
Kinetic and structural insights into the requirement of fungal tRNA ligase for a 2′-phosphate end
- Corresponding author: shumans{at}mskcc.org
-
Handling editor: Eric Phizicky
Abstract
Fungal RNA ligase (LIG) is an essential tRNA splicing enzyme that joins 3′-OH,2′-PO4 and 5′-PO4 RNA ends to form a 2′-PO4,3′-5′ phosphodiester splice junction. Sealing entails three divalent cation-dependent adenylate transfer steps. First, LIG reacts with ATP to form a covalent ligase-(lysyl-Nζ)-AMP intermediate and displace pyrophosphate. Second, LIG transfers AMP to the 5′-PO4 RNA terminus to form an RNA-adenylate intermediate (A5′pp5′RNA). Third, LIG directs the attack of an RNA 3′-OH on AppRNA to form the splice junction and displace AMP. A defining feature of fungal LIG vis-à-vis canonical polynucleotide ligases is the requirement for a 2′-PO4 to synthesize a 3′–5′ phosphodiester bond. Fungal LIG consists of an N-terminal adenylyltransferase domain and a unique C-terminal domain. The C-domain of Chaetomium thermophilum LIG (CthLIG) engages a sulfate anion thought to be a mimetic of the terminal 2′-PO4. Here, we interrogated the contributions of the C-domain and the conserved sulfate ligands (His227, Arg334, Arg337) to ligation of a pRNA2′p substrate. We find that the C-domain is essential for end-joining but dispensable for ligase adenylylation. Mutations H227A, R334A, and R337A slowed the rate of step 2 RNA adenylation by 420-fold, 120-fold, and 60-fold, respectively, vis-à-vis wild-type CthLIG. An R334A-R337A double-mutation slowed step 2 by 580-fold. These results fortify the case for the strictly conserved His–Arg–Arg triad as the enforcer of the 2′-PO4 end-specificity of fungal tRNA ligases and as a target for small molecule interdiction of fungal tRNA splicing.
Keywords
INTRODUCTION
tRNA splicing was discovered in 1978 by the Rutter, Goodman, and Abelson laboratories when it became apparent that the yeast genes encoding certain tRNAs contained intervening sequences (introns) that were transcribed but not present in mature tRNAs (Knapp et al. 1978; O'Farrell et al. 1978; Valenzuela et al. 1978). At least one intron-containing pre-tRNA is present in all eukaryal taxa, inserted after nt 37 of the anticodon loop. The prevalence of tRNA introns within a given eukaryal taxon spans a wide range (Chan and Lowe 2016). At the high end, 128/131 (98%) of tRNA genes contain introns in the fungal pathogen Cryptococcus neoformans. At the other extreme, only 1/67 of the tRNA genes in the protozoan pathogen Trypanosoma brucei has an intron. All eukarya exploit a conserved mechanism of RNA breakage by a tRNA splicing endonuclease (TSEN) to excise the intron from the pre-tRNA (Hayne et al. 2023). Dual TSEN incision at the exon–intron junctions leaves 2′,3′ > p termini and 5′-OH termini on the tRNA halves and on the excised intron.
Nature has evolved two entirely different RNA repair pathways by which the broken tRNA halves are joined to form the mature spliced tRNA (Shuman 2023). Fungi and plants rectify the tRNA breaks via a multistep, multienzyme “healing and sealing” pathway in which: (i) the 2′,3′ > p end is hydrolyzed to a 3′-OH,2′-PO4 by a cyclic phosphodiesterase (CPD); (ii) the 5′-OH end is phosphorylated by an NTP-dependent polynucleotide kinase (KIN); (iii) the 3′-OH,2′-PO4 and 5′-PO4 ends are sealed by an ATP-dependent RNA ligase (LIG) to form an unconventional 2′-PO4,3′-5′ phosphodiester at the splice junction (Fig. 1A); and (iv) the 2′-PO4 mark at the splice junction is removed via transfer to NAD +. In plants and in most fungi, the LIG, KIN, and CPD catalytic modules are linked in cis in a single polypeptide (Fig. 1B). In contrast, metazoa and archaea use a completely different pathway in which the 2′,3′ > p and 5′-OH ends are joined by a GTP-dependent RNA ligase, RtcB, via an RNA3′ppG intermediate, without an intervening 5′ end-healing step. The stark differences in the chemistry of tRNA break repair, and the entirely different structures of the tRNA ligase enzymes in fungi versus mammals, advocate for tRNA splicing as a target for antifungal drug discovery.
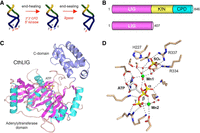
Domain structure and candidate 2′-PO4 binding site of Chaetomium thermophilum LIG (CthLIG). (A) Illustration of the end-healing and end-sealing phases of the fungal tRNA splicing pathway. (B) Cartoon depiction of the domain organization of full-length 846-aa C. thermophilum Trl1 consisting of N-terminal ligase (LIG), central kinase (KIN), and C-terminal cyclic phosphodiesterase (CPD) domains. The N-terminal 407-aa segment comprising an autonomous LIG enzyme is the subject of the present study. (C) The tertiary structure of CthLIG is shown with the N-terminal adenylyltransferase module colored by secondary structure (magenta β strands and cyan α helices) and the unique C-terminal domain (aa 328-406) colored blue. ATP bound to the adenylyltransferase domain is depicted as a stick model. (D) View of the CthLIG active site depicting atomic contacts to ATP, two Mn2+ ions (green spheres), and metal-bound waters (red spheres). The image highlights a sulfate anion (stick model) bound to the C-domain, adjacent to the catalytic metal (Mn1), that is thought to mimic the 2′-PO4 of the RNA substrate. Sulfate-binding residues His227, Arg334, and Arg337 were mutated to alanine in the present study.
Sealing of RNA ends by fungal and plant LIG enzymes proceeds via three divalent cation-dependent adenylate transfer steps. First, LIG reacts with ATP to form a covalent LIG-(lysyl-Nζ)-AMP intermediate and displace pyrophosphate. Second, LIG transfers AMP to the 5′-PO4 RNA terminus to form an RNA-adenylate intermediate (A5′pp5′RNA). Third, LIG directs the attack of an RNA 3′-OH on AppRNA to form the splice junction and displace AMP. A defining feature of fungal and plant tRNA ligases, vis-à-vis other ATP-dependent RNA ligases (e.g., T4 RNA ligases 1 and 2), is the requirement for a terminal 2′-PO4 to synthesize a 3′–5′ phosphodiester bond (Konarska et al. 1982; Greer et al. 1983; Remus and Shuman 2013; Ghosh et al. 2024). A better understanding of how the 2′-PO4 contributes to catalysis of end-joining and of the structural basis for 2′-PO4 recognition would aid in the design/discovery of inhibitors/poisons of the ligation reactions.
Toward that end, crystal structures of a catalytically active tRNA ligase from the fungus Chaetomium thermophilum (CthLIG) have been solved at discrete steps along the reaction pathway: the covalent LIG-(lysyl-Nζ)-AMP•Mn2+ intermediate; a LIG•ATP•(Mn2+)2 Michaelis complex (Fig. 1C,D); and a binary complex with AMPCPP lacking a divalent metal (Banerjee et al. 2019; Peschek and Walter 2019). These structures highlighted a two-metal mechanism whereby a pentahydrated metal complex stabilizes the transition state of the ATP α-phosphate and a second metal bridges the β and γ phosphates to help orient the pyrophosphate leaving group (Fig. 1D). The CthLIG tertiary structure consists of two domains (Fig. 1C): (i) an N-terminal adenylyltransferase module (from aa 39 to 327), homologous to the N-terminal adenylyltransferase domain of T4 RNA ligase 1 (El Omari et al. 2005; Unciuleac et al. 2017), that includes six peptide motifs (I, Ia, III, IIIa, IV, and V) conserved among ATP-dependent polynucleotide ligases, constituents of which engage the ATP substrate and the catalytic metal ion; and (ii) a C-terminal module, composed of five α-helices, that is conserved among fungal LIG enzymes. This distinctive C-domain instates CthLIG as the founder of an Rnl6 clade of RNA ligase (Banerjee et al. 2019). It is suggested that the C-domain is a determinant of RNA end specificity, insofar as a LIG-bound sulfate anion engaged by amino acids Arg334 and Arg337 in the C-terminal domain, and by His227 in the adenylyltransferase domain, is thought to be a mimetic of the RNA terminal 2′-PO4 (Fig. 1D).
In the present study, we conduct a mutational analysis of CthLIG focusing on the role of the C-domain and the enzymic sulfate ligands in promoting ligation of a pRNA2′p substrate in vitro. We report that the isolated N-terminal domain of CthLIG reacts with ATP to form the covalent CthLIG–AMP intermediate but is not adept at ligation. We find that single alanine mutations of His227, Arg334, and Arg337 slow step 2 RNA adenylation by 420-fold, 120-fold, and 60-fold, respectively, and that an R334A–R337A double-mutation slows step 2 by 580-fold. This His–Arg–Arg triad is conserved in the tRNA ligases of all high-priority human fungal pathogens designated by the World Health Organization.
RESULTS
The C-domain of Chaetomium thermophilum LIG is essential for end joining
Chaetomium thermophilum Trl1 is an 846-aa tRNA splicing enzyme composed of N-terminal ligase (LIG), central kinase (KIN), and C-terminal CPD domains (Fig. 1B; Banerjee et al. 2019; Peschek and Walter 2019). The N-terminal segment (aa 1–407), referred to henceforth at CthLIG, is an autonomous 3′-OH,2′-PO4/5′-PO4 end-sealing enzyme (Fig. 1B; Banerjee et al. 2019). Here, we produced recombinant wild-type CthLIG and a truncated version CthLIG(1-327) in Escherichia coli as His-tagged fusions and purified them from soluble extracts by sequential Ni-affinity chromatography and Superdex-200 gel filtration steps. SDS-PAGE indicated that both preparations were pure, and that the C-domain deleted version of CthLIG displayed the expected lower apparent molecular weight (Fig. 2A).
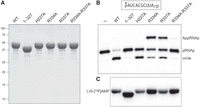
Structure-guided mutagenesis of CthLIG. (A) Aliquots (10 µg) of the purified full-length wild-type CthLIG, the indicated full-length alanine mutants, and the N-terminal adenylyltransferase domain (1–327) were analyzed by SDS-PAGE. The Coomassie blue-stained gel is shown. The positions and sizes (kDa) of marker polypeptides are indicated on the left. (B) Ligation reaction mixtures (10 µL) containing 50 mM Tris-HCl, pH 8.0, 50 mM NaCl, 2 mM DTT, 10 mM MgCl2, 100 µM ATP, 1 pmol (0.1 µM) 32P-labeled 10-mer pRNA2′p (shown at the top, with the 32P-label indicated by filled circle [•]), and either no enzyme (lane –), or 1 pmol (0.1 µM) of the indicated CthLIG protein were incubated at 37°C for 30 min. The reactions were quenched with an equal volume of 95% formamide/50 mM EDTA, and the products were analyzed by electrophoresis (at 58 W constant power) through a 40 cm 20% polyacrylamide gel containing 7 M urea in 45 mM Tris-borate, 1 mM EDTA. The radiolabeled RNAs were visualized by scanning the gel with a Typhoon FLA-7000 imaging device. The positions of the 5′-PO4,2′-PO4 RNA substrate (pRNAp), the 5-adenylylated intermediate (AppRNAp), and the 10-mer circle product of intramolecular ligation are indicated on the right. (C) Ligase adenylylation. Reaction mixtures (10 µL) containing 50 mM Tris-HCl, pH 8.0, 10 mM MgCl2, 50 µM [α32P]ATP, and 25 µM (250 pmol) of the indicated CthLIG protein were incubated at 37˚C for 5 min, then quenched with SDS, and analyzed by SDS-PAGE. A scan of the gel is shown.
To assay ligase activity, we deployed a 5′ 32P-labeled synthetic 10-mer RNA with a 3′-OH,2′-PO4 end (Ghosh et al. 2024). The pRNA2′p substrate (100 nM) was reacted for 30 min at 37˚C with CthLIG (100 nM) in the presence of magnesium and 100 µM ATP. The products were analyzed by urea-PAGE and visualized by scanning with a phosphorimager (Fig. 2B). Wild-type CthLIG converted 93 ± 1.5% of the pRNA2′p substrate into a 10-mer circular RNA—formed by intramolecular ligation—that migrated ahead of the substrate strand. In contrast, CthLIG(1-327) circularized only 1.0 ± 0.25% of the pRNA2′p substrate (Fig. 2B). (These extents of product formation are the averages of four replicate experiments ± SEM.)
Because end joining depends on ligase-adenylylation (step 1 of the ligase pathway), we assayed the adenylyltransferase activity of CthLIG and CthLIG(1-327). In both cases, their reaction with magnesium and 50 µM [α32P]ATP resulted in formation of a covalent CthLIG–[32P]AMP adduct that migrated at the expected size during SDS-PAGE (Fig. 2C). We conclude that the N-terminal domain of CthLIG is an autonomous adenylyltransferase and that the C-domain of CthLIG is essential for one or both ensuing steps of the ligation pathway: RNA adenylylation and phosphodiester synthesis.
Effect of mutating the CthLIG sulfate-binding residues on end joining
Guided by the CthLIG crystal structure, we produced and purified mutated versions of CthLIG in which sulfate ligands His227, Arg334, and Arg337 were replaced individually by alanine. We also produced and purified a R334A-R337A double-mutant. The purity of the recombinant Ala-mutants was equivalent to that of wild-type CthLIG (Fig. 2A). The H227A, R334A, R337A, and R334A–R337A proteins were all active in ligase adenylylation (Fig. 2C), consistent with the CthLIG structure, which showed that these residues do not contact ATP or the catalytic metal ion (Fig. 1B).
Initial assays of pRNAp ligation by the four alanine mutants in parallel with wild-type CthLIG and CthLIG(1-327) were highly instructive, as follows. The H227A enzyme circularized 68 ± 1.1% of the input substrate (Fig. 2B). In contrast, the R334A and R337A mutants circularized only 14 ± 0.3% and 23 ± 0.8% of the input pRNA2′p, respectively, while accumulating a slower-migrating AppRNA2′p intermediate to an extent of 63 ± 2.9% and 38 ± 1.3% of total RNA, respectively (Fig. 2B). Mutating both Arg334 and Arg337 had an additive effect, such that AppRNA2′p and ligated circles comprised 1.4 ± 0.37% and 0.89 ± 0.23% of the total RNA, respectively (Fig. 2B). (These extents of RNA-adenylate and RNA circle formation are the averages of four replicate experiments ± SEM.)
Characterization of the ligation reaction products
To affirm the identities of the 32P-labeled reaction products formed by wild-type CthLIG and the R334A mutant, we tested their sensitivity to alkaline phosphatase (CIP). As expected, the 5′ 32P phosphomonoester of the input pRNA2′p substrate was hydrolyzed completely by CIP (Fig. 3A,C). In the case of the AppRNAp product formed by R334A, the radiolabel in the 5′ AppA– phosphoanhydride structure is protected from CIP digestion but the unlabeled 2′-PO4 is susceptible to CIP (Fig. 3C), thereby leading to complete conversion of AppRNA2′p to a slower-migrating AppRNAOH (Fig. 3A). Ligation of the pRNAp substrates embeds the 32P-labeled phosphate in a CIP-resistant 3′,5′ phosphodiester linkage (Fig. 3C). The vicinal unlabeled 2′-PO4 at the splice junction is known to be relatively refractory to hydrolysis by CIP (McCraith and Phizicky 1990). Thus, only a minority of the monomer circle produced during reaction of wild-type CthLIG with the 2′-PO4 substrate was converted by CIP into a slower-moving circle with a 2′-OH (denoted by the arrowhead in Fig. 3A,B). Tpt1 is a dedicated NAD +-dependent RNA 2′-phosphotransferase that removes the junction 2′-PO4 to yield the mature tRNA splice junction (McCraith and Phizicky 1991). Tpt1 accomplishes this in two chemical steps in which: (i) NAD+ reacts with the RNA 2′-phosphate to expel nicotinamide and generate a 2′-phospho-ADP-ribosylated RNA intermediate; and (ii) transesterification of the ADP-ribose 2′-OH to the tRNA 2′-phosphate displaces the 2′-OH tRNA product and generates ADP-ribose-1′′,2′′-cyclic phosphate (Culver et al. 1993; Spinelli et al. 1999; Munir et al. 2018). Here, we see that treatment of the ligated circle formed by wild-type CthLIG with purified recombinant Tpt1 and 1 mM NAD+ results in complete removal of the 2′-PO4 to form a slower-migrating circle with a 2′-OH (Fig. 3B,C).
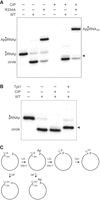
Analysis of the ligation reaction products. (A) Product analysis via treatment with CIP (calf intestine alkaline phosphatase). Ligation reactions containing 0.5 pmol 5′ 32P-labeled pRNA2′p and 10 pmol CthLIG (where indicated by +) were incubated at 37˚C for 30 min. The mixtures were then incubated for 10 min at 37˚C with 10 U CIP (purchased from NEB) where indicated by + above the lanes. The mixtures were quenched and then analyzed by urea-PAGE. (B) Product analysis via treatment with Tpt1 (RNA 2′-phosphotransferase). Ligation reactions containing 0.5 pmol 5′ 32P-labeled pRNA2′p and 10 pmol CthLIG (where indicated by +) were incubated at 37˚C for 30 min. The mixtures were then incubated for 30 min at 37˚C with 5 pmol Runella slithyformis Tpt1 (RsTpt1) and 1 mM NAD +. The mixtures were quenched and then analyzed by urea-PAGE. The 5′ 32P-phosphate is denoted by filled circle (•) in panels A and B. The ligated circular RNA with a 2′-OH splice junction formed after treatment with Tpt1 is indicated by the arrowhead at right in panels A and B. (C) Schematic of steps 2 and 3 of the ligation reaction and the species generated by treatment with CIP and Tpt1.
Reaction of CthLIG with 2′-PO4 versus 2′-OH substrates
The 10-mer pRNA2′p substrate (1 pmol; 0.1 µM) was reacted for 5 min with 5 mM Mg2+ and increasing amounts of CthLIG in the absence of exogenous ATP. Under these conditions, the extent of ligation depends on the level of catalytically active covalent CthLIG–AMP adduct in the enzyme preparation. CthLIG (at ≥10 pmol; 1 µM) converted 95% the 2′-PO4 RNA substrate into ligated circles (Fig. 4). From the slope of the titration curve between 0 and 1 pmol CthLIG, we calculated that ∼60% of the enzyme preparation was CthLIG–AMP (Fig. 4). We conducted parallel enzyme titrations in which CthLIG was reacted for 20 min with an otherwise identical 5′ 32P-labeled 10-mer pRNAOH substrate (1 pmol; 0.1 µM) lacking a terminal 2′-PO4 group. In this case, CthLIG (at ≥10 pmol) circularized only 0.12% of the input pRNAOH and generated AppRNA to an extent of no greater than 0.05% of total RNA at any point in the titration experiments (not shown). Thus, end sealing by CthLIG is stringently dependent on the 2′-PO4 group.
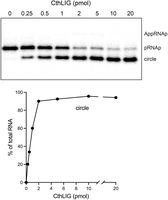
ATP-independent ligation by CthLIG–AMP. (Top panel) Reaction mixtures (10 µL) containing 50 mM Tris-HCl, pH 8.0, 2 mM DTT, 5 mM MgCl2, 1 pmol (0.1 µM) 32P-labeled 10-mer pRNA2’p, and CthLIG protein as specified were incubated at 37°C for 5 min and then quenched with formamide/EDTA. The mixtures were analyzed by urea-PAGE and the radiolabeled RNAs were visualized by scanning the gel. The positions of the pRNAp substrate, AppRNAp intermediate, and ligated circle product are indicated on the right. (Bottom panel) The extent of ligation is plotted as a function of input CthLIG. Each datum in the graph is the average of three independent titration experiments ± SEM. The error values are small, such that the error bars do not extent beyond the symbols.
Kinetics of single-turnover ligation
By omitting ATP and working at a 20-fold molar excess of wild-type CthLIG to 5′-32P-labeled pRNAp (corresponding to a 12-fold excess of LIG–AMP), we conducted a single-turnover kinetic analysis of the RNA adenylylation and phosphodiester formation steps. Wild-type CthLIG was added to individual prewarmed mixtures (five replicates per time point), and the reactions were quenched after 5, 10, and 15 sec with EDTA/formamide. An apparent reaction end point was achieved in 10 to 15 sec with 94% of the input linear RNA converted to ligated circle, while AppRNA2′p comprised only 0.3% of total RNA at the 5 and 10 sec times before declining to ∼0.03% at 15 sec (Fig. 5A). Plotting the sum of the RNA-adenylate intermediate and the ligated RNA product and a nonlinear least-squares regression fit of the data to a single exponential (one-phase association) in Prism yielded an apparent step 2 rate constant of 0.60 ± 0.042 sec−1 (Fig. 5A, right panel). A nonlinear least-squares regression fit of the data to a unidirectional two-step mechanism executed in Prism returned apparent step 2 (RNA adenylylation) and step 3 (phosphodiester formation) rate constants of 0.54 ± 0.027 sec−1 and 19 ± 64 sec−1, respectively (Fig. 5A, left panel). Whereas the two methods of data fitting provide a reliable value for kstep2, the large SEM for the calculated kstep3 value reflects the uncertainty arising from the paucity of RNA-adenylate intermediate. Nonetheless, it is clear that step 2 is rate-limiting and step 3 is much faster than step 2.

Kinetics of single-turnover ligation. (A) Individual replicate reaction mixtures (10 µL) containing 50 mM Tris-HCl, pH 8.0, 2 mM DTT, 5 mM MgCl2, 0.1 µM 5′ 32P-labeled 10-mer pRNAp substrate, and 2 µM wild-type CthLIG were incubated at 37˚C. The reactions were initiated by adding CthLIG to a prewarmed reaction mixture. The reactions were quenched after 5, 10, or 15 sec with an equal volume of 95% formamide/50 mM EDTA. The products were analyzed by urea-PAGE. The distribution of radiolabeled RNAs, either the individual AppRNAp and the ligated circle species (left panel) or the sum of AppRNAp and ligated circle (right panel) is plotted as a function of reaction time. Each datum in the graph is an average of five replicate reactions ± SEM. (B–E) Reaction mixtures (100 µL) containing 50 mM Tris-HCl, pH 7.5, 5 mM MgCl2, 0.1 µM 5′ 32P-labeled 10-mer pRNAp substrate, and 2 µM mutant CthLIG as specified were incubated at 37˚C. The reactions were initiated by adding mutant CthLIG to a prewarmed reaction mixture. Aliquots (10 µL) were withdrawn before adding enzyme (time 0) or at the times specified after enzyme addition and quenched immediately with an equal volume of formamide/EDTA. The products were analyzed by urea-PAGE. The distribution of radiolabeled RNAs is plotted as a function of reaction time. Each datum in the graphs is an average of three independent experiments ± SEM. The data in the graphs in the right column were fit by nonlinear regression to a one-phase association in Prism. The apparent step 2 (RNA adenylylation) rate constants are shown. The data in the graphs in the left column were fit by nonlinear regression to a unidirectional two-step kinetic mechanism. The Prism-calculated step 2 and step 3 (phosphodiester synthesis) rate constants are shown. Because there was no detectable AppRNAp formed during the H227A reaction, panel E includes only a one-phase association fit of the data in the graph in the right column.
To gauge single-turnover ligation by the arginine mutants, 2 µM CthLIG was added to a prewarmed mixture containing 0.1 µM pRNAp, and aliquots were withdrawn at sequential times (between 30 sec and 30 min) and quenched with EDTA/formamide. The data plotted in Figure 5B–D are averages of three experiments. In the case of R334A, ligated product accumulated steadily over 5 min and attained an end point at 20–30 min, at which time 93% of the input RNA was sealed. RNA-adenylate intermediate peaked at 1 min, comprising 2.3% of total labeled RNA, and declined thereafter to ∼0.06% of total RNA (Fig. 5B). Fitting the sum of intermediate and product to a one-phase association yielded a step 2 rate constant of 0.29 ± 0.024 min−1 (Fig. 5B, right panel). Thus, based on the one-phase values for kstep2, the R334A mutation slowed RNA adenylylation by 120-fold vis-à-vis wild type. The data fit to a two-step mechanism returned apparent step 2 and step 3 rate constants of 0.28 ± 0.015 min−1 and 14 ± 10 min−1, respectively (Fig. 5B, left panel). The two-step data fits indicated that R334A slowed phosphodiester formation by ∼80-fold.
The kinetic profile of the R337A mutant displayed steady formation of ligated circles over 5 min, achieving an endpoint at 10 min with 94% of input RNA sealed (Fig. 5C). RNA-adenylate peaked at 0.5 min, comprising 0.9% of total labeled RNA. Fit to a one-phase association indicated a step 2 rate constant of 0.54 ± 0.018 min−1 (Fig. 5C). The data fit to a two-step mechanism returned step 2 and step 3 rate constants of 0.54 ± 0.013 min−1 and 28 ± 13 min−1, respectively (Fig. 5C). Compared to wild type, the R334A mutation slowed steps 2 and 3 by 60-fold and ∼40-fold, respectively.
Combining the R334A and R337A mutations exerted a more severe impact on the rate and extent of sealing than either of the single mutations. RNA-adenylate formation exceeded circularization at early times (0.5–5 min) and plateaued at a 10–20 min, with intermediate comprising 2.8% of total RNA. Ligated product accumulated slowly to the extent of 8.5% of total RNA at 30 min (Fig. 5D). Fitting the data to a one-phase association indicated that the reaction would react an end point with 13% of the input pRNAp being converted to AppRNAp plus ligated circle, with an apparent kstep2 of 0.063 ± 0.014 min−1 (Fig. 5D, right panel). The apparent rate of RNA adenylylation by the R334A–R337A double-mutant was 580-fold slower versus wild type and fivefold and ninefold slower than the R334A and R337A single mutants, respectively. Applying the endpoint value derived from the one-phase association model, a fit of the R334A–R337A data to a two-step kinetic mechanism yielded kstep2 and kstep3 values of 0.055 ± 0.003 min−1 and 0.11 ± 0.007 min−1, respectively (Fig. 5D, left panel). The modeled R334A–R337A step 3 rate was ∼10,000-fold slower than wild type and ∼130-fold and ∼250-fold slower than the R334A and R337A step3 rates, respectively.
The kinetic profile of single-turnover ligation by the H227A mutant revealed progressive accumulation of RNA circles between 2 and 30 min, attaining an end point of 88% ligation at 60 min (Fig. 5E). RNA-adenylate was undetectable. Fit to a one-phase association indicated a step 2 rate constant of 0.085 ± 0.004 min−1 (Fig. 5E). Thus, the loss of the imidazole moiety slowed the rate of RNA adenylation by ∼420-fold.
DISCUSSION
Three key themes emerge from our structure-function study of CthLIG, an exemplary fungal tRNA ligase. First, we establish that the N-terminal domain of CthLIG is an autonomous adenylyltransferase capable of reaction with ATP to form the covalent LIG-(lysyl-Nζ)–AMP intermediate, but unable to execute the RNA sealing steps. The signature C-domain of fungal tRNA ligases in thereby implicated in substrate RNA recognition. In this respect, fungal tRNA ligase resembles T4 RNA ligase 1, a bona fide tRNA repair enzyme (Shuman 2023). T4 Rnl1 is composed of an N-terminal adenylyltransferase domain fused to a unique C-terminal domain that defines the Rnl1 family of ATP-dependent RNA ligases (El Omari et al. 2005; Unciuleac et al. 2017). The T4 Rnl1 C-domain, which is dispensable for ligase adenylylation, confers specificity for tRNA as the substrate for end sealing whereby Rnl1 recognizes the tRNA tertiary structure (Wang et al. 2007: Nandakumar et al. 2008).
In contrast, fungal and plant tRNA ligases are directed to their substrates, not by the tRNA tertiary structure, but rather by recognition of the 3′-OH,2′-PO4 and 5′-PO4 RNA ends formed during the upstream end-healing steps of tRNA splicing (Nandakumar et al. 2008). This is affirmed here by our second key finding that CthLIG is incapable of ligating 3′-OH,2′-OH and 5′-PO4 RNA ends in the context of a 10-mer RNA that, when decorated with a 2′-PO4, is an excellent substrate for sealing by CthLIG. It is worth noting that, although plant and fungal ligases are unquestionably specific for 3′-OH,2′-PO4 ends, ligases from different taxa may vary in the stringency of this specificity. For example, when presented with the same 10-mer 2′-PO4, and 2′-OH RNAs used here, Mucor circinelloides RNA ligase (MciRNL) could circularize the 2′-OH RNA under single-turnover conditions, albeit very slowly. Comparing the kinetic profiles of the reactions of Mucor RNL with 2′-PO4 and 2′-OH substrates, we found that the 2′-PO4 enhanced the rates of step 2 RNA adenylylation and step 3 end-sealing by 125-fold and 6200-fold, respectively (Ghosh et al. 2024). Using 20-mer pRNA substrates to interrogate Arabidopsis thaliana tRNA ligase (AtRNL), it was found that a 2′-PO4 enhanced the rate of step 2 RNA adenylylation by 330-fold (Remus and Shuman 2013).
By implementing an alanine scan of the histidine and two arginine residues that coordinate a sulfate anion in the crystal structure of CthLIG, we provide evidence, via single-turnover kinetic analyses, in support of the hypothesis that these three amino acids are responsible for engagement of the essential 2′-PO4 end. In the single-turnover format of sealing by preformed LIG–AMP, the H227A, R334A, and R337A mutations exerted strong effects on step 2 kinetics without accumulating significant amounts of the AppRNAp intermediate (Fig. 5), thus signifying that step 2 continued to be the rate-limiting transaction. Yet, when ATP was included in the reaction, the R334A and R337A mutants generated high levels of AppRNAp (Fig. 2B). The likely reason for this discrepancy is that: (i) the de-adenylylated R334A and R337A proteins are prone to dissociate from the AppRNAp intermediate after execution of step 2, at which point they become re-adenylylated by reaction with ATP; and (ii) LIG–AMP is unable to bind AppRNAp because the adenylate pocket is occupied, resulting in free AppRNAp as an aborted dead-end product. In the absence of ATP, the nonadenylylated R334A and R337A enzymes can rebind to free AppRNAp and catalyze strand sealing. Prior studies have shown that abortive AppRNA or AppDNA species accumulate during reactions of mutant RNA/DNA ligases with ATP substrates containing sub-optimally poised 3′ and 5′ ends (Sriskanda and Shuman 1998; Yin et al. 2003, 2004; Zhu and Shuman 2007).
The CthLIG residues His227, Arg334, and Arg337 that are implicated presently in 2′-PO4 recognition are conserved in S. cerevisiae Trl1 (SceTrl1) as His193, Arg295, and Arg298, respectively. An alanine scan of the LIG module of SceTrl1 (aa 1-388) revealed that SceLIG mutants H193A and R298A were unable to complement growth of a budding yeast trl1Δ strain when coexpressed with the kinase-CPD module SceTrl1-(389-827) (Wang and Shuman 2005). The histidine and arginine were judged to be strictly essential for SceLIG activity in vivo, insofar as conservative mutations H193N and H193Q and R298K and R298Q were all lethal in vivo (Wang and Shuman 2005). Although these SceLIG mutants were not characterized biochemically, the mutational results in vivo are in accord with the catalytic defects observed for the corresponding CthLIG H227A and R337A proteins in vitro. In contrast, the SceLIG R295A mutant was active in vivo (Wang and Shuman 2005), whereas the corresponding CthLIG R334A mutant was severely defective for ligation in vitro when tested with the 10-mer pRNA2′p substrate. It is conceivable that LIG enzymes from different fungal taxa rely to different extents on individual amino acids that comprise the 2′-PO4 binding site. Alternatively, the mutation of the upstream arginine of the His–Arg–Arg triad to alanine might have a lesser impact on the joining of tRNA exons (the physiological substrates) than it does on circularization of the 10-mer pRNA2′p substrate deployed here.
Interference with the 2′-PO4 binding site of fungal tRNA ligase by a small molecule ligand that overlaps or occupies that site is a potential means to specifically interdict tRNA splicing without affecting the activity of RNA ligases that do not require a 2′-PO4 for end joining. This idea is plausible only if the His–Arg–Arg triad that we assign as the 2′-PO4 binding site of CthLIG is conserved in the tRNA ligases of fungal species that cause human disease. The World Health Organization has designated the fungal species shown in Figure 6, which can cause invasive, acute, and subacute systemic infections, as a priority public health concern and a focus for antifungal discovery (World Health Organization 2002). As shown in Figure 6, it is the indeed case that the histidine and two arginines are invariant in their presence and their locations (relative to adenylyltransferase motifs III and V) in all 18 priority fungal pathogens. The triad histidine is situated nine amino acids downstream from the signature motif III glutamate that coordinates the ribose 2′-OH of ATP and a water engaged to the catalytic metal ion (see Fig. 1D). The triad arginine residues are located just downstream from the two motif V lysine side chains that that engage the ATP α phosphate. This conservation bodes well for tRNA ligase as an antifungal target.
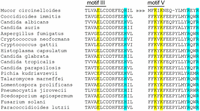
The His–Arg–Arg triad implicated in 2′-PO4 binding is conserved in the tRNA ligases of fungal pathogens. The amino acid sequences in and flanking adenylyltransferase motifs III and V are aligned for the tRNA ligase enzymes of 18 human fungal pathogens designated as public health priorities by WHO. The defining glutamate of motif III and two lysines of motif V are shaded yellow. The histidine adjacent to motif III (corresponding to CthLIG His227) and the two arginine residues downstream from motif V (corresponding to CthLIG Arg334 and Arg337) are shaded cyan.
MATERIALS AND METHODS
Recombinant Chaetomium thermophilum LIG proteins
Bacterial expression plasmid pET28-Trl1-LIG encodes CthLIG (aa 1–407) fused to an N-terminal hexahistidine tag under the control of a bacteriophage T7 promoter (Banerjee et al. 2019). Alanine mutations were introduced into the pET28-Trl1-LIG expression plasmid by two-stage overlap extension PCR with mutagenic primers. Plasmid pET28-Trl1-LIG(1-327) encodes a truncated version of CthLIG that includes the adenylyltransferase motifs but lacks the C-terminal module conserved among fungal LIG enzymes. The pET28-Trl1-LIG plasmids were transformed into E. coli BL21(DE3)-Codon Plus. Cultures (1 L) amplified from single kanamycin/chloramphenicol-resistant transformants were grown at 37°C in Luria Bertani medium containing 60 μg/mL kanamycin and 34 μg/mL chloramphenicol until the A600 reached 0.6–0.7. The cultures were chilled on ice for 1 h, adjusted to 2% (v/v) ethanol and 0.5 mM IPTG, and then incubated for 17 h at 18°C with constant shaking. Cells were harvested by centrifugation and stored at −80°C. All subsequent steps were performed at 4°C. Cells were thawed and resuspended in 25 mL of buffer A (50 mM Tris-HCl, pH 8.0, 1 M NaCl, 20 mM imidazole, and 10% glycerol) containing one complete EDTA-free protease inhibitor cocktail tablet (Roche). Lysozyme was added to a concentration of 1 mg/mL. After incubation for 30 min, the lysate was sonicated to reduce viscosity and the insoluble material was removed by centrifugation at 38,000g for 45 min. The supernatant was mixed for 1 h with 5 mL of Ni–nitrilotriacetic acid agarose resin (Qiagen) that had been equilibrated with buffer A. The resin was recovered by centrifugation and washed twice with 50 mL of buffer A, then with 20 mL of 50 mM Tris-HCl, pH 8.0, 3 M KCl, followed by 100 mL of buffer B (50 mM Tris-HCl, pH 8.0, 300 mM NaCl) containing 20 mM imidazole. The resin was centrifuged again, resuspended in 20 mL of buffer B with 20 mM imidazole, and poured into a column. The bound material was serially step-eluted with 5 mL of buffer B containing 50 mM imidazole, 15 mL of buffer B containing 150 mM imidazole, and 15 mL of buffer B containing 250 mM imidazole while collecting 5 mL fractions. The polypeptide compositions of the flow-through and eluate fractions were monitored by SDS-PAGE. The 50 mM imidazole and 150 mM imidazole eluate fractions containing CthLIG or CthLIG(1-327) were pooled and concentrated by centrifugal ultrafiltration [Amicon Ultra-15; 30 kDa cutoff for Trl1-LIG; 10 kDa cutoff for Trl1-LIG(1-327)] to 5 mL volume and then gel-filtered through a 125 mL 16/60 HiLoad Superdex 200 column (GE Healthcare) equilibrated in buffer D (20 mM HEPES–NaOH, pH 7.0, 200 mM NaCl, 1 mM DTT, and 5% glycerol) at a flow rate of 0.7 mL/min while collecting 2 mL fractions. The peak fractions were pooled, concentrated by centrifugal ultrafiltration, and stored at –80°C. Protein concentrations were determined with the Bio-Rad dye reagent using bovine serum albumin as the standard. The yields were 35–50 mg of protein per liter of bacterial culture.
5′ 32P-labeled oligoribonucleotide substrates
Synthetic 10-mer RNAs (5′-HOAUCACGCUUA) with either 2′-PO4 or 2′-OH ends (Ghosh et al. 2024) were 5′ 32P-labeled by reaction with phosphatase-dead T4 polynucleotide kinase Pnkp-D165N (Das and Shuman 2013) in the presence of [γ32P]ATP. The radiolabeled RNAs were gel-purified by electrophoresis through a 40 cm 20% nondenaturing polyacrylamide gel containing 45 mM Tris-borate, 1 mM EDTA. The radiolabeled RNAs were eluted from excised gel slices during overnight incubation in 10 mM Tris-HCl, pH 6.8, 1 mM EDTA, and then stored at −20°C.
ACKNOWLEDGMENTS
This work was supported by National Institutes of Health (NIH) grant R35-GM126945 (S.S.).
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080120.124.
- Received June 3, 2024.
- Accepted July 2, 2024.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








