
Co-phase separation of Y14 and RNA in vitro and its implication for DNA repair
- 1Institute of Biomedical Sciences, Academia Sinica, Taipei 115, Taiwan
- 2Institute of Molecular Medicine, College of Medicine, National Taiwan University, Taipei 100, Taiwan
- 3Biomedical Translation Research Center, Academia Sinica, Taipei 115, Taiwan
- Corresponding author: wtarn{at}ibms.sinica.edu.tw
Abstract
The multifunctional RNA recognition motif-containing protein Y14/RBM8A participates in mRNA metabolism and is essential for the efficient repair of DNA double-strand breaks (DSBs). Y14 contains highly charged, low-complexity sequences in both the amino- and carboxy-terminal domains. The feature of charge segregation suggests that Y14 may undergo liquid–liquid phase separation (LLPS). Recombinant Y14 formed phase-separated droplets, which were sensitive to pH and salt concentration. Domain mapping suggested that LLPS of Y14 involves multivalent electrostatic interactions and is partly determined by the net charge of its low-complexity regions. Phospho-mimicry of the carboxy-terminal arginine-serine dipeptides of Y14 suppressed phase separation. Moreover, RNA could phase separate into Y14 droplets and modulate Y14 LLPS in a concentration-dependent manner. Finally, the capacity of Y14 in LLPS and coacervation with RNA in vitro correlated with its activity in DSB repair. These results reveal a molecular rule for LLPS of Y14 in vitro and an implication for its co-condensation with RNA in genome stability.
Keywords
INTRODUCTION
Mammalian cells contain more than 1000 distinct RNA binding proteins (RBPs) that participate in RNA metabolism, and many RBPs have additional roles in DNA damage repair, immune response, and cellular signaling (Gerstberger et al. 2014). RBPs have modular structures (Lunde et al. 2007), and most have one or more RNA recognition motifs (RRM) or K homology domains for RNA binding and additionally contain regions with low-complexity sequences. These regions tend to be structurally disordered and have the capacity to mediate liquid–liquid phase separation (LLPS) of RBPs in vitro under certain conditions (Wiedner and Giudice 2021). RBPs containing intrinsically disordered regions (IDRs) are compartmentalized in membrane-less ribonucleoprotein granules in cells (Fay and Anderson 2018; Roden and Gladfelter 2021). RNA may serve as a scaffold or modulator for RBP phase transition. For example, stress granules assemble through multivalent RBP–RNA interactions. The IDRs of the stress-granule nucleator G3BP as well as its associated RNAs facilitate and regulate the formation of condensates (Guillen-Boixet et al. 2020; Yang et al. 2020). Similarly, other ribonucleoprotein granules such as the nucleolus and paraspeckles also assemble through LLPS. Moreover, LLPS is widely used as a mechanism for RNA-mediated cellular function. For example, the long noncoding (lnc) RNA NORAD promotes condensation of the RBP Pumilio to facilitate its function in genome maintenance (Elguindy and Mendell 2021). Finally, various cellular signaling pathways affect the formation of biological assemblies via post-translational modification of proteins or by regulating RNA abundance (Itakura et al. 2018).
LLPS-mediated biomolecular condensates ensure the specificity and efficiency of various cellular processes. However, dysregulation of LLPS may trigger protein aggregation in many neurodegenerative diseases (Zbinden et al. 2020). Pathological mutations in the IDRs of several RBPs may drive liquid-to-solid phase transition. For example, amyotrophic lateral sclerosis-associated mutations in fused in sarcoma (Fus) induce aberrant phase transitions within liquid-like compartments, resulting in its amyloid-like aggregation (Patel et al. 2015; Murthy et al. 2019). TAR DNA-binding protein-43 has a glutamine/asparagine-rich prion-like domain in the carboxyl terminus; mutations in this domain cause cytoplasmic mislocalization and aggregation, leading to proteinopathies in different types of neurodegenerative diseases including amyotrophic lateral sclerosis, Alzheimer's disease and Parkinson's disease (Gao et al. 2018; Zbinden et al. 2020).
Y14 (RBM8A) primarily acts as a core factor of the exon junction complex (EJC) that binds newly spliced mature mRNAs. The EJC functions in mRNA surveillance, by which premature stop codon-containing mRNAs are identified and subsequently degraded (Boehm and Gehring 2016). Y14 forms a stable heterodimer with Magoh, which subsequently associates with eIF4A3 in the EJC core (Boehm and Gehring 2016). Moreover, Y14 also contributes to the regulation of precursor mRNA splicing and degradation, cellular signaling, and DNA damage repair (Chuang et al. 2013; Togi et al. 2013; Fukumura et al. 2016; Lu et al. 2017; Chuang et al. 2019). Y14 colocalizes with splicing factors in nuclear speckles and interacts with the mRNA export factor NXF1 around speckles, where the EJC is assembled (Schmidt et al. 2006; Daguenet et al. 2012). The phosphorylation status of the carboxy-terminal arginine-serine (RS) dipeptides of Y14 affects its subnuclear localization (Tatsuno and Ishigaki 2018). It has been reported that a portion of Y14 does not associate with Magoh (Choudhury et al. 2016; Tatsuno and Ishigaki 2018). We have also shown that as much as 50% Y14 but not Magoh is present in the chromatin-enriched fraction, indicating that a substantial fraction of nuclear Y14 is possibly Magoh-free (Chuang et al. 2019). Y14 plays a role in DNA double-strand break (DSB) repair via its interaction with the DNA repair factors and localized to DNA damage sites (Chuang et al. 2019). Whether Y14 exerts its cellular function via LLPS has not been tested.
The central region of Drosophila Y14 contains an RRM that contributes to heterodimerization with Mago (Fribourg et al. 2003; Shi and Xu 2003). The amino-terminal region contains an α-helix followed by a β-strand, which contacts Mago in the heterodimer (Fribourg et al. 2003). The unstructured carboxy-terminal region of Y14 was eliminated in structural studies of the Y14/Mago heterodimer (Fribourg et al. 2003; Shi and Xu 2003). We noted that both the amino- and carboxy-terminal low-complexity regions of Y14 are rich in charged amino acid residues that may contribute to LLPS. Therefore, we characterized LLPS of Y14 in vitro and investigated whether it contributes to Y14 function.
RESULTS
Y14 contains evolutionarily conserved low-complexity domains rich in charged residues
All Y14 homologs have the same modular structure, comprising a central RRM flanked by low-complexity terminal domains (Fig. 1A). Both the amino- and carboxy-terminal domains are essentially unstructured except that the former contains a short α-helix and β-sheet (Fig. 1A; Fribourg et al. 2003; Shi and Xu 2003). Human Y14 is 100% identical to its mouse homolog, whereas it is only ∼64% identical to Drosophila Y14/Tsunagi and differs substantially from Caenorhabditis elegans Ce-Y14 (Fig. 1B). However, analysis of charge distribution in the terminal domains of human, Drosophila and Caenorhabditis Y14 homologs (Fig. 1B) using Classification of Intrinsically Disordered Ensemble Regions (http://157.245.85.131:8000/CIDER/) (Holehouse et al. 2017) revealed a conserved distribution of charged residues. The amino-terminal domain contains alternating stretches that are rich in either positively or negatively charged residues (Fig. 1C). This pattern of charge segregation is reminiscent of a set of proteins that have a high propensity to undergo LLPS, such as nucleolar nucleoplasmin 1 (NPM1), the stress-granule nucleator G3BP1, and the microtubule-associated protein tau (Mitrea et al. 2016, 2018; Boyko et al. 2019; Yang et al. 2020). Moreover, the carboxy-terminal region (ranging from 9 to 23 residues) of three Y14 homologs is highly positively charged. Therefore, these uniformly charged segments possibly contribute to electrostatic interactions intramolecularly and/or intermolecularly. Finally, the disorder-promoting score (Das and Pappu 2013) for each of the amino- and carboxy-terminal domains of human Y14 was calculated as 0.8 and 1.0, respectively (Supplemental Table S1), implying that they are relatively unstructured. The IDRs of Y14 do not have glutamine/asparagine-rich sequences that are typical of several disease-linked RBPs (Zbinden et al. 2020). Nevertheless, it was important to decipher whether Y14 undergoes LLPS and, if so, whether LLPS occurs in a manner similar to or distinct from that of disease-linked RBPs.
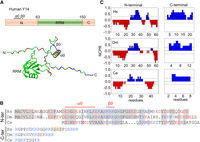
Charge distribution of the terminal domains of Y14. (A, upper panel) Domain organization of human Y14. (Lower panel) The structure of human Y14 was predicted by the AlphaFold Protein Structure network (https://github.com/deepmind/alphafold); the image was manipulated in PyMol Software (pymol.org). In all panels, acidic and basic residues are indicated in red and blue, respectively. RRM, RNA recognition motif; α0 and β0 represent the short α-helix and β-sheet, respectively. (B) Amino acid sequence alignment of human (Hs), Drosophila (Dm), and Caenorhabditis (Ce) Y14 homologs is shown according to a previous report (Fribourg et al. 2003). Conserved residues (identical or similar) in the amino-terminal domain are highlighted in gray. The two RS dipeptides in the carboxy-terminal domain of human Y14 are highlighted in yellow. (C) The amino- and carboxy-terminal domains of the three Y14 homologs in B were analyzed via Classification of Intrinsically Disordered Ensemble Regions (http://pappulab.wustl.edu/CIDER/analysis/). (NCPR) Net charge per residue distribution.
Y14 undergoes LLPS in a pH-dependent manner
To investigate whether Y14 forms liquid-like droplets in vitro, we generated recombinant Y14 containing an amino-terminal S peptide (to promote expression in Escherichia coli) and a carboxy-terminal polyhistidine (His) tag (to enable affinity purification) (Fig. 2A; Supplemental Fig. 1A; Hsu et al. 2005). Analytic ultracentrifugation of purified recombinant Y14 at pH 7.9 revealed a single peak at the estimated molecular mass, reflecting its monomeric form (Supplemental Fig. 1B). This dual-tagged Y14 had an estimated isoelectric point (pI) value of 5.86 (Supplemental Table S2). We initially evaluated droplet formation of Y14 (20 µM) in buffers containing 50 mM KCl at pH values ranging from 6 to 7.5 using a differential interference contrast (DIC) microscope. Spherical droplets of Y14 formed after a 2-h incubation in pH 6.5 buffer (Fig. 2B). The number and size of Y14 droplets decreased gradually with increasing pH. On the other hand, Y14 droplets tended to form large amorphous assemblies at pH 6.0 (Fig. 2B). RNase treatment of recombinant Y14 did not affect droplet formation (Supplemental Fig. 1C). Next, we performed light scattering measurements at 400 nm to monitor the dynamics of droplet formation. As the optical density was recorded at 5-min intervals, we observed that Y14 underwent LLPS immediately when the pH changed from 7.9 to the range of 6.0 to 7.0, whereas no droplets formed at pH 7.5 (Fig. 2C, upper), consistent with the DIC observations. Y14 droplets precipitated over time with a rate that varied with pH. Faster precipitation at pH 6.0 coincided with larger droplets or aggregates of Y14 (Fig. 2C, upper). Evaluation during the first 25 min at 10-sec intervals revealed that the turbidity of 20 µM Y14 at pH 6.5 peaked after ∼5 min of incubation (Fig. 2C, bottom), indicating that pH alteration immediately triggered Y14 droplet formation. Y14 droplets that formed at pH 6.5 dissolved when the pH was adjusted to 8 and reformed after the pH returned to 6.5, and this cycle could be repeated (Fig. 2D). Reversible formation of Y14 droplets upon a pH change from 6.5 to 8 suggested a pH-dependent Y14 phase separation. Therefore, mild acidic conditions might promote Y14 droplet formation by reducing electrostatic repulsion. The observation that the phase behavior of Y14 could cycle with pH supported the idea that electrostatic interactions contribute to LLPS of Y14.
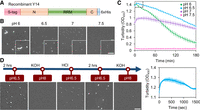
Y14 forms phase-separated droplets in a pH-dependent manner. (A) Recombinant Y14 containing the S peptide (S-tag) and hexahistidine (6 × His) tag. All recombinant Y14 variants contained these two tags. (B) In vitro LLPS of 20 µM recombinant Y14 was performed in buffers containing 50 mM KCl at different pH values for 2 h. DIC images show Y14 droplets (400 × 400 pixels; 1 pixel = 0.114942 µm); inset shows magnified images (100 × 100 pixels) of a selected area. Bar, 5 µm. The experiment was repeated more than five times (N > 5). (C) Turbidity assay showing the kinetics of LLPS of Y14 (20 µM) as in B. Turbidity at 400 nm (OD400) was measured. Optical density was recorded at 5-min intervals for 3 h (upper). Dashed square: recording was additionally taken at 10-sec intervals for the first 25 min (lower). Mean and standard deviation were calculated for two to four replicates. (D) Y14 (20 µM) was initially incubated in a pH 6.5 buffer. The pH of the buffer was subsequently adjusted by sequential addition of KOH, HCl, and KOH again, as indicated. DIC images were acquired immediately or after a 2-h incubation, as indicated. Bar, 5 µm. N = 2.
Y14 droplets are dynamic and sensitive to salt concentration
Phase-separated droplets have several characteristics including fusion capacity, reversibility and dynamic exchange of components (Guo et al. 2021). The above result demonstrated that Y14 droplet formation could be reversed in response to a change in pH (Fig. 2D). Using DIC microscopy, we observed that two Y14 droplets could rapidly coalesce in 50 mM KCl/pH 6.5 buffer (Fig. 3A). Although Y14 underwent phase separation independent of molecular crowders, the presence of PEG4000 slightly increased the size of Y14 droplets (Supplemental Fig. 2A). Next, we examined the effect of KCl concentration on Y14 LLPS. The size of Y14 droplets increased with increasing KCl concentration up to 100 mM, although higher KCl concentrations inhibited droplet formation (Fig. 3B). Therefore, except for electrostatic interactions, Y14 LLPS may also involve other types of interaction forces. The notion that the aliphatic alcohol 1,6-hexanediol substantially disrupted Y14 droplets (Fig. 3C) implied the involvement of hydrophobic interactions. Conceivably, droplet size was proportional to Y14 concentration ranging from 5 to 50 µM in pH 6.5 buffer containing 50 mM KCl (Supplemental Fig. 2B). To measure the size of Y14 droplets, we conjugated purified Y14 with Alexa 488. Fluorescent Y14 signals overlapped well with the droplets, as observed by DIC and fluorescence microscopy (Supplemental Fig. 2C). Both coverage rate and droplet size gradually increased over time and peaked at ∼3 h and decreased thereafter (Fig. 3D for images and graph, and Supplemental Fig. 2D). For 20 µM Y14 at 3 h, the average droplet diameter was ∼0.9 µm. Together, the above results confirmed the liquid-like property of Y14 droplets, which were sensitive to pH and KCl concentration.
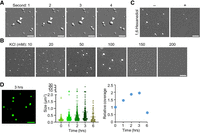
Characterization of phase-separated droplets of Y14. (A) The LLPS assay was performed using 50 µM Y14 in a pH 6.5 buffer containing 50 mM KCl. DIC images show the time course of a fusion event of Y14 droplets (arrowhead). Bars, 5 µm. (B) Y14 (20 µM) was incubated in a pH 6.5 buffer containing different concentrations of KCl as indicated. Bar, 5 µm. N = 3. (C) Y14 (20 µM) was incubated in a pH 6.5 buffer containing 50 mM KCl in the absence or presence of 1,6-hexanediol. Bar, 5 µm. (D) A mixture of Alexa Fluor 488-conjugated Y14 and unlabeled Y14 at the molar ratio of 1:50 was incubated in a pH 6.5 buffer containing 50 mM KCl for different times. A representative fluorescent image (200 × 200 pixels) shows the droplets after 3-h of incubation. Bar, 5 µm. The coverage rate (the coverage area of fluorescent signals divided by the total area) was measured by the software MetaMorph. Mean size and relative coverage rate (time 0 was set to 1) of the droplets are shown in the dot graphs.
Y14 droplet formation is determined by multivalent electrostatic interactions
Next, we explored how the highly charged IDRs of Y14 contribute to its LLPS. The amino-terminal domain can be roughly divided into three subdomains, namely N1, N2, and N3, that are rich in negatively, positively and negatively charged residues, respectively (Fig. 4A). We generated and purified recombinant Y14 truncations, each of which lacked one or two subdomains (Fig. 4B). Removal of either positively charged domain (N2 or C) disrupted Y14 droplet formation at pH 6.5–7.5 and caused Y14 aggregation at pH 6, indicating that both domains are essential for LLPS of Y14 and cannot compensate each other (Fig. 4C, ΔN2 and ΔC). In contrast, deletion of either negatively charged N1 or N3 resulted in larger droplets at pH 6.5 (Fig. 4C, ΔN1 and ΔN3). Because these two truncations increased the pI value compared with wild-type Y14 (Supplemental Table S2), their droplets were detectable at higher pH values (7 and 7.5). We therefore concluded that the presence of oppositely charged regions was necessary for electrostatic interactions whereas one negatively charged region appeared to be sufficient for LLPS. Surprisingly, removal of both N1 and a large portion of N3, leaving very few negatively charged residues, did not disrupt droplet formation (Fig. 4C, ΔN1N3), and instead increased the droplet size at pH 7.5, in contrast to the full-length and the truncations lacking either N1 or N3 that developed larger droplets at pH 6.5 than higher pH (Fig. 4D). LLPS of ΔN1ΔN3 may involve other types of interaction forces (see Discussion).

Charged regions of Y14 differentially influence LLPS. (A) The domain and subdomain organization of human Y14. Negatively and positively charged regions are indicated in red and blue, respectively. (B) SDS-PAGE gel showing purified recombinant full-length and truncated Y14 proteins. Protein bands were stained with ToolStart Blue Staining Reagent. ΔN2 showed a molecular weight larger than expected perhaps due to its high negative charge density in the amino terminus. (C) Diagrams showing domains of full-length and truncated Y14 proteins. Droplet formation for all Y14 versions (20 µM each) was performed at different pH values. Representative DIC images are shown. Bar, 5 µm. N > 3. (D) The droplet size of Y14 proteins as indicated was measured from the images of C. Mean and standard deviation were calculated for 50 droplets each. (E) DIC images showing droplet formation of wild-type Y14 or Y14-SD in buffer containing 50 mM KCl at the pH values indicated. Representative images (200 × 200 pixels) are shown. Bar, 5 µm. N = 3. Bar graph: the turbidity was measured as in E (mean ± SD; N = 2–4; [**] P < 0.01, [****] P < 0.0001).
Phosphorylation suppresses Y14 LLPS
Y14 has two RS dipeptide repeats in the carboxy-terminal domain. Phosphorylation of Y14 at the RS repeats modulates its protein–protein interactions as well as its subcellular localization (Hsu et al. 2005; Chuang et al. 2013; Tatsuno and Ishigaki 2018). We generated a phosphomimetic mutant of Y14 containing two serine-to-aspartate (SD) substitutions at the RS dipeptides. Y14-SD formed droplets at pH 6.5, albeit slightly fewer than that of wild-type Y14, but droplets did not form at pH 7.0 (Fig. 4E, DIC). This observation was confirmed by the turbidity assay (Fig. 4E, bar graph). This result may reflect the effect of the lower pI value of Y14-SD compared with wild-type or, alternatively, disruption of the charge blockiness of the carboxy-terminal IDR (see Discussion). Nevertheless, this result suggested that phosphorylation of the RS dipeptides may inhibit LLPS of Y14.
Positively charged peptides in trans promote LLPS of truncated Y14
Deletion of the Arg-rich carboxy-terminal domain disrupted Y14 LLPS (Fig. 4C). Therefore, we wondered whether the synthetic carboxy-terminal peptide could in trans promote droplet formation of ΔC. The carboxy-terminal peptide of human Y14 (HsC23), although failed to form droplets by itself, induced LLPS of Y14ΔC when mixed at an equimolar concentration at pH 6.5 (Fig. 5A,B, HsC23). This result was reminiscent of a previous finding that the oppositely charged domains of tau—although individually having low propensity for LLPS—could form droplets after mixing (Boyko et al. 2019). Surprisingly, HsC23 also promoted LLPS of ΔN2, despite N2 containing more lysine residues than Arg residues (Fig. 5A). This observation indicated that positively charged residues in the N2 or C domain are important for LLPS. We noted that, in contrast to human Y14, Drosophila and Caenorhabditis Y14 had a shorter and Lys-rich carboxyl terminus (Fig. 5A). DmC14 could promote droplet formation of both ΔC and ΔN2, although the size of DmC14-induced droplets was smaller than those induced by HsC23 (Fig. 5C). CeC9 barely induced LLPS perhaps because it was too short. The amino-terminal IDR of the human papillomavirus E7 protein, lacking positively charged residues, had no effect on in trans LLPS induction (Fig. 5B, E7N). Our result emphasized the critical role of the positively charged N2 and C regions in LLPS; however, Arg and Lys residues appeared to be interchangeable, at least to some extent.
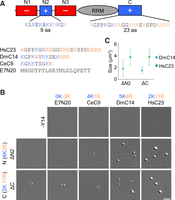
Carboxy-terminal peptides of Y14 in trans promote LLPS of truncated Y14. (A) Schematic diagram shows the sequence of N2 and C regions of human Y14 (upper), and the peptides derived from the carboxy-terminal domain of three Y14 homologs and the amino-terminal IDR of HPV E7 protein (E7N20). (B) The LLPS assay was performed with indicated peptide alone (20 µM) or in a mixture containing ΔN2 or ΔC (20 µM) in a 50 mM KCl pH 6.5 buffer. Representative DIC images are shown. Bar, 5 µm. N = 2. (C) The droplet size of truncated Y14/peptide mixtures was measured. Mean and standard deviation were calculated for 50 droplets each.
RNA modulates Y14 droplet formation
RNA acts as a scaffold for several membrane-less organelles and regulates the size of ribonucleoprotein assemblies in different cellular processes such as transcription and stress-granule formation (Fay and Anderson 2018; Maharana et al. 2018; Garcia-Jove Navarro et al. 2019; Henninger et al. 2021). Therefore, we evaluated whether RNA could co-condense with Y14 and thereby modulate Y14 droplet formation. Both DIC and fluorescence microscopy revealed that Cy5-labeled poly(U) 25-mer (U25) RNA colocalized with Y14 droplets (Fig. 6A, wild-type), whereas U25 RNA alone did not form droplets. We previously identified a single point mutation (W73V) of Y14 that substantially decreases its RNA binding capacity (Chuang et al. 2016). In the absence of RNA, Y14–W73V formed a greater number of slightly smaller droplets as compared with wild-type Y14 (Fig. 6A, W73V, no RNA). When Cy5–U25 was added to W73V, fluorescently labeled droplets were barely detected (Fig. 6A, +RNA). A similar result was observed using fluorescent poly(A) 25-mer (A25) RNA (Supplemental Fig. 3A). Therefore, the RNA binding activity of Y14 is critical for its recruitment of RNA to phase-separated condensates. Next, we titrated U25 in the phase separation assay. Consistent with previous findings (Maharana et al. 2018; Garcia-Jove Navarro et al. 2019; Henninger et al. 2021), low levels of U25 promoted phase separation of Y14, whereas high levels of U25 had an inhibitory effect, indicating that RNA influences Y14 LLPS in a concentration-dependent manner (Fig. 6B; Supplemental Fig. 3B). Yeast total RNA, however, inhibited Y14 droplet formation even at very low concentrations (Supplemental Fig. 3C). Considering that cellular RNAs are, on average, >100-fold longer than synthetic polymers, the excess negative charge of RNA may interfere with Y14 LLPS, consistent with previous observations (Maharana et al. 2018).
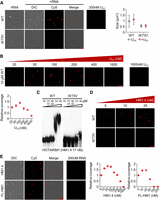
RNA modulates LLPS of Y14. (A) Wild-type or W73V Y14 (20 µM) was mock-incubated (no RNA, DIC) or incubated with 200 nM Cy5-labeled U25 (DIC, Cy5, and merge) in a pH 6.5 buffer containing 50 mM KCl for 2 h. Cy5-labeled U25 (200 nM) alone is shown at right. Bar, 5 µm. Dot graph shows the size of droplets in the absence or presence of RNA (50 droplets were used for measurement). (B) Y14 (10 µM) was incubated with increasing concentrations of U25 RNA (a mixture of Cy5-labeled and unlabeled U25 RNA). Bar, 5 µm. Cy5-labeled U25 (1600 nM) alone is shown at right. Dot graph shows the relative coverage rate of Y14 in the presence of different concentrations of U25. Coverage rate at 25 nM U25 was set to 1. N = 3. (C) EMSA showing Y14 (wild-type or W73V, concentration as indicated) binding to 32P-labeled HM1.4 (HOTAIRM1 632-882, 17 nM). (D) Wild-type or W73V Y14 (10 µM) was mixed with Cy5-labeled HM1.4 (concentration as indicated) for the LLPS assay. Bar, 5 µm. (E) Wild-type Y14 (10 µM) was mixed with Cy5-labeled HM1.4 (5 to 1600 nM) or full-length HOTAIRM1 (FL-HM1, 5 to 200 nM) for the LLPS assay. Representative images (DIC, fluorescence, merged) show Y14 LLPS in the presence of 25 nM RNA. Cy5-labeled RNAs (200 nM) alone are shown at right. Bar, 5 µm. Dot graphs show the relative coverage rate that was measured over the entire range of concentrations of HM1.4 and full-length HOTAIRM1 (FL-HM1). N = 3.
Y14 interacts with DNA repair factors via the lncRNA HOTAIRM1, of 1044 nt in length (Chuang et al. 2023). Therefore, we tested whether HOTAIRM1 could promote Y14 LLPS under certain circumstances. To determine how Y14 binds HOTAIRM1, we generated five HOTAIRM1 fragments, of which the size was suitable for electrophoretic mobility shift assay (EMSA). The result revealed that Y14 could bind all, albeit with a slightly higher affinity to the fragment of nucleotide 632–882 (namely HM1.4, Supplemental Fig. 3D). Y14–W73V did not bind HM1.4 (Fig. 6C). Cy5-labeled HM1.4 alone did not form droplets, whereas it co-phase separated with Y14 and promoted Y14 LLPS at lower concentrations (Fig. 6D; Supplemental Fig. 3E). Full-length HOTAIRM1 behaved similarly to HM1.4 (Fig. 6E, images). However, at higher concentrations, full-length HOTAIRM1 had a stronger effect than HM1.4 on suppressing Y14 LLPS (Fig. 6E, dot graphs). These results indicated that HOTAIRM1 not only co-condenses with Y14 but also has the capacity to modulate Y14 LLPS—possibly during the DNA repair process.
A potential role for Y14 LLPS in repairing DNA damage
Our in vitro analysis revealed that the carboxy-terminal truncation and W73V mutation abrogated the LLPS capacity of Y14 to different extents. Next, we evaluated the effects of these two mutants on the cellular distribution and function of Y14. Using a fusion protein comprising enhanced green fluorescent protein (GFP) and wild-type or mutant Y14, we examined their intracellular localization and mobility. Confocal microscopy revealed that wild-type Y14 and two mutants (W73V and ΔN2) formed speckles in the nucleoplasm, which were excluded from the nucleoli. In contrast, Y14ΔC showed a less prominent speckled pattern, and in part distributed into the nucleoli (Fig. 7A). Fluorescence recovery after photobleaching (FRAP) analysis revealed that wild-type and mutant Y14 foci were dynamic to different extents (Fig. 7B). All three mutants showed higher mobility than the wild-type. In particular, the fluorescence intensity of ΔC fluctuated, suggesting that it was least constrained.
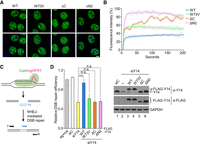
Co-phase separation of Y14 and RNA is essential for DNA damage repair. (A) U2OS cells were transfected with an expression vector encoding GFP-tagged Y14 (wild-type, W73V, ΔC, and ΔN2). Confocal microscopy images showing their subnuclear localization. Bar, 10 µm. (B) Graph showing the recovery kinetics of FRAP analysis of GFP-Y14 (as in A). Percentage represents normalized fluorescence intensity (to that of the first post-bleach value). Mean and standard-deviation values were calculated for 18–24 samples. (C) Diagram showing the design of NHEJ-mediated repair of DSBs. DSB was induced by transfection of cells with the Cas9/single guide RNA (sgHPRT) vector. Incorporation of the double-stranded oligonucleotide “Ins” into the DSB sites was measured by PCR using the indicated primers. (D) HeLa cells were transfected with Cas9/sgHPRT (all samples) without or with the indicated siRNA and siRNA-resistant FLAG-Y14 (wild-type or mutant). Genomic DNA was recovered for qPCR using the primer sets I/R and F/R; the latter was used for normalization. Bar graph shows relative PCR products (sg + Ins was set to 1), representing DSB repair efficiency (mean ± SD; N = 2–4; (***) P < 0.001; n.s., no significant difference). Immunoblotting was performed using anti-Y14, which detected endogenous Y14 and overexpressed FLAG-Y14, and anti-FLAG, which detected FLAG-Y14 (endogenous and FLAG-Y14ΔC at the same position on SDS-PAGE). GAPDH was used as the control.
Next, we evaluated whether the LLPS capacity of Y14 contributes to its activity in nonhomologous end joining (NHEJ)-mediated DSB repair. We took advantage of a CRISPR/Cas9-induced DNA cleavage system to measure DSB repair (Fig. 7C; Du et al. 2018). The Cas9 vector coexpressing a single guide RNA targeting HPRT, which encodes hypoxanthine guanine phospho-ribosyltransferase (Cas9/sgHPRT) was transfected into HeLa cells to generate a site-specific DSB in HPRT. Cotransfected double-stranded oligonucleotide (Ins) can be incorporated into HPRT via the NHEJ pathway (Du et al. 2018). We measured Ins incorporation by PCR using the primers complementary to the Ins sequence and HPRT. The level of PCR products represented DNA repair. The DNA repair efficiency was reduced by siRNA-mediated Y14 depletion by ∼50% (Fig. 7D, lane 3), and was restored by cotransfection of cells with siRNA-resistant wild-type Y14 (lane 4) but not by any Y14 mutant (lanes 5–7). This result supported that the LLPS potential of Y14 may contribute to its activity in DSB repair. Nevertheless, the observation that all three mutants, unlike the wild-type, failed to interact with the NHEJ factors and further accumulate on the chromatin after IR (Supplemental Fig. 4) provided a second explanation for their inability to repair DNA. The evaluation of Y14/RNA LLPS at DNA damage foci requires further investigation.
In conclusion, our results demonstrate that Y14 undergoes LLPS through multivalent electrostatic interactions mediated by its highly charged IDRs in vitro and that LLPS of Y14 may in part contribute to its function in DNA damage repair.
DISCUSSION
LLPS mediated by the interaction between oppositely charged IDRs is, in general, sensitive to pH and salt. For example, formation of p53 droplets, which is driven by its terminal domains with opposite net charges, is pH dependent and suppressed by high salt (Kamagata et al. 2020). Y14 was similar to p53 with respect to pH-sensitive droplet formation (Fig. 2), indicating that LLPS of Y14 largely involves electrostatic interactions between IDRs. However, in contrast to p53, Y14 droplets became larger with increasing KCl concentration up to 100 mM, although higher concentrations of KCl completely suppressed droplet formation (Fig. 3). Therefore, in line with the observation that 1,6-hexanediol disrupted Y14 droplets, a moderate level of KCl may stabilize certain intramolecular or intermolecular interactions of Y14, which involve the hydrophobic or aromatic residues in the IDRs or the RRM (Fig. 8A). Such interactions may even become a major driving force for LLPS of ΔN1N3, which retained only a few negatively charged residues in the IDRs (Fig. 4). Besides, the RRM has a net negative charge (Fig. 8B). Therefore, it is possible that the RRM participates in LLPS of full-length or truncated Y14 to different extents or in different aspects.
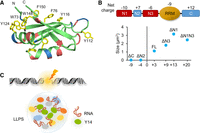
Model for LLPS of Y14. (A) Ribbon diagram showing the RRM of Y14, in which seven exposed aromatic side chains are presented. (B) Diagram shows net charge of each charged region of Y14. Note that 16 negatively charged residues and seven positively charged residues present in the RRM give it a net charge of approximately −9 at pH 7. Plot graph shows total net charge of the IDRs of indicated Y14 versions against their droplet size (the droplet size of ΔN1N3 was measured at pH 7.5 and all others at pH 6.5). (C) Y14 in concert with lncRNAs (such as HOTAIRM1) undergoes LLPS during DNA damage (lightning bolt) to facilitate repair. Colored ovals except for green (Y14) represent Y14- or lncRNA-associated DNA repair factors.
Y14 was found to share several LLPS features with a group of proteins containing alternating charge blocks, such as NPM1, G3BP1, tau, and p53 (Mitrea et al. 2018; Boyko et al. 2019; Kamagata et al. 2020; Yang et al. 2020). Y14 is similar to G3BP1 in domain organization, in that both proteins have a single RRM flanked by charged IDRs. The carboxy-terminal positively charged IDR of both proteins can bind RNA nonspecifically and contribute to LLPS (Fig. 4). The amino-terminal domain of Y14 is similar to that of NPM1, which has several positively charged and negatively charged regions that have an alternating arrangement (Mitrea et al. 2016, 2018). Electrostatic interactions between oppositely charged motifs of NPM1 drive homotypic LLPS. Moreover, NPM1 undergoes heterotypic LLPS with its interacting partners such as ribosomal proteins and the nonribosomal protein Surfeit locus protein 6 (SURF6) that contain Arg-rich linear motifs (Mitrea et al. 2018). Therefore, our result that the carboxy-terminal peptide of human and Drosophila Y14 restored LLPS of ΔC or ΔN2 (Fig. 5) is similar to the scenario of the heterotypic condensates of NPM1 and its partners. The tau protein can be roughly divided into the amino-terminal negatively charged domain and carboxy-terminal positively charged domain (Boyko et al. 2019). Neither can form phase-separated droplets, but the mixture of the two individual domains undergoes LLPS, as was observed for Y14, indicating that LLPS for these two proteins may occur via similar mechanisms.
Charge patterning and phosphorylation of IDRs affect LLPS. The C. elegans RNA helicase LAF-1 contains mixed-charge residues in its IDRs; its phase separation is enhanced by experimentally shuffling a portion of the charged residues in the IDR, which increases charge segregation (Schuster et al. 2020). This observation supports charge segregation as an important factor for Y14 LLPS. Also notable is that hyperphosphorylation of the large nucleolar protein Ki-67 during mitosis promotes its LLPS by creating alternating charge blocks (Yamazaki et al. 2022). In contrast, hyperphosphorylation reduces the propensity of NPM1 to undergo LLPS by disrupting charge segregation (Yamazaki et al. 2022). Therefore, it is possible that phospho-mimicking mutations of Y14 at the carboxy-terminal RS dipeptides disrupted charge homogeneity and hence suppressed LLPS (Fig. 4). Many splicing factors containing Arg-enriched mixed-charge domains form speckled structures in the nucleus. Interestingly, increasing negative or positive charge of such domains, respectively, abolishes or enhances protein condensation in speckles (Greig et al. 2020). It appears that the overall net charge of the Y14 IDRs had a substantial influence on LLPS, in that Y14 with a net negative charge prohibited LLPS whereas a net positive charge promoted LLPS (Fig. 8B). Moreover, a greater net positive charge in general increased the droplet size. ΔN1N3 was exceptional because its LLPS may involve a different spectrum of interactions, which affect droplet size.
LLPS of Y14 was independent of RNA. Therefore, Y14–W73V could form droplets, albeit of smaller size than those formed with wild-type Y14 (Fig. 6). Wild-type Y14, but not W73V, could co-condensate with RNA. Moreover, HOTAIRM1 promoted LLPS of Y14, supporting their coordinated function in DNA damage repair (Chuang et al. 2023). As has been observed for several RBPs such as Fus and hnRNP A1 (Maharana et al. 2018), low RNA:protein ratios could promote LLPS of Y14, whereas higher concentrations of RNA inhibited LLPS. Such a regulatory role of RNA in phase separation has been implicated in various cellular processes including transcriptional initiation and stress-granule formation (Yang et al. 2020; Henninger et al. 2021). Therefore, whether the level of DSB-induced transcripts (Michelini et al. 2017) also modulates phase separation of Y14 at DNA damage sites warrants further investigation. In conclusion, this study implies that Y14 may, in concert with its associated lncRNA HOTAIRM1, function in DSB repair via LLPS (Fig. 8C).
In this study, we demonstrated Y14 co-phase separation with RNA in vitro, which may contribute to its role in DNA repair. However, it was intriguing whether the Y14/Magoh heterodimer may undergo LLPS. Purified S-/His-tagged Magoh formed aggregates (Supplemental Fig. 5A). After fluorescent labeling, we observed that Magoh no longer formed aggregates when mixed with Y14, and instead formed co-condensates with Y14, indicating heterotypic LLPS of the Y14/Magoh complex (Supplemental Fig. 5A). Therefore, whether this co-condensate formation occurs during EJC assembly in perispeckles (Daguenet et al. 2012) or in the cytoplasm remains to be investigated.
MATERIALS AND METHODS
Plasmids
The pET29b-S-Y14-His vector encoding S- and 6 × His-tagged Y14 was as previously described (Hsu et al. 2005). The expression vectors of pEGFP-Y14 and pcDNA-FLAG-Y14 have been described (Chuang et al. 2013). All mutations (S166,168D, W73V) and truncations (Supplemental Table S2) were generated by PCR-based mutagenesis/deletion. The sequence of the resulting vectors was confirmed. The pET29b-S-Magoh-His vector was described in Chuang et al. (2011). For in vitro transcription, the cDNAs encoding full-length HOTAIRM1 or fragments (HM1.1, 1–217 HM1.2, 221–395; HM1.3, 396–631; HM1.4, 632–682; HM1.5, 882–1044) were each inserted into the pGEM-T vector (Promega). The flanking sequence was shortened by restriction digestion with NsiI/NcoI or ApaI/SacII, following by blunt-ending and religation. The sequence of all the constructs was confirmed by Sanger sequencing.
Protein expression and purification
All recombinant Y14 versions used in this study contained dual tags. Y14 was overexpressed in E. coli strain BL21 (DE3). Transformed bacteria were grown at 37°C in Luria broth (LB) containing kanamycin. At OD600 0.4–0.6, 1 mM isopropyl β-d-1-thiogalactopyranoside (IPTG) was added. Bacteria were subsequently shifted to 15°C for 12–16 h. Cell pellets were harvested and resuspended with the resuspension buffer (0.5 M NaCl, 20 mM Tris-HCl, pH 7.9), and sonicated, followed by centrifugation at 13,000 rpm for 30 min at 4°C. The supernatant was passed over a Ni Sepharose 6 Fast Flow medium (GE Healthcare), washed with the resuspension buffer containing 60 mM imidazole and eluted with the same buffer containing 500 mM imidazole. Purified full-length Y14 was dialyzed against the storage buffer (50 mM KCl, 20 mM HEPES pH 7.9, 20% glycerol) and stored at −80°C. Recombinant S-/His-tagged Magoh was also overexpressed in E. coli and purified through Ni Sepharose as above. For truncated Y14 proteins and Magoh, storage buffer was listed in Supplemental Table S2. Proteins were analyzed on SDS-polyacrylamide gel electrophoresis followed by TOOLStart Blue (BIOTOOLS) staining. Nucleic acid contamination was evaluated on a Nanodrop.
Peptides
Four peptides used in LLPS included HsC23 (RGPPKGKRRGGRRRSRSPDRRRR), DmC14 (KGPKRVKKSEKRRR), CeC9 (KGKKTSGKR), and E7N (MHGDTPTLHEYMLDLQPETT). All peptides were dissolved in a 50 mM KCl, 20 mM HEPES pH 7.9 buffer.
Droplet formation assay
Recombinant Y14 proteins were mixed with buffers containing 20 mM MES/HEPES and different concentrations of KCl at different pHs. For protein/RNA co-condensation analysis, proteins and RNA were mixed before dilution into the LLPS reaction buffer. For Y14/Magoh co-condensation analysis, recombinant Y14 (labeled: unlabeled = 1:200) and Magoh (labeled: unlabeled = 1:100) were mixed in a 300 mM KCl/pH7.9 buffer and incubated on ice for 3 h prior dilution into the LLPS reaction buffer. After dilution, samples were incubated for 2 h at room temperature and then placed onto the coverslip for visualization using a Leica microscope (DMI6000B) with a 100× oil objective under the differential interference contrast (DIC) or fluorescence mode. To analyze Y14 LLPS, 1,6-hexanediol and PEG4000 were purchased from Merck.
Turbidity assay
Protein samples were prepared as described above. OD400 of 125 µL samples in microcuvettes (Taitek Scientific) was measured using a DU 800 Spectrophotometer (Beckman).
Fluorescent labeling of proteins
Proteins were fluorescently labeled using Alexa Fluor 488 or Alexa Fluor 647 Conjugation Kit (Fast)-Lightning-Link (Abcam). As for Y14/Magoh co-condensation analysis, Y14 was labeled using Alexa Fluor 488 C5 Maleimide (Thermo Fisher Scientific) according to the manufacturer's instructions. For visualization, the unlabeled proteins were mixed with the labeled protein with the ratio 50:1 unless otherwise stated.
Electrophoretic mobility shift assay (EMSA)
For in vitro transcription, the pGEM vectors each encoding a HOTAIRM1 fragment were linearized with NotI (for Sp6 transcription) or SacII (for T7 transcription) (New England Lab) and used as template. Radiolabeled HOTAIRM1 fragments were synthesized using T7 or Sp6 RNA polymerase (Promega) according to the manufacturer's instructions. In brief, each transcription reaction contained 0.5 mM ATP, CTP and GTP, 0.1 mM UTP, and 3.3 µM of α32P-UTP (3000 Ci/mmol, 10 mCi/ml, PerkinElmer). RNA was gel purified. The length of HOTAIRM1 fragments containing the flanking linker regions ranged from 214 to 289 nt. The specific radioactivity of in vitro transcribed RNAs was ∼3 × 106 cpm/µg.
For EMSA, 5 × 104 cpm (17–26 nM) 32P-labeled HOTAIRM1 fragment was incubated with Y14 in a buffer containing 50 mM HEPES (pH 7.0), 150 mM NaCl, 1 mM DTT, 10% glycerol, 0.5 mg/mL BSA on ice for 20 min. The reaction was mixed with the loading dye containing 0.01% xylene cyanol, 0.01% bromophenol blue and separated by electrophoresis on a 4% polyacrylamide gel in Tris-Borate-EDTA buffer. Gels were vacuum-dried and imaged on a Typhoon scanner (GE Healthcare). Quantification of radiolabeled RNA was performed using ImageQuantTL software (GE Healthcare).
Cy5-labeling of RNA
Both 5′-Cy5 labeled and unlabeled U25 and A25 oligoribonucleotides were purchased from Genomics. For in vitro transcription, the HOTAIRM1 or fragment vectors were linearized with an appropriate restriction enzyme and used as the template. The reaction containing the template, a mixture of nucleotides (2 mM ATP, CTP and GTP, and 1.9 mM UTP/0.1 mM of Cy5-UTP [Enzo] for full-length or 1.8 mM UTP/0.2 mM of Cy5-UTP for fragments) and T7 or Sp6 RNA polymerase (Promega) was carried out according to the manufacturer's instructions. After phenol–chloroform extraction, RNA was precipitated by ethanol.
Immunofluorescence and fluorescence recovery after photobleaching (FRAP)
U2OS cells were cultured in Dulbecco's modified Eagle's medium supplemented with penicillin-streptomycin and 10% fetal bovine serum (Gibco), and transiently transfected with the vector encoding GFP-Y14 (wild-type or mutant) using lipofectamine 2000 (Thermo Fisher Scientific). The transfectants were observed using a laser-scanning confocal microscope (LSM 780, Carl Zeiss) coupled with an image analysis system. For FRAP, transfected U2OS cells were seeded in Chambered Cover glass (Thermo Fisher Scientific) for 48 h. Photobleaching was performed using a laser-scanning confocal microscope (LSM 780, Carl Zeiss) with a 488-nm laser diode. Images were continuously taken during 100 cycles for a total time of 200 sec. FRAP analysis and fluorescence intensity measurement were done using ZEN Black Software (Carl Zeiss).
Nonhomologous end joining (NHEJ) assays
HeLa cells were maintained as above for U2OS cells. To assess the NHEJ activity, 1 µg of the Cas9/sgHPRT expressing vector (sgHPRT targeting sequences: AAAGGGTGTTTATTCCTCA), 25 pmol of dsDNA oligonucleotides Insertion (Ins dsDNA-1: 5′-TTAATTGAGTTGTCATATGTTAATAACCGG-3′; Ins dsDNA-2: 5′-ACCGTTATTAACATATGACAACTCAATTA-3′, Du et al. 2018), 40 pmol Y14 targeting siRNA (5′-ggguauacucuaguugaaa-3′, Thermo Fisher Scientific) and 4 µg of siRNA-resistant Y14-expressing vectors were transfected into 2 × 105 HeLa cells (Chuang et al. 2023). Genomic DNA was collected 48 h post-transfection and extracted by PureLink Genomic DNA Mini Kit (Thermo Fisher Scientific). For qPCR, the reactions containing 100 ng genomic DNA, specific primers, and PerfeCTa SYBR Green FastMix PCR Reagent (Quanta Biosciences) were performed in a LightCycler 480 Real-Time PCR System (Roche). The primers used were Cas9/sgHPRT forward (5′-ATCCAATCAAATGTTTGTATCCTGT-3′), Ins (5′-GAGTTGTCATATGTTAATAACGG-3′) and reverse (5′-CCCTTCAATGTTTACTTTGTTCTGG-3′).
Immunoprecipitation and immunoblotting
For immunoprecipitation, transfected HEK293 cell lysates were incubated with anti-FLAG M2 affinity gel (Sigma) at 4°C for 2 h. Beads were then washed with NET-2 buffer (150 mM NaCl, 50mM Tris-HCl and 0.05% NP-40), followed by RNA extraction using TRIzol reagent (Thermo Fisher Scientific) or protein extraction using SDS-PAGE sample buffer. Antibodies against the following epitope or proteins were used for immunoblotting: FLAG (polyclonal, Sigma), Y14 (polyclonal, GeneTex), γH2AX (monoclonal, Millipore), histone 3 (polyclonal, Cell Signaling Technology), and GAPDH (monoclonal, ProteinTech).
Chromatin fractionation
Chromatin fractionation of transfected U2OS cells was essentially performed as described in Chuang et al. (2023). In brief, cells were incubated with the cytoskeleton (CSK) buffer containing 10 mM PIPES (pH 7.0), 100 mM NaCl, 300 mM sucrose, 3 mM MgCl2, and 0.7% Triton X-100 at 4°C for 3 min. After removal of the soluble fraction by centrifugation at 12,000g for 15 min, the chromatin-enriched fraction was resuspended in 500 mM NaCl-containing CSK buffer followed by incubation at 4°C for 15 min.
Quantification and statistical analysis
Statistical analyses were essentially performed by a two-tailed, unpaired Student's t-test using Prism (GraphPad) in this study. Data were presented as mean ± SD. P-values are indicated in figure legends.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
We thank Rita P.-Y. Chen (Institute of Biological Chemistry, Academia Sinica, Taipei) for discussion. We particularly thank Chun-Hao Su for extensive technical assistance as well as Wei-Chi Ke for plasmid construction, and thank the Common Equipment Core of the Institute of Biomedical Sciences and the Biophysics Core of the Institute of Biological Chemistry for technical assistance. This study was supported by Academia Sinica Investigator Award (AS-IA-107-L04) to W.-Y.T.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079514.122.
-
Freely available online through the RNA Open Access option.
- Received November 9, 2022.
- Accepted March 13, 2023.
This article, published in RNA, is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Chia-Lin Yu is the first author of this paper, “Co-phase separation of Y14 and RNA in vitro and its implication for DNA repair.” After graduating with a Master of Science degree in Chemistry and Biochemistry from the University of California, Santa Cruz, Chia-Lin joined Dr. Tarn's laboratory at the Institute of Biomedical Sciences, Academia Sinica in Taiwan as a research assistant.
What are the major results described in your paper and how do they impact this branch of the field?
We unveiled the mechanism of liquid–liquid phase separation (LLPS) of Y14 and its ability to coalesce with RNA into phase-separated droplets. Re-entrant phase transition has also been observed as the concentration of RNA increases, implying a role of RNA in regulating condensates formation.
What led you to study RNA or this aspect of RNA science?
My research interest in RNA started during my master's program, where I learned that only 1%–2% of the genome encodes proteins, whereas a large portion of the genome is transcribed into RNA. This made me curious about the functions of these noncoding RNAs. LLPS is also an emerging research area nowadays and studies have shown that noncoding RNA could also participate in various processes through LLPS. I find it very interesting in studying how RNA undergoes phase separation and how it facilitates cellular processes via LLPS.
During the course of these experiments, were there any surprising results or particular difficulties that altered your thinking and subsequent focus?
We were surprised to find out that ΔN1N3 could form droplets on its own since it lacks the negatively charged residues for electrostatic interaction. We then searched for the possibility of other forces that underlie its phase transition. When adding 1,6-hexanediol to preformed ΔN1N3 droplets, the droplets started to dissolve, indicating the involvement of hydrophobic interactions. Other types of driving forces and how these forces participate in Y14 LLPS need further investigation.
Are there specific individuals or groups who have influenced your philosophy or approach to science?
I'm lucky to have joined Dr. Tarn's laboratory, where every member of our group supports me when I encounter difficulties. They are incredible mentors since their expertise covers a wide range of topics. Through coworking in our group I became more logical and critical in my thinking in aspects of my research.








