values of
) between the unmodified duplex sSSO/sE10 or sSSO/sI9 and the modified duplexes of the same type, whereas Tables 2 and 3 present the thermodynamic data. Values of 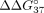 less than 0 indicate thermodynamic stabilization of the duplex, whereas values greater than 0 correspond to duplex destabilization.
The full thermodynamic data can be found in the Supplemental Material.
less than 0 indicate thermodynamic stabilization of the duplex, whereas values greater than 0 correspond to duplex destabilization.
The full thermodynamic data can be found in the Supplemental Material.
The free energy changes (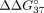 ) of duplexes formed by exon 10 fragment sE10 (A) and intron 9 fragment sI9 (B) with modified sSSOs.
) of duplexes formed by exon 10 fragment sE10 (A) and intron 9 fragment sI9 (B) with modified sSSOs.
Thermodynamic parameters of LNA-modified sSSO/RNA duplexes
Thermodynamic parameters of UNA-modified sSSO/RNA duplexes
A single LNA residue increases the thermodynamic stability of all sSSO duplexes formed with sE10 as well as with sI9, with
the exception of the sSSO2/sI9 duplex (Fig. 4; Table 2). However, this effect is larger for the sSSO/sE10 duplex, in which there is full base pair complementarity. The highest
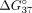 change was observed for the sSSO2/sE10 duplex. The presence of LNA-A in the internal position contributed to an increase
in thermodynamic stability (
change was observed for the sSSO2/sE10 duplex. The presence of LNA-A in the internal position contributed to an increase
in thermodynamic stability (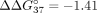 kcal/mol). This effect is consistent with previously published studies for 2′-O-Me-RNA/RNA duplexes, where it was shown that
internal LNA-A stabilizes the double helix by 0.96–1.42 kcal/mol, depending on the neighboring nucleotides (Kierzek et al. 2005). A less significant
kcal/mol). This effect is consistent with previously published studies for 2′-O-Me-RNA/RNA duplexes, where it was shown that
internal LNA-A stabilizes the double helix by 0.96–1.42 kcal/mol, depending on the neighboring nucleotides (Kierzek et al. 2005). A less significant 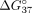 value reduction (
value reduction (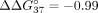 kcal/mol) was observed for the sSSO3/sE10 duplex with LNA-C at the penultimate position of the 3′ end. This is a less pronounced
effect than previously described for LNA-C (
kcal/mol) was observed for the sSSO3/sE10 duplex with LNA-C at the penultimate position of the 3′ end. This is a less pronounced
effect than previously described for LNA-C (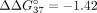 kcal/mol). However, the difference might be due to another type of neighboring nucleotide that surrounds the LNA-C residue
in the two types of duplexes and, consequently, results in different interactions between the residual charges of the functional
groups of the heterocyclic bases. LNA-C, which is present at the 5′ terminal position, increases sSSO1/sE10 duplex stability
by 1.11 kcal/mol. A slightly stronger stabilization was observed for the sSSO4/sE10 duplex with LNA-A at the 3′ end. Thus,
when inserting LNA residues into terminal fragments of the duplex, a marginally larger stabilizing effect is obtained by modifying
the 3′ end of an sSSO. A similar conclusion is confirmed in the literature, although the stabilizing effect of the LNA-C residue
at the 5′ end of sSSO1 was found to be stronger than the effects previously described for LNA residues (
kcal/mol). However, the difference might be due to another type of neighboring nucleotide that surrounds the LNA-C residue
in the two types of duplexes and, consequently, results in different interactions between the residual charges of the functional
groups of the heterocyclic bases. LNA-C, which is present at the 5′ terminal position, increases sSSO1/sE10 duplex stability
by 1.11 kcal/mol. A slightly stronger stabilization was observed for the sSSO4/sE10 duplex with LNA-A at the 3′ end. Thus,
when inserting LNA residues into terminal fragments of the duplex, a marginally larger stabilizing effect is obtained by modifying
the 3′ end of an sSSO. A similar conclusion is confirmed in the literature, although the stabilizing effect of the LNA-C residue
at the 5′ end of sSSO1 was found to be stronger than the effects previously described for LNA residues (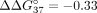 ÷ −0.59 kcal/mol) at the 5′ terminal position (Kierzek et al. 2005). It was shown that a cytidine residue in the LNA series, which is present at the 5′ end of the 2′-O-Me-RNA strand, increases
the thermodynamic stability of the 2′-O-Me-RNA/RNA duplex by 0.40 kcal/mol, whereas an adenosine residue in the LNA series
at the 3′ end increases the thermodynamic stability of the duplex by 1.30 kcal/mol. The stabilizing effect of 3′ terminal
LNA was observed for every modified nucleotide residue except uridine. Furthermore, in the cited studies, it was also found
that the greatest stabilizing effect of the 2′-O-Me-RNA/RNA duplex is obtained by inserting an LNA residue at the central
position of the oligonucleotide. This effect also occurs for each LNA nucleotide residue, except uridine (Kierzek et al. 2005).
÷ −0.59 kcal/mol) at the 5′ terminal position (Kierzek et al. 2005). It was shown that a cytidine residue in the LNA series, which is present at the 5′ end of the 2′-O-Me-RNA strand, increases
the thermodynamic stability of the 2′-O-Me-RNA/RNA duplex by 0.40 kcal/mol, whereas an adenosine residue in the LNA series
at the 3′ end increases the thermodynamic stability of the duplex by 1.30 kcal/mol. The stabilizing effect of 3′ terminal
LNA was observed for every modified nucleotide residue except uridine. Furthermore, in the cited studies, it was also found
that the greatest stabilizing effect of the 2′-O-Me-RNA/RNA duplex is obtained by inserting an LNA residue at the central
position of the oligonucleotide. This effect also occurs for each LNA nucleotide residue, except uridine (Kierzek et al. 2005).
Our studies provide the general conclusion that LNA modification increases the thermodynamic stability of duplexes with the
exon 10 fragment. The LNA residue also influences the thermodynamic stabilization of three sSSO/sI9 duplexes, although to
a lesser extent (Fig. 4B). This effect was expected since the LNA residues are present in mismatch positions of the sSSO/sI9 duplex. The most significant
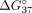 change was noted for sSSO3/sI9 and sSSO4/sI9 duplexes, where
change was noted for sSSO3/sI9 and sSSO4/sI9 duplexes, where 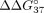 was equal to −0.56 and −0.57 kcal/mol, respectively. In these oligonucleotides, the LNA is present in the penultimate and
the 3′ terminal positions. Interestingly, the sSSO2 oligonucleotide that formed the most stable duplex with sE10 hybridizes
to sI9 with similar thermodynamic stability as unmodified sSSO (Fig. 4B). This effect might be due to LNA-derived reduction of degrees of freedom that impedes adopting a more favorable LNA-A/A
conformation.
was equal to −0.56 and −0.57 kcal/mol, respectively. In these oligonucleotides, the LNA is present in the penultimate and
the 3′ terminal positions. Interestingly, the sSSO2 oligonucleotide that formed the most stable duplex with sE10 hybridizes
to sI9 with similar thermodynamic stability as unmodified sSSO (Fig. 4B). This effect might be due to LNA-derived reduction of degrees of freedom that impedes adopting a more favorable LNA-A/A
conformation.
UNA modifications have an opposing influence on the thermodynamic stability of duplexes to LNA. We observed a decrease in
the thermodynamic stability of all duplexes with shortened fragments of exon 10 (Fig. 4A). In the case of sSSO/sI9 duplexes, UNAs induced various thermodynamic effects (Fig. 4B). The stability of sSSO5/sI9 and sSSO8/sI9 was reduced, but the stability of sSSO7/sI9 increased. Additionally, for sSSO6/sI9,
we noticed no 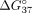 change in comparison to the unmodified duplex. The stronger influence of UNAs was observed more for the complementary duplex
(sSSO/sE10) than for mismatched sSSO/sI9 structures. Interestingly, in the case of UNA modification within the sSSO/sE10 duplex,
we noticed a strong difference between the thermodynamic effect of central UNA residues and terminal ones. The destabilization
effect was the most noticeable for the sSSO6/sE10 duplex containing internal UNA adenosine (
change in comparison to the unmodified duplex. The stronger influence of UNAs was observed more for the complementary duplex
(sSSO/sE10) than for mismatched sSSO/sI9 structures. Interestingly, in the case of UNA modification within the sSSO/sE10 duplex,
we noticed a strong difference between the thermodynamic effect of central UNA residues and terminal ones. The destabilization
effect was the most noticeable for the sSSO6/sE10 duplex containing internal UNA adenosine (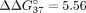 kcal/mol). The UNA-C at the 3′ penultimate position (sSSO7) reduced duplex thermodynamic stability by 3.77 kcal/mol. Both
terminal UNAs had a weaker influence on sSSO/sE10 duplex thermodynamic stability in reference to internal modifications, as
manifested by the 1.23 and 1.27 kcal/mol increase in
kcal/mol). The UNA-C at the 3′ penultimate position (sSSO7) reduced duplex thermodynamic stability by 3.77 kcal/mol. Both
terminal UNAs had a weaker influence on sSSO/sE10 duplex thermodynamic stability in reference to internal modifications, as
manifested by the 1.23 and 1.27 kcal/mol increase in 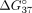 value for the 5′ and 3′ terminal residue, respectively. These results agree with currently available data, which indicate
that the most significant RNA/RNA duplex destabilization might be achieved when a UNA residue is present at the central position.
Moreover, destabilization observed for the sSSO6/sE10 duplex is comparable to the effects previously observed for RNA/RNA
duplexes with a centrally positioned UNA-modified adenosine residue (
value for the 5′ and 3′ terminal residue, respectively. These results agree with currently available data, which indicate
that the most significant RNA/RNA duplex destabilization might be achieved when a UNA residue is present at the central position.
Moreover, destabilization observed for the sSSO6/sE10 duplex is comparable to the effects previously observed for RNA/RNA
duplexes with a centrally positioned UNA-modified adenosine residue (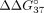 = 4.52 ÷ 5.36 kcal/mol) (Pasternak and Wengel 2010).
= 4.52 ÷ 5.36 kcal/mol) (Pasternak and Wengel 2010).
UNAs, analogically to LNA residues, have a lower impact on the thermodynamic stability of sSSO/sI9 duplexes. It is particularly
interesting that UNA-modified sSSO7 oligonucleotide formed a more stable duplex with sI9 than did unmodified sSSO (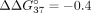 kcal/mol). This is the only position in both types of duplexes where a UNA residue contributes to slight stabilization of
the duplex. However, the UNA-C/U mismatch is adjacent to another nucleotide residue that also forms a mismatch (G/A) instead
of a canonical base pair. Therefore, the presence of a modification that increases the flexibility of the C/U mismatch might
simultaneously relax terminal mismatch within the duplex and facilitate the rearrangement of the structure toward energetically
more favorable interactions. Scientific data provide an example of another UNA-derived stabilization effect observed for the
UNA-U/G mismatch located at the 5′ terminal position (Pasternak and Wengel 2010). Unfortunately, different modified nucleotide positions, as well as different base pair compositions, impede drawing final
conclusions about the origin of the UNA stabilization effect. However, it needs to be considered that this modification, contrary
to expectations, can increase duplex stability in certain conditions. Another unexpected result is the comparable stability
of the sSSO9/sI9 duplex and its unmodified counterpart. In the sSSO9 oligonucleotide, the UNA-A is placed at the internal
position. As described above, this modified position has a strong influence on the thermodynamic properties of the sSSO9/sE10
duplex. However, the duplex with the intron 9 fragment manifests no thermodynamic sensitivity for this modification. This
might indicate that there are no interactions between the two adenosine residues at this position that affect the stability
of the duplex or, more likely, that the local increase in the flexibility of the A/A mismatch does not negatively affect the
interactions between these and neighboring nucleotide residues. This effect might depend on UNA-A neighboring guanosine residues
that form stable pairs with cytidines. In published studies, it was shown that the introduction of UNA-A modification into
the centrally positioned A/A type mismatch destabilizes the duplex by 0.92 kcal/mol (Pasternak and Wengel 2010). However, in the duplex studied by the authors, the A/A mismatch was adjacent to one G-C pair and one U-A pair, so the nearest
neighborhood was different than in our studies.
kcal/mol). This is the only position in both types of duplexes where a UNA residue contributes to slight stabilization of
the duplex. However, the UNA-C/U mismatch is adjacent to another nucleotide residue that also forms a mismatch (G/A) instead
of a canonical base pair. Therefore, the presence of a modification that increases the flexibility of the C/U mismatch might
simultaneously relax terminal mismatch within the duplex and facilitate the rearrangement of the structure toward energetically
more favorable interactions. Scientific data provide an example of another UNA-derived stabilization effect observed for the
UNA-U/G mismatch located at the 5′ terminal position (Pasternak and Wengel 2010). Unfortunately, different modified nucleotide positions, as well as different base pair compositions, impede drawing final
conclusions about the origin of the UNA stabilization effect. However, it needs to be considered that this modification, contrary
to expectations, can increase duplex stability in certain conditions. Another unexpected result is the comparable stability
of the sSSO9/sI9 duplex and its unmodified counterpart. In the sSSO9 oligonucleotide, the UNA-A is placed at the internal
position. As described above, this modified position has a strong influence on the thermodynamic properties of the sSSO9/sE10
duplex. However, the duplex with the intron 9 fragment manifests no thermodynamic sensitivity for this modification. This
might indicate that there are no interactions between the two adenosine residues at this position that affect the stability
of the duplex or, more likely, that the local increase in the flexibility of the A/A mismatch does not negatively affect the
interactions between these and neighboring nucleotide residues. This effect might depend on UNA-A neighboring guanosine residues
that form stable pairs with cytidines. In published studies, it was shown that the introduction of UNA-A modification into
the centrally positioned A/A type mismatch destabilizes the duplex by 0.92 kcal/mol (Pasternak and Wengel 2010). However, in the duplex studied by the authors, the A/A mismatch was adjacent to one G-C pair and one U-A pair, so the nearest
neighborhood was different than in our studies.
The influence of modified nucleotides on mismatch discrimination
The thermodynamic studies allowed the identification of positions that, when modified, significantly influence the thermodynamic
stability of sSSO/sE10 and sSSO/sI9 duplexes. The aim was to manipulate the preference of SSO for hybridization to two PKM pre-mRNA fragments. Therefore, to conclude which positions and modifications can facilitate the most effective redirection
of SSO hybridization, we calculated 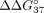 and
and  parameters as follows:
parameters as follows:
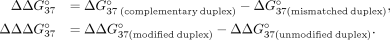
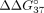 corresponds to the difference in thermodynamic stability between complementary and mismatched duplexes and indicates the
preference of the oligonucleotide to interact with sE10 in reference to sI9.
corresponds to the difference in thermodynamic stability between complementary and mismatched duplexes and indicates the
preference of the oligonucleotide to interact with sE10 in reference to sI9. 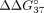 for unmodified oligonucleotide is equal to −6.55 kcal/mol. An increase in this value (
for unmodified oligonucleotide is equal to −6.55 kcal/mol. An increase in this value ( > 0) indicates a decrease in mismatch discrimination. Conversely, the decrease of
> 0) indicates a decrease in mismatch discrimination. Conversely, the decrease of 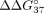 (
( < 0) reflects increased mismatch discrimination. Table 4 presents all the
< 0) reflects increased mismatch discrimination. Table 4 presents all the 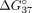 used for calculation and calculated
used for calculation and calculated 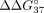 and
and  values for each modified oligonucleotide. The graphical presentation of mismatch discrimination parameters is shown in Figure 5.
values for each modified oligonucleotide. The graphical presentation of mismatch discrimination parameters is shown in Figure 5.
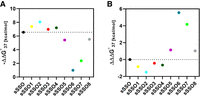
The influence of modified nucleotides on mismatch discrimination. (A) Difference in thermodynamic stability between complementary and mismatched duplex, 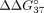 (kcal/mol) (B) mismatch discrimination change,
(kcal/mol) (B) mismatch discrimination change,  (kcal/mol) in reference to unmodified duplex.
(kcal/mol) in reference to unmodified duplex.
Mismatch discrimination properties of modified sSSOs
Our studies show that LNA modification in each mismatch position increases mismatch discrimination (Fig. 5; Table 4). As described above, LNA more significantly increases the thermodynamic stability of sSSO/sE10 than that of sSSO/sI9. Consequently,
the difference in stability (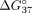 ) between sSSO/sE10 and sSSO/sI9 is elevated. This is in line with increased hybridization of sSSO with a full-complementary
PKM fragment (exon 10). The strongest effect is observed for duplexes with modified central adenosine, for which the hybridization
specificity increases (
) between sSSO/sE10 and sSSO/sI9 is elevated. This is in line with increased hybridization of sSSO with a full-complementary
PKM fragment (exon 10). The strongest effect is observed for duplexes with modified central adenosine, for which the hybridization
specificity increases ( = −1.50 kcal/mol,
= −1.50 kcal/mol, 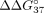 = −8.05 kcal/mol) in reference to the unmodified duplex. It also results in an 11-fold increased hybridization constant for
duplex formation of sSSO2/sE10 versus sSSO2/sI9. A significant increase in mismatch discrimination was also noted for sSSO1-based
duplexes modified with LNA-C (
= −8.05 kcal/mol) in reference to the unmodified duplex. It also results in an 11-fold increased hybridization constant for
duplex formation of sSSO2/sE10 versus sSSO2/sI9. A significant increase in mismatch discrimination was also noted for sSSO1-based
duplexes modified with LNA-C ( = −0.84 kcal/mol,
= −0.84 kcal/mol, 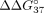 = −7.39 kcal/mol) (Table 4). Both LNA residues located at the 3′ end of SSO result in a much weaker thermodynamic effect (
= −7.39 kcal/mol) (Table 4). Both LNA residues located at the 3′ end of SSO result in a much weaker thermodynamic effect ( = −0.43 for sSSO3 and −0.65 kcal/mol for sSSO4).
= −0.43 for sSSO3 and −0.65 kcal/mol for sSSO4).
The application of UNA residues in our system results in a reversed effect from that observed for LNA residues. UNA modification
introduced at each mismatch position contributes to reduction of thermodynamic discrimination 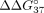 (Fig. 5; Table 4). Although full complementarity is present in the sSSO/sE10 duplex, the presence of UNA decreases sSSO preference for interaction
with sE10 over sI9. Therefore, the hybridization preference is shifted toward simultaneous interaction with both PKM fragments. In the case of UNA residues, modification of central adenosine has the strongest influence on
(Fig. 5; Table 4). Although full complementarity is present in the sSSO/sE10 duplex, the presence of UNA decreases sSSO preference for interaction
with sE10 over sI9. Therefore, the hybridization preference is shifted toward simultaneous interaction with both PKM fragments. In the case of UNA residues, modification of central adenosine has the strongest influence on 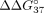 , which changes from −6.55 to −0.99 kcal/mol. This value shows that mismatch discrimination was strongly reduced, with a simultaneous
800-fold reduction of the hybridization constant for duplex formation for sSSO6/sE10 versus sSSO6/sI9. According to the literature,
UNA-A/A mismatch contributes to the most significant decrease in mismatch discrimination when taking into account every possible
adenosine mismatch (Pasternak and Wengel 2010). The reduction of mismatch discrimination was also observed for the sSSO7 oligonucleotide, for which
, which changes from −6.55 to −0.99 kcal/mol. This value shows that mismatch discrimination was strongly reduced, with a simultaneous
800-fold reduction of the hybridization constant for duplex formation for sSSO6/sE10 versus sSSO6/sI9. According to the literature,
UNA-A/A mismatch contributes to the most significant decrease in mismatch discrimination when taking into account every possible
adenosine mismatch (Pasternak and Wengel 2010). The reduction of mismatch discrimination was also observed for the sSSO7 oligonucleotide, for which  = 4.17 kcal/mol (Table 4). The weakest impact on mismatch discrimination reduction was noted for terminal nucleotide residues. Modified 5′ cytidine
increased
= 4.17 kcal/mol (Table 4). The weakest impact on mismatch discrimination reduction was noted for terminal nucleotide residues. Modified 5′ cytidine
increased 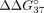 to the value of −5.41 kcal/mol (
to the value of −5.41 kcal/mol ( = 1.14 kcal/mol), and modified adenosine at the 3′ end decreased mismatch discrimination by 1.03 kcal/mol (
= 1.14 kcal/mol), and modified adenosine at the 3′ end decreased mismatch discrimination by 1.03 kcal/mol (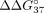 = −5.52 kcal/mol).
= −5.52 kcal/mol).
The influence of modified SSOs on PKM alternative splicing regulation
Thermodynamic studies clearly show that the application of UNA and LNA-modified residues within mismatches has a strong impact
on the thermodynamic parameters of duplexes. According to the presented data, we might expect that an LNA residue in mismatch
positions will enhance the preference of the oligonucleotide to interact with a complementary fragment (exon 10). On the other
hand, the UNA application will result in equalization of 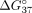 values of both duplexes. In consequence, the oligonucleotide should interact with similar probability with both pre-mRNA
fragments (intron 9 and exon 10). Our next aim was to verify the usefulness of mismatch discrimination in modulating PKM alternative splicing regulation. Therefore, we synthesized 15 nt long oligonucleotides with different numbers of modified
residues and transfected them to the HeLa cell line (Table 5). The qPCR method was used to evaluate the levels of PKM1 and PKM2 mRNA expressed in response to transfection of the modified
oligonucleotides.
values of both duplexes. In consequence, the oligonucleotide should interact with similar probability with both pre-mRNA
fragments (intron 9 and exon 10). Our next aim was to verify the usefulness of mismatch discrimination in modulating PKM alternative splicing regulation. Therefore, we synthesized 15 nt long oligonucleotides with different numbers of modified
residues and transfected them to the HeLa cell line (Table 5). The qPCR method was used to evaluate the levels of PKM1 and PKM2 mRNA expressed in response to transfection of the modified
oligonucleotides.
The list of oligonucleotides applied in cell culture studies. All oligonucleotides are 2'MOE-modified with phosphorothioate backbone.
According to the level of thermodynamic change, the most significant positions were chosen to be modified in full-length SSOs. In the oligonucleotides SSO9 and SSO10, the central adenosine residue was replaced by LNA-A and UNA-A, respectively (Table 5).
The in vitro experiments revealed that our assumptions are correct. We noticed an interesting tendency of increased PKM1 level and reduced PKM2 level when mismatch discrimination was enhanced in reference to an unmodified oligonucleotide (Fig. 6, SSO9). Additionally, the opposite tendency was observed when mismatch discrimination was reduced by UNA-modified residue (SSO10); PKM1 level was significantly reduced to 4.6, whereas for unmodified oligonucleotide, PKM1 expression was equal to 11.8. These results indicate that specific hybridization of oligonucleotide with exon 10 results in stronger regulatory properties of the studied oligonucleotide. On the contrary, the SSO10 interactions with both PKM fragments reduce the regulatory effect of the oligonucleotide. To support our hypothesis, we synthesized another set of modified oligonucleotides with additional modified positions (Table 5). As before, we followed the extent of thermodynamic change observed for the modified positions. Except for LNA-modified central adenosine, the LNA modification was also used in 5′ and 3′ terminal positions (SSO11). On the contrary, the UNA residues were used only in two positions. The magnitude of thermodynamic change caused by UNA modifications is stronger than for LNA residues, and a larger number of UNA residues might result in a lack of hybridization even with exon 10. Therefore, the UNA residues were used only in the central and penultimate positions (SSO12).
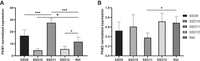
The normalized expression of PKM1 (A) and PKM2 (B) mRNA in response to transfection of SSOs with single or multiple UNA or LNA modifications. The expression of PKM1 and PKM2 mRNA was normalized against untreated cells mRNA. The error bars represent the standard deviation for six biological repeats. The statistical significance, marked with * and ***, was calculated with one-way ANOVA test. (*) P < 0.05; (***) P < 0.001.
The presence of three LNA residues results in a significant increase in oligonucleotide regulatory properties. We noticed a twice higher level of PKM1 mRNA for cells transfected with SSO11 in reference to an unmodified oligonucleotide (Fig. 6A). At the same time, the expression level of PKM2 mRNA was reduced from 0.7 to 0.4. These results prove that increased mismatch discrimination enhances the regulatory effect of SSOs. Surprisingly, the SSO12 oligonucleotide with two UNA residues showed a similar effect as SSO10 with one UNA residue (Fig. 6). It seems that additional modification did not further change the hybridization of the oligonucleotide with exon 10. Presumably, the preference of SSO12 for hybridization with exon 10 and intron 9 is similar to that of SSO10. According to the thermodynamic studies described above, one LNA residue increased the thermodynamic stability of sSSO/sE10 by about 1 kcal/mol and, at best, about half of this value for sSSO/sI9. Therefore, it can be deduced that LNA application in three positions increases SSO hybridization with exon 10 mainly, and not with intron 9. This, in turn, indicates that reduction of the SSO/I9 duplex contribution in the overall SSO-mediated duplex formation results in a stronger splicing regulatory effect. It provides evidence that exon 10 is the regulatory sequence responsible for exon 10 splicing. This conclusion is partially consistent with that of the Krainer group, who proposed that exon 10 is a main regulatory element; however, part of the regulatory effect is caused by hybridization with the intron 9 fragment (Wang et al. 2012). Our studies indicate that oligonucleotide interaction with intron 9 is not required to achieve a splicing switch, and off-target interactions with intron 9 decrease the regulatory potential of SSO. The introduction of UNA into SSO results in strong destabilization, mainly of the duplex with exon 10. At the same time, we increased the probability of SSO interactions with intron 9, resulting in significant sSSO/sE10 destabilization and only slight destabilization of sSSO/sI9. In this scenario, we predict that both potential regulatory fragments (intron 9, exon 10) are sterically blocked by SSO. Our studies show that it weakens the regulatory properties of SSO and that efforts must be made to maximize mismatch discrimination and hybridization with specific sequence in exon 10.
Our studies confirm that UNA and LNA modifications might be highly useful in oligonucleotide-mediated splicing regulation. The mechanism of action of SSOs is based on the hybridization with pre-mRNA regulatory motifs, which prevents regulatory proteins from binding. The example of PKM pre-mRNA shows that in one gene, we can find two similar sequences that are prone to interact with the same oligonucleotide. We used thermodynamic analyses as well as cell culture studies to determine more favorable hybridization with PKM pre-mRNA for SSO activity. In the PKM pre-mRNA, the hybridization with one specific RNA fragment provided more benefits than simultaneous interactions with two fragments. However, it cannot be excluded that altering mismatch discrimination to cover two or more regulatory motifs in different target genes would increase the therapeutic potential of other SSOs.
Furthermore, sequentially similar fragments might be present in the whole cellular transcriptome, and although they do not form fully complementary duplexes, their interactions with SSO might be stable enough to influence gene expression. The model oligonucleotide studied herein is prone to potentially hybridize with 22 different genes, except the intron 9 fragment. A significant change in splicing pattern was observed for 14 of them as a result of SSO transfection (Scharner et al. 2020). Unspecific interactions usually unfavorably influence the oligonucleotide therapeutic effect. Therefore, there is a high need to search for methods that will allow minimization of off-target hybridization. Our studies proved that LNA increases mismatch discrimination and simultaneously improves hybridization with target RNA. On the other hand, the UNA modifications reduce mismatch discrimination and allow for oligonucleotide hybridization even with sequences that are not fully complementary. It opens the possibility to design oligonucleotides for other approaches where simultaneous interactions with two different molecular targets are desirable.
MATERIALS AND METHODS
Synthesis of oligonucleotides
All oligonucleotides were synthesized on MerMade12 using β-cyanoethyl phosphoramidite chemistry and commercially available nucleoside phosphoramidites. RNA oligonucleotides were treated with a 30% ammonia/ethanol (3:1) solution and incubated at 55°C for 18 h. The oligonucleotide solutions were then separated from the solid support and evaporated. Next, oligonucleotides were incubated with triethylamine trihydrofluoride, in the presence of dimethylformamide as a solvent, at 55°C for 2–3 h. Oligonucleotides were precipitated in butanol, followed by Sephadex column desalting. Oligonucleotides were purified via 12% polyacrylamide gel electrophoresis in denaturing conditions. 2′MOE oligonucleotides were deprotected by 16 h incubation with 1.5 mL of 30% ammonia at 55°C. After solid support removal and ammonia evaporation, the oligonucleotides were purified via 12% polyacrylamide gel electrophoresis. All oligonucleotides were desalted before starting experiments. The composition of all oligonucleotides was confirmed by MALDI-TOF mass spectrometry. Concentrations of oligonucleotide stock solutions were determined by UV measurements at λ = 260 nm, 83°C.
Thermodynamic studies of modified and unmodified duplexes
Thermodynamic measurements were carried out on a JASCO V 650 spectrophotometer with a thermoprogrammer. Nine solutions of SSO oligonucleotides with RNA were prepared in a molar ratio of 1:1 and concentration range 10−3–10−6 M. Solutions were prepared in melting buffer (50 mM NaCl, 20 mM sodium cacodylate, 0.5 mM Na2EDTA, pH 7.0). Quartz microcuvettes with optical paths of 1 mm (30 μL), 5 mm (150 μL), and 10 mm (300 μL) were used. Melting was performed at 260 nm in the temperature range of 4°C–90°C, with a 1°C/min ramp rate. The absorbance values of the samples were collected every 0.5 min. The resulting melting curves were analyzed in MeltWin 3.5 software.
Cell culture and oligonucleotides transfection
HeLa cell line was cultured in RPMI 1640 medium supplemented with vitamins, antibiotics, and 10% FBS. Cells were seeded in 24-well plates at a density of 1 × 105 cells/well, which gave 95% confluence the next day. The cells were incubated at 37°C with 5% CO2 and 95% humidity. After 24 h, the medium was exchanged for antibiotic-free medium. Oligonucleotides were dissolved in OPTI-MEM medium with Lipofectamine 3000 as a transfection reagent (1 μL of Lipofectamine and 1 µL of P3000 reagent per one reaction) and added to cells in a final concentration of 125 nM. Cells were harvested 48 h after transfection. All transfections were performed at least in biological triplicate.
RT-qPCR analysis
The RNA from the cultured cells was isolated using acid guanidinium thiocyanate-phenol-chloroform extraction, and the RNA was treated with DNase I.
After DNAse treatment, a 200 ng RNA template was used for cDNA synthesis using the LunaScript RT SuperMix Kit. qPCR was performed on a CFX96 real-time PCR system (Bio-Rad) using Luna Universal qPCR Master Mix and 96-well clear plates. Two pairs of target gene primers were designed to quantify the amount of PKM1 and PKM2 isoforms. Primers for PKM2 isoform amplification: 5′ ATTGCCCGTGAGGCAGAGG 3′ and 5′ TGCCAGACTTGGTGAGGACGATTA 3′. Primers for PKM1 isoform amplification: 5′ GTTCCACCGCAAGCTGTTTGAAGA 3′ and 5′ TGCCAGACTCCGTCAGAACTATCA 3′. The expression of isoforms was normalized against the β-actin gene (reference gene primers: 5′ GCCAGCAGCCTCTGATCTG 3′ and 5′ CTGGTTCTTGCCAGCCTCTAG 3′). The Ct values of the human β-actin gene were in the range of 17–19. The qPCR cycles were as follows: 95°C, 1 min for predenaturation step: (95°C, 15 sec and 60°C, 30 sec) for 34 cycles. Analysis of qPCR data was performed in Bio-Rad CFX Maestro software.
Statistical analysis
The statistical analysis for qPCR results was performed with GraphPad 8.0.1 software. An ordinary one-way ANOVA test was applied followed by Tukey's multiple comparisons test. A P-value < 0.05 was considered significant.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080212.124.
- Received August 2, 2024.
- Accepted December 13, 2024.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Natalia Bartyś is the first author of this paper, “Thermodynamic control of mismatch discrimination for extensive splicing regulation of PKM pre-mRNA.” Natalia is an assistant professor at the Institute of Bioorganic Chemistry, Polish Academy of Sciences. Her scientific interests are focused on the application of various nucleic acid–based tools for cancer therapy, with a particular emphasis on the regulation of alternative splicing.
What are the major results described in your paper and how do they impact this branch of the field?
In the article, we presented an innovative approach to enhance the regulatory properties of splice-switching oligonucleotides designed to correct aberrant alternative splicing patterns of PKM pre-mRNA in cancer cells. For this purpose, we utilized modified nucleotides that exert opposite thermodynamic effects on duplex stability. These properties allowed us to eliminate off-target interactions with intron 9 and to improve the regulatory properties of the studied oligonucleotide. We believe that this approach could be valuable not only for minimizing off-target interactions but also for situations where targeting multiple RNA sequences is necessary.
What led you to study RNA or this aspect of RNA science?
My PhD project was focused on the optimization of different oligonucleotide tools that might be used to regulate mutually exclusive alternative splicing of PKM pre-mRNA. Although I mainly focused on bifunctional oligonucleotides optimization, I also aimed to use the knowledge and techniques related with thermodynamic characterization of nucleic acids to boost the regulatory effect of SSO designed by Prof. Krainer's group. I believe that every research group has the potential to contribute another vital piece to the puzzle, further refining a particular drug or molecule. The collaboration across various disciplines and institutions strengthens the overall pursuit of scientific advancements, and together we move closer to breakthroughs that can significantly improve human health.
If you were able to give one piece of advice to your younger self, what would that be?
This answer will be quite simple—do not be afraid to ask questions or ask for help if you do not understand. The only person who can lose if you do not do this is you. Actually, I feel like I still need to remind myself of this advice.
Are there specific individuals or groups who have influenced your philosophy or approach to science?
I consider myself incredibly fortunate to have begun my scientific journey in the Department of Nucleic Acids Bioengineering, under the leadership of Prof. Anna Pasternak. My supervisor and all of my colleagues uphold the highest scientific standards and share a deep commitment to advancing research with the goal of developing new therapies. We work collaboratively every day, exchanging ideas, knowledge, and experiences to drive our studies forward. I believe our shared, strong motivation to contribute therapeutic solutions stems from a profound sense of empathy and significant personal experiences, which fuel our dedication to making a meaningful impact in the field of medical science.
What are your subsequent near- or long-term career plans?
In the near future, I aim to focus my scientific efforts on the further development of oligonucleotide drugs. I plan to initiate my second independent project, which will focus on the regulation of alternative splicing, with a particular emphasis on the role of RNA secondary structure.








