
Structural and functional characterization of the SLA′ structure at the 3′ terminus of the Zika virus negative-strand intermediate
- 1Department of Biochemistry, McGill University, Montreal, Quebec, Canada, H3A 1A3
- 2Department of Microbiology and Immunology, University of British Columbia, British Columbia, Canada, V6T 1Z3
- 3Department of Microbiology and Immunology, McGill University, Montreal, Quebec, Canada, H3A 1A3
- Corresponding author: selena.sagan{at}ubc.ca
-
Handling editor: Britt Glaunsinger
Abstract
Flavivirus infections, including those of Dengue virus (DENV) and Zika virus (ZIKV), result in a high disease burden globally, yet many aspects of their viral life cycle remain poorly understood. For example, while some features of the mechanism of negative-strand RNA synthesis are known, relatively little is known about the initiation of positive-strand RNA synthesis in the flavivirus life cycle. Viral RNA replication is initiated via the recruitment of the viral NS5 RNA-dependent RNA polymerase (RdRp) to stem–loop A (SLA) at the 5′ terminus of positive-strand genomic RNA. Subsequent genome cyclization is thought to facilitate loading of NS5 onto the 3′ terminus of the genomic RNA to initiate negative-strand RNA synthesis. Conversely, it is not clear whether RNA structures in the negative-strand replicative intermediate similarly recruit NS5 to promote positive-strand RNA synthesis, providing specificity to this process. Herein, we characterized the secondary structure of the 3′ terminus of the negative-strand replicative intermediate in ZIKV and DENV1–4 in silico and in vitro. We observed that the 3′ terminus of the negative strand is capable of forming a secondary structure which mirrors SLA, which we term SLA′. While we demonstrate that SLA′ forms in vitro and is capable of interacting with NS5, introduction of G·U wobble base pairs that disrupt SLA′, while keeping SLA largely intact, suggest that SLA′ is not necessary for viral RNA replication. As such, this work suggests that in contrast to related viruses, the positive-strand promoter is unlikely to be provided by specific structure(s) at the 3′ terminus of the negative-strand replicative intermediate.
Keywords
- flavivirus
- Zika virus
- SHAPE
- stem–loop A (SLA)
- nonstructural protein 5 (NS5)
- negative-strand replicative intermediate
INTRODUCTION
Flaviviruses, positive-sense single-stranded RNA viruses from the genus Flavivirus, include several emerging pathogens with a large global disease burden. Their ranks include the mosquito-borne Dengue virus (DENV) and Zika virus (ZIKV), which together result in significant disease burden, ranging from mild febrile illness to severe dengue and neurological complications (Guillain–Barré and fetal microcephaly), respectively (Bhatt et al. 2013; Cauchemez et al. 2016). More than three billion people are at risk of flaviviral infection globally, with climate change–induced expansion in the range of their insect vectors threatening to exacerbate the issue (Iwamura et al. 2020; Pierson and Diamond 2020). With limited vaccines and antiviral therapies available for most flaviviruses, it is critical to increase our understanding of their life cycle to provide new targets for the development of antiviral interventions.
As positive-sense single-stranded RNA viruses, flaviviruses must undergo a multistep process to produce progeny positive-strand genomic RNAs during infection. After entry and uncoating, the capped positive-strand genomic RNA can be directly translated by host cell ribosomes to synthesize the viral proteins. Once enough viral proteins have accumulated, translation must be switched off since ribosomes travel in the 5′ to 3′ direction, while the viral nonstructural protein 5 (NS5), the RNA-dependent RNA polymerase (RdRp), needs to travel along the genome in the 3′ to 5′ direction (Barton et al. 1999; Sanford et al. 2019; Fajardo et al. 2020; Sagan and Weber 2023). The switch from translation to replication is thought to be mediated by recruitment of the NS5 protein to stem–loop A (SLA) at the 5′ terminus of the viral positive-strand genomic RNA (Filomatori et al. 2006; Fajardo et al. 2020; Abram et al. 2024). This leads to ribosome exclusion and genome cyclization, mediated through complementary sequences in the 5′ and 3′ termini of the genome (Alvarez et al. 2005; Villordo and Gamarnik 2009; Villordo et al. 2010; Sanford et al. 2019; Fajardo et al. 2020). Subsequently, NS5 can be loaded onto the 3′ terminus to initiate negative-strand RNA synthesis, resulting in a double-stranded RNA (dsRNA) intermediate (Filomatori et al. 2006, 2011; Abram et al. 2024). The negative strand of the dsRNA intermediate is then used as a template for the subsequent production of progeny positive-sense genomic RNAs. Finally, newly synthesized positive-sense genomic RNAs are capped and can either be shuttled into translation or packaged into progeny viral particles (Mazeaud et al. 2018; Barnard et al. 2021; Abram et al. 2024). Of note, while SLA at the 5′ terminus of the viral genome promotes negative-strand RNA synthesis, whether RNA structures in the negative strand similarly act as a promoter for positive-strand RNA synthesis, and what factors govern selection of the negative-strand template in the dsRNA replicative intermediate, remains unclear.
Interestingly, regions of the negative strand in the closely related hepatitis C virus (HCV) and other positive-sense RNA viruses can form local structures with low frequency (Herod et al. 2022). In particular, the 3′ terminus of the negative strand has been suggested to serve as the promoter for positive-strand RNA synthesis in HCV (Friebe and Bartenschlager 2009; Chahal et al. 2021). Furthermore, in the related flavivirus, Japanese encephalitis virus (JEV), the viral NS5 protein has a higher affinity for the 3′ end of the negative strand than the 3′ end of the positive-strand genomic RNA (Kim et al. 2007). Moreover, this region has also been implicated in efficient regulation of ZIKV positive-strand RNA synthesis in vitro (Calmels et al. 2022). As such, we investigated the structure of the 3′ terminus of the negative-strand RNAs of ZIKV and DENV in silico and in vitro. We demonstrate that the 3′ terminus of the negative strand of both ZIKV and DENV is capable of folding into a structure that mirrors SLA, which we term herein SLA′. Moreover, we show that SLA′ can interact with NS5 in a manner similar to SLA–NS5 interactions, albeit with reduced affinity. However, when we introduced G·U wobble base pairs into the ZIKV genome which disrupt SLA′, while leaving SLA largely intact, we found that SLA′ was not required for viral RNA accumulation in a ZIKV subgenomic replicon system. Thus, in contrast to what has been suggested in the related HCV, the specific structure of the 3′ terminus of the negative-strand replicative intermediate does not appear to function as the promoter for positive-strand RNA synthesis in flaviviruses.
RESULTS
The 3′ terminus of the negative strand of ZIKV and DENV is predicted to form SLA′
Previous studies demonstrate that SLA at the 5′ terminus of the positive-sense genomic RNA plays a central role in the flavivirus life cycle, regulating translational shut-off, initiation of negative-strand RNA synthesis, and genome capping (Fig. 1A; Filomatori et al. 2006; Dong et al. 2007; Fajardo et al. 2020; Abram et al. 2024). SLA is typically depicted as a “Y”-shaped structure in two-dimensional (2D) space, with a base stem, top stem–loop (TSL), and side-stem–loop (SSL) (Fig. 1A,B). However, recent structural studies suggest that SLA takes on more of a “V”- or “L”-shaped structure in three-dimensional (3D) space (Lee et al. 2021; Sun and Varani 2022; Osawa et al. 2023). Much less is known about the structure(s) of the complementary region at the 3′ terminus of the negative strand as well as their importance in the flaviviral life cycle.
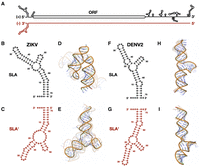
Zika virus (ZIKV) and Dengue virus 2 (DENV2) are predicted to form SLA′ at the 3′ terminus of their negative strands. (A) Cartoon diagram of the ZIKV positive-strand genomic RNA (black) and its complementary negative-strand intermediate (cayenne). Stem–loop A (SLA) is shown at the 5′ terminus of the positive strand, while the predicted SLA′ structure is shown at the 3′ terminus of the negative strand. Predicted secondary and tertiary structures of the first 70 nt of the 5′ terminus of the positive strand and the last 70 nt of the 3′ terminus of the negative strand of (B–E) ZIKV and (F–I) DENV2, respectively. Tick marks represent 10 nt intervals. The highest confidence tertiary structure prediction is fully opaque, while the next four highest-ranked predictions are translucent.
To explore the structural architecture of this region, we first performed secondary and tertiary structure predictions of the 5′ terminus of the positive strand and the 3′ terminus of the negative strand of ZIKV and DENV2 (Fig. 1B–I). Notably, prediction analyses in both cases revealed “Y”-shaped structures at the 3′ terminus of the negative strand that largely mirror SLA, which we termed SLA′ (Fig. 1B,C,F,G). In the case of ZIKV, two G·U wobble base pairs in SLA result in a larger central bulge in SLA′, but the architecture of the base stem, TSL, and SSL are largely maintained (Fig. 1B,C). In contrast, due to the absence of G·U wobble base pairs, the DENV2 SLA′ structure is a direct mirror image of SLA (Fig. 1F,G). Interestingly, the predicted Gibbs free energy of SLA′ in ZIKV was significantly higher than that of SLA, while the predicted Gibbs free energy of DENV2 SLA and SLA′ were quite similar, suggesting that the thermodynamic stability of these structures differs between the two viruses (Table 1).
Predicted Gibbs free energy of SLA and SLA′ in ZIKV and DENV
As recent structural studies have suggested that the tertiary structure of SLA resembles a “V”- or “L”-shaped structure, we also performed in silico tertiary structure predictions of SLA and SLA′ in ZIKV and DENV2 using AlphaFold 3 (Fig. 1D,E,H,I; Lee et al. 2021; Sun and Varani 2022; Osawa et al. 2023). While the predicted local distance difference test (pLDDT) scores at each nucleotide were predominantly low (0.5 < pLDDT < 0.7) indicating low prediction confidence, we found that the tertiary structure prediction of ZIKV SLA was a “V”-shaped structure, similar to what has been reported in X-ray crystallography studies (Lee et al. 2021). Interestingly, ZIKV SLA′ was predicted to take on a more open “L”-shaped conformation (Fig. 1D,E). This is likely due to G56·U16 wobble base-pairing in SLA, which corresponds to a loss of two base pairs at the top of the base stem of SLA′, resulting in a reduced rotation of the base stem helix before the central bulge (Fig. 1E). However, the respective SSL and TSL conformations are well aligned with the secondary structure predictions, with the unpaired “AG” motif in the unpaired loop of the TSL (Fig. 1E). In contrast, the tertiary structure predictions of DENV2 SLA and SLA′ are largely mirror images, albeit with a more open “L”- or “I”-shaped conformation than what has been observed in recent structural studies (Fig. 1H,I; Lee et al. 2021; Sun and Varani 2022; Osawa et al. 2023). Additionally, when we performed similar secondary and tertiary structural predictions for the 5′ terminus of the positive strand and 3′ terminus of the negative strand of DENV1, DENV3, and DENV4, we also observed SLA′ structures that resemble the more open “L”- or “I”-shaped conformation (Supplemental Fig. S1; Lee et al. 2021; Sun and Varani 2022; Osawa et al. 2023). In these three DENV serotypes, the base stem and TSL of SLA′ largely mirror that of their respective SLA, with greater divergence in the structure of the central bulge between the two stems. Interestingly, the predicted Gibbs free energy of SLA′ was similar to that of the corresponding SLA for DENV1 and DENV4, but for DENV3 was significantly higher in SLA than in SLA′ (Table 1). In addition, the DENV3 SLA′ had the lowest predicted Gibbs free energy, which correlates with the SLA of DENV3 containing the most G·U wobble base pairs (Table 1; Supplemental Fig. S1). Taken together, we find that for both ZIKV and all four DENV serotypes, the 3′ terminus of the negative-strand replicative intermediate is predicted to form SLA′, a structure which largely mirrors SLA in both secondary and tertiary structures.
In vitro SHAPE analyses are consistent with predictions for SLA′ in both ZIKV and DENV
We next sought to further validate the structural predictions using in vitro selective 2′ hydroxyl acylation analyzed by primer extension (SHAPE) analysis on the 5′ terminus of the positive-strand and 3′ terminus of the negative-strand viral RNAs, respectively (Figs. 2, 3). For both strands, we performed SHAPE analysis on the terminal 163 nt, a region on the positive strand that includes the complete 5′ untranslated region (UTR), encompassing both SLA and stem–loop B (SLB), as well as the capsid hairpin (cHP) structure, and all three viral cyclization sequences (including the 5′ upstream of AUG region, 5′ downstream from the AUG region, and cyclization sequence) (Supplemental Figs. S2, S3; Alvarez et al. 2005; Clyde and Harris 2006; Filomatori et al. 2006, 2011; Clyde et al. 2008; Lodeiro et al. 2009; Villordo and Gamarnik 2009; Villordo et al. 2010; Abram et al. 2024). While we focus herein on the first 70 nt of the viral genome which encodes SLA, our SHAPE-constrained structural predictions of these 163 nt RNAs were consistent with structural predictions and prior mutational analyses, demonstrating the robustness of our in vitro SHAPE approach (Figs. 2, 3; Supplemental Figs. S2, S3). Importantly, in vitro SHAPE analyses supported the formation of SLA′ in the 3′ terminus of the ZIKV and DENV2 negative strands, as well as two additional structures which we termed SLB′ and SLC′ (Figs. 2, 3; Supplemental Figs. S2, S3). However, due to the presence of G·U wobble base pairs, SLB′ and SLC′ do not have the same “mirrored” topology to their positive-strand RNA counterparts. Due to the importance of SLA in the flavivirus life cycle, and the mirrored topology we observed for SLA′, we focused our attention on these structures. Similar patterns of SHAPE reactivity were also observed for SLA and SLA′ in DENV1, 3, and 4 (Supplemental Figs. S4–S6). Taken together, our in vitro SHAPE analyses indicate that both SLA and SLA′ are able to form in vitro at the 5′ terminus of the positive-strand and the 3′ terminus of the negative-strand RNA, respectively, in ZIKV and DENV1-4.
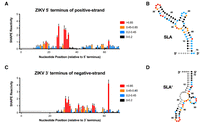
In vitro selective 2′ hydroxyl acylation analyzed by primer extension (SHAPE) analysis of the 3′ terminus of the negative strand in Zika virus (ZIKV) is consistent with SLA′ formation. (A) Normalized SHAPE reactivities of the first 70 nt at the 5′ terminus of the positive strand of ZIKV. Data are shown as the normalized SHAPE reactivity from four biological replicates, and error bars represent the SEM. Nucleotides with very low (≤0.2), low (0.2–0.4, blue), intermediate (0.4–0.85, orange), and high (≥0.85, red) SHAPE reactivity are indicated. Nucleotides 1–6 were omitted due to high background reactivity (light gray). (B) Prediction of the lowest free energy structure formed by the first 70 nt of the ZIKV positive strand as constrained by the normalized SHAPE reactivity data from (A). Tick marks represent 10 nt intervals. (C) Normalized SHAPE reactivities of the last 70 nt at the 3′ terminus of the negative strand of ZIKV. Data are shown as the normalized SHAPE reactivity from four biological replicates, and error bars represent the SEM. Nucleotides 1–15 were bound by the primer (dark gray), and nucleotides 16–23 and 157–163 were omitted due to high background reactivity (light gray). (D) Prediction of the lowest free energy structure formed by the last 70 nt of the ZIKV negative strand as constrained by the normalized SHAPE reactivity data from (C).
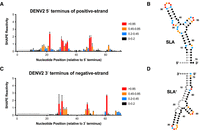
In vitro SHAPE analysis of the 3′ end of the negative strand in DENV2 is also consistent with SLA′ formation. (A) Normalized SHAPE reactivities of the first 70 nt at the 5′ terminus of the positive strand of DENV2. Data are shown as the normalized SHAPE reactivity from four biological replicates, and error bars represent the SEM. Nucleotides with very low (≤0.2), low (0.2–0.4, blue), intermediate (0.4–0.85, orange), and high (≥0.85, red) SHAPE reactivity are indicated. Nucleotides 1–7 were omitted due to high background reactivity (light gray). (B) Prediction of the lowest free energy structure formed by the first 70 nt of the DENV2 positive strand as constrained by the normalized SHAPE reactivity data from (A). Tick marks represent 10 nt intervals. (C) Normalized SHAPE reactivities of the last 70 nt at the 3′ terminus of the negative strand of DENV2. Data are shown as the normalized SHAPE reactivity from four biological replicates, and error bars represent the SEM. Nucleotides 1–15 were bound by the primer (dark gray), and nucleotides 16–24 and 157–163 were omitted due to high background reactivity (light gray). (D) Prediction of the lowest free energy structure formed by the last 70 nt of the DENV2 negative strand as constrained by the normalized SHAPE reactivity data from (C).
ZIKV NS5 can bind to SLA′, but with a lower affinity than SLA
As SLA′ is largely a mirror image of SLA in ZIKV and DENV, we wondered whether SLA′ was also capable of interacting with NS5, the viral RdRp. To test this hypothesis, we performed electrophoretic mobility shift assays (EMSAs) with full-length ZIKV NS5 and the 5′ terminus of the positive-strand and 3′ terminus of the negative-strand RNA, which contain SLA and SLA′, respectively (Fig. 4). Consistent with previous studies that demonstrated binding of the flavivirus NS5 protein to SLA, we observed a shift in mobility of the 5′ terminus of the positive strand (SLA) with increasing molar ratio of NS5, starting at a molar ratio of around 1:1 (Fig. 4A; Fajardo et al. 2020; Wang et al. 2020; Lee et al. 2021). Notably, since NS5 exists in monomeric, dimeric, and higher-ordered oligomeric states in solution, we observe multiple shifts in the EMSA, consistent with monomeric, dimeric, and higher-ordered NS5 complexes (Klema et al. 2016; Ferrero et al. 2019; Fajardo et al. 2020; Wang et al. 2020; Lee et al. 2021; Yang et al. 2021). Interestingly, we also observed a shift in the mobility of the 3′ terminus of the negative strand (SLA′) with increasing molar ratio of NS5, albeit to a lesser extent than for the 5′ terminus of the positive strand (Fig. 4B). Again, we observed multiple shifts consistent with monomeric, dimeric, and higher-ordered NS5 complexes. In addition, we used our EMSA data to generate binding affinity curves for NS5 interactions with SLA- or SLA′-containing RNAs, allowing us to estimate equilibrium dissociation constants (Kd) of Kd = 148.0 nM (140.9–155) and 252.7 nM (245.9–259.5), respectively (Fig. 4C,D). Thus, NS5 is able to interact with both SLA and SLA′, at least in vitro.
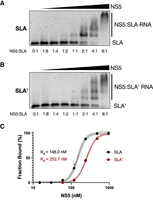
ZIKV NS5 binds to both SLA- and SLA′-containing RNAs. Electrophoretic mobility shift assays (EMSAs) were performed using ZIKV NS5 and (A) the first 163 nt of the ZIKV positive strand, or (B) the last 163 nt of the ZIKV negative strand. For each RNA, 2.5 pmol (150 ng) of RNA was incubated with increasing (twofold) amounts of NS5 ranging from 0 to 20 pmol (0–2 µg) and then analyzed via agarose gel electrophoresis. Molar ratios of NS5:RNA are indicated. Data shown are representative of four independent replicates. (C) Binding curves of NS5 to SLA- and SLA′-containing RNAs based on the bound RNA fraction from the EMSAs in A and B. Data for each RNA were fit to a sigmoidal four-parameter logistic curve model, with R2 values of R2 = 0.9962 and R2 = 0.9987 for SLA and SLA′, respectively. The dashed lines indicate 95% confidence intervals.
Since we observed differences in the binding affinity of NS5 for RNAs containing SLA and SLA′, we wondered if NS5 interacts with both RNAs in a similar manner. For SLA, the RdRp and methyltransferase (MTase) domains of NS5 are thought to interact with the TSL and base stem, respectively (Lee et al. 2021; Osawa et al. 2023). Binding is predicted to occur via a three-step process where the RdRp binds first to the TSL, followed by MTase binding to the base stem, before a final structural rearrangement to stabilize the NS5–SLA interaction (Bujalowski et al. 2020). Importantly, the NS5 RdRp domain is thought to interact with the “AG” motif in the unpaired loop of the TSL to provide specificity to the interaction (Tilgner et al. 2005; Dong et al. 2008; Filomatori et al. 2011; Hodge et al. 2016; Lee et al. 2021; Wang et al. 2022; Upstone et al. 2023). To determine how NS5 binding to SLA and SLA′ altered the secondary structure of the RNA, we performed in vitro SHAPE analysis at 0:1 and 2:1 molar ratios of NS5:RNA (Fig. 5; Supplemental Fig. S7). For the 5′ terminus of the positive strand, NS5 binding resulted in a substantial decrease in SHAPE reactivity broadly across SLA (Fig. 5A,B). In particular, we observed a large reduction in SHAPE reactivity in the loop region of the TSL, consistent with previous studies that suggest binding of this region by the RdRp domain of NS5 (Fig. 5B, inset; Tilgner et al. 2005; Dong et al. 2008; Filomatori et al. 2011; Hodge et al. 2016; Bujalowski et al. 2020; Lee et al. 2021; Wang et al. 2022; Upstone et al. 2023). We additionally observed an overall reduction in SHAPE reactivity across the SLB structure (Supplemental Fig. S8). In agreement with our EMSA results, we further observed an overall reduction in SHAPE reactivity for the 3′ terminus of the negative strand. However, the reduction was much more modest than for the 5′ terminus of the positive strand, consistent with NS5 having a lower affinity for SLA′ than for SLA (Fig. 5C,D; Supplemental Fig. S7). Nonetheless, we still observed a modest decrease in SHAPE reactivity in the loop region of the TSL of SLA′, perhaps implicating this region in NS5–SLA′ interactions (Fig. 5D, inset). In contrast, we did not observe a consistent decrease in SHAPE reactivity in the identified SLB′ and SLC′ structures that would be consistent with NS5 interacting with these structures (Supplemental Fig. S9). However, there were overall increases in SHAPE reactivity in the single-stranded regions at the 3′ and 5′ termini of the SLA- and SLA′-containing RNAs, respectively (Supplemental Figs. S8, S9). Taken together, our EMSA and SHAPE analyses suggest that NS5 is capable of binding both SLA and SLA′ in vitro; however, NS5 has a lower affinity for SLA′ than for SLA.

NS5 binding results in a change in SHAPE reactivity in the top loop of SLA and SLA′. ΔSHAPE reactivity plots for (A), the first 70 nt of the 5′ terminus of the positive strand and (C), the last 70 nt of the 3′ terminus of the negative-strand in the presence or absence of ZIKV NS5 (i.e., ΔSHAPE = NS5 + RNA reactivity − RNA only reactivity). Data are shown as the normalized change in SHAPE reactivity upon NS5 binding at each nucleotide position from four biological replicates, and error bars represent the SEM. Significant (>magnitude of the average ΔSHAPE reactivity) increases (red) and decreases (blue) in SHAPE reactivity upon NS5 binding are indicated. The baseline significance is indicated with a dashed line. Nucleotides 1–3 in A and 16–20 in C were omitted due to high background reactivity (light gray), while nucleotides 1–15 in C were bound by the primer (dark gray). (B,D) ΔSHAPE reactivity data from (A,C) mapped onto the predicted structures of (B) SLA and (D) SLA′. Tick marks represent 10 nt intervals. Insets represent the normalized SHAPE reactivities of the respective top loop regions in the absence and presence of NS5. Nucleotides with very low (≤0.2), low (0.2–0.4, blue), intermediate (0.4–0.85, orange), and high (≥0.85, red) normalized SHAPE reactivity are indicated in each inset.
SLA′ is not necessary for ZIKV RNA replication
While it was intriguing that SLA′ forms a structure that mirrors SLA and can interact with NS5 in vitro, it remained unclear whether SLA′ plays a specific role in the flaviviral life cycle. While this region of the genome has been suggested to serve as the promoter for positive-strand RNA synthesis in related viruses, we were curious whether the specific structure/conformation of SLA′ was important for ZIKV viral RNA accumulation in cell culture. However, due to the importance of the complementary SLA structure in viral RNA replication and genome capping, any mutations introduced to disrupt the SLA′ structure will concomitantly result in changes to SLA. To overcome this issue, we introduced G·U wobble base pairs into SLA in the place of existing A-U Watson–Crick base pairs, such that the overall SLA structure and thermodynamic stability would be largely maintained, while the conformation of SLA′ would be significantly disrupted (Fig. 6; Table 2). More specifically, we prepared three independent mutants, where mutations were introduced into the upper and lower portions of the TSL, TSL-U (G·U) and TSL-L (G·U), and to the base stem, BS (G·U). For the two TSL mutants, additional mutations were introduced to remove the possibility of alternative base-pairing interactions within SLA, but maintained the overall thermodynamic stability by preserving the number of G-C base pairs in each portion of the TSL. In all cases, the mutations resulted in a small reduction in the thermodynamic stability (Table 2), but preserved the predicted base-pairing and overall structure of SLA; while the 3′ terminus of the negative strand (i.e., SLA′) is predicted to have a significantly different overall topology (Fig. 6A,B,E). To verify that the introduced mutations largely preserved SLA while disrupting SLA′, we performed in vitro SHAPE analysis on the 5′ terminus of the positive strand and 3′ terminus of the negative strand of the G·U mutant RNAs (Supplemental Figs. S10, S11). For each of the G·U mutants, the predicted structure of SLA constrained by SHAPE reactivity differed slightly from WT ZIKV SLA, resembling the “V”- or “L”-shaped structures observed in recent tertiary structures of flavivirus SLA (Supplemental Fig. S10; Lee et al. 2021; Sun and Varani 2022; Osawa et al. 2023). Of note, the TSL-U (G·U) mutant displayed increased reactivity in the region adjacent to the top loop region of the TSL, in contrast with WT SLA and the other G·U mutant SLAs (Supplemental Fig. S10). Conversely, the in vitro SHAPE data verified that the structure of SLA′ was significantly disrupted for each of the G·U mutants, with the BS (G·U) mutant having the most drastic change in overall structure (Fig. 6E; Supplemental Fig. S11). To test the importance of SLA′ for viral RNA accumulation, each of these mutants were introduced into the context of a ZIKV Renilla luciferase (RLuc) reporter subgenomic replicon RNA (Fig. 6C). Wild-type (WT) and mutant subgenomic RNAs were electroporated into BHK-21 cells, and viral RNA accumulation was monitored by luciferase activity (Fig. 6D). Interestingly, while each of the mutants were impaired with respect to the WT subgenomic replicon, both the TSL-L (G·U) and BS (G·U) mutants were able to recover to WT levels by 72 h post-electroporation (Fig. 6D). In contrast, the TSL-U (G·U) mutant behaved similarly to the replication-defective subgenomic replicon, containing inactivating mutations in the NS5 RdRp active site (GNN), which served as a negative control (Fig. 6D). Importantly, there was no clear correlation between replicative capacity and the predicted thermodynamic stability of either SLA or the disrupted SLA′ structure (Fig. 6D; Table 2). Given that the TSL-L (G·U) and BS (G·U) mutants result in significant disruption of the SLA′ conformation (Fig. 6E; Supplemental Figs. S10, S11), these results indicate that the SLA′ structure is not necessary for ZIKV RNA replication. As such, the strong conservation of SLA′ is more likely related to the function of the complementary SLA structure in the positive-strand genomic RNA.
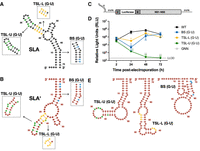
SLA′ is not necessary for ZIKV RNA replication. Cartoon diagrams of WT (A) SLA and (B) SLA′ with insets for the three sets of mutations that introduce G·U wobble base pairs into the lower and upper portion of the TSL (TSL-L [G·U], yellow and TSL-U [G·U], green), and base stem (BS [G·U], blue). (C) Schematic representation of the ZIKV subgenomic replicon system, which contains the Renilla luciferase (RLuc) gene in place of the majority of the ZIKV structural proteins. (D) Full-length capped WT, TSL-L (G·U), TSL-U (G·U), or BS (G·U) ZIKV subgenomic RLuc reporter replicon RNAs were electroporated into BHK-21 cells. Luciferase activity was measured at 2–72 h post-electroporation. The lower limit of detection is indicated (LLOD). Data are representative of three independent biological replicates with three technical replicates, and error bars represent standard deviation (SD) of the mean. Statistical significance was determined by multiple Student's t-tests, (*) P ≤ 0.1, (**) P ≤ 0.05. (E) Most stable predicted secondary structure of the last 70 nt at the 3′ terminus of the negative strand for the TSL-U (G·U), TSL-L (G·U), and BS (G·U) mutants, as constrained by normalized SHAPE reactivity data.
Predicted Gibbs free energy of SLA and SLA′ in ZIKV G·U mutants
DISCUSSION
Herein, we explored the RNA secondary and tertiary structures of the 3′ terminus of the negative strand in both ZIKV and DENV1-4. Secondary and tertiary structure predictions identified SLA′, which is largely a mirror image of SLA, the terminal structure at the 5′ terminus of the positive-strand genomic RNA. This was somewhat surprising, because the presence of G·U wobble base pairs often means that complementary RNAs do not possess a mirrored structural architecture. This is exemplified by the 3′ terminus of the HCV negative strand, which forms several structures that do not resemble those in the 5′ terminus of the positive-strand genomic RNA (Friebe and Bartenschlager 2009). Thus, it was intriguing that in both ZIKV and DENV1–4, SLA′ was predicted to have a similar mirrored architecture to SLA, the promoter for negative-strand RNA synthesis. Subsequently, we were able to confirm that these structures were able to form in vitro in the 3′ terminus of both ZIKV and DENV1–4 RNAs, and that both ZIKV SLA and SLA′ could interact with NS5, albeit with differing affinities.
Our EMSA and SHAPE analyses provide support for NS5 binding to both SLA and SLA′-containing RNAs in vitro and suggest that this interaction may also be mediated through interactions with the top loop region of the TSL, as previously suggested for NS5–SLA interactions (Tilgner et al. 2005; Dong et al. 2008; Filomatori et al. 2011; Hodge et al. 2016; Lee et al. 2021; Wang et al. 2022; Upstone et al. 2023). While the “AG” motifs in the top loops of SLA and the 3′SL are highly conserved across flaviviruses, the A in this motif is not a strict requirement for NS5 binding. Indeed, DENV2 mutants with “CG” and “UG” top loop motifs in the 3′SL are readily bound by NS5, while DENV2 with a “GG” top loop motif in SLA is also replication-competent (Filomatori et al. 2006; Hodge et al. 2016). As such, the “UG” motif in the top loop of SLA′ could theoretically allow for NS5 binding to this structure as well (Tilgner et al. 2005; Dong et al. 2008; Filomatori et al. 2011; Hodge et al. 2016; Lee et al. 2021; Wang et al. 2022; Upstone et al. 2023). On the other hand, while we also observed an overall decrease in SHAPE reactivity across SLB as expected, this was not the case for the identified SLB′ and SLC′ structures at the 3′ terminus of the negative strand. As such, in contrast to the role SLB plays in stabilizing the NS5–SLA interaction, it does not appear that SLB′ or SLC′ play an equivalent part in facilitating NS5 binding to SLA′ (Liu et al. 2016).
Further to this point, there is likely a fair degree of plasticity in the NS5–SLA and NS5–SLA′ interactions, since NS5s of different flaviviruses are often interchangeable with respect to SLA binding even in spite of the observed differences in predicted SLA structures between ZIKV and the different DENV serotypes (Yu et al. 2008; Bujalowski et al. 2017). Based on the similar thermodynamic stability across the predicted SLA and SLA′ structures, it is thus possible that both structures could form in a manner that supports NS5 binding (Table 1). However, given that the negative strand primarily exists as part of a dsRNA replicative intermediate during infection, it remains to be seen whether the environment in the replication organelle or during some step in the process of initiation of positive-strand RNA synthesis could favor the formation of SLA′ (Sagan and Weber 2023; Dahmane et al. 2024). To this end, a recent study suggests that local structures may indeed be able to form in the negative strand during infection, as ribozymes encoded in the negative strand of a variety of positive-strand RNA viruses were shown to be active during replication in cell culture (Herod et al. 2022). Nonetheless, as these events appear to occur at relatively low frequency, whether formation of SLA′ at the 3′ terminus of the negative strand would be stable long enough to both recruit and remain stably bound to NS5 remains an open question (Herod et al. 2022).
Finally, we sought to assess the functional relevance of SLA′ and understand whether this structure participates in positive-strand RNA synthesis in the context of viral RNA replication. Interestingly, introduction of G·U wobble base pairs into both the TSL and base stem of SLA, which retained the SLA conformation, but disrupted SLA′, resulted in only modest delays in viral RNA accumulation in two of the three mutants tested. It is likely that the inability of the TSL-U (G·U) mutant to replicate is related to the reduced thermodynamic stability of the upper portion of the TSL of SLA (ΔG = −7.8 vs. −8.2) (Supplemental Fig. S10), which may impair SLA interactions with the RdRp domain of NS5. Nevertheless, these findings suggest that SLA′ is not necessary for viral RNA accumulation in cell culture. Furthermore, since there was no correlation between the predicted stability of the 3′ terminal structure in these mutants and their ability to replicate, ZIKV RNA accumulation does not seem to require a stable structure at the 3′ terminus of the negative strand. Given these findings, it remains unclear how ZIKV and other flaviviruses select the negative strand from the dsRNA replicative intermediate as a template for progeny positive-strand genomic RNA synthesis. It is possible that proximity of the 3′ terminus upon completion of the negative-strand RNA in the dsRNA intermediate is sufficient to select the template or that end-breathing results in re-formation of SLA at the 5′ terminus of the positive strand, resulting in a freeing of the 3′ terminus of the negative strand for template selection. Alternatively, ZIKV could use a mechanism more akin to that of polioviruses, where the 5′ terminal structure of the positive strand acts as the promoter for both negative-strand and positive-strand RNA synthesis, acting in trans to initiate the latter process (Vogt and Andino 2010). However, more research will be needed to determine the mechanism of template strand selection during flavivirus RNA replication.
MATERIALS AND METHODS
Maintenance of cell lines
Baby hamster kidney-21 (BHK-21) cells were maintained in vented, tissue-culture treated flasks (Sarstedt) in complete media (Dulbecco's minimal essential medium [DMEM] supplemented with 10% fetal bovine serum [Wisent], 1× MEM nonessential amino acids [Corning], and 2 mM l-glutamine [Gibco]). Cells were cultured at 37°C and 5% CO2 with regularly screening for mycoplasma contamination.
Cloning
To generate the ZIKV NS5-His expression plasmid for production of full-length ZIKV NS5, the pET22b plasmid was first modified by reverse PCR with Q5 Hot Start High-Fidelity 2X Master Mix (New England BioLabs) using the pET22b GSSS RPCR Fwd and pET22b GSSS RPCR Rev primers (Supplemental Table S1) to add a GSSS linker sequence to the N terminus of the encoded 6X His-tag. Next, the NS5 coding sequence from the ZIKV PRVABC59 strain (GenBank accession: KU501215.1) was codon optimized for BL21 Escherichia coli using the IDT Codon Optimization Tool to generate a geneblock (IDT, Supplemental Table S2). The codon-optimized NS5 sequence was inserted between the start codon and the GSSS linker of the modified pET22b expression plasmid via Gibson assembly using the ZIKV CO NS5 and pET22b CO RPCR primer sets (Supplemental Table S1). The resulting plasmid, pET22b ZIKV NS5-His, encodes the codon-optimized ZIKV full-length NS5 fused to a 6X His-tag via a GSSS linker. The plasmid was prepared using the QIAprep Spin Miniprep kit (QIAGEN), according to the manufacturer's instructions and verified via Sanger sequencing (Sequencing + Bioinformatics Consortium) and Oxford Nanopore sequencing (Plasmidsaurus).
To produce the IVT template plasmids, geneblocks (IDT) were generated for the 5′ end of the positive strand for DENV1 (GenBank accession: NC_001477.1), DENV2 (GenBank accession: NC_001474.2), DENV3 (GenBank accession: NC_001475.2), and DENV4 (GenBank accession: NC_002640.1), and for the 3′ end of the negative strand of ZIKV and DENV2 (Supplemental Table S2). The geneblocks for the 5′ end of the positive strand of DENV1–4 consisted of an in vitro transcription cassette in which the T7 promoter controls transcription of the first 359 nt of the respective reference genome followed by nt 360–383 of the ZIKV PRVABC59 genome (complementary to the ZIKV [+] 5′-UTR PE oligo) and the hepatitis delta virus ribozyme (HDVr). This transcription cassette was flanked by EcoRI and BamHI cut sites to facilitate cloning into the pUC18 backbone. For the 3′ end of the negative strand of ZIKV and DENV2, the region between the T7 promoter and the HDVr in the transcription cassette was replaced with the respective terminal 163 nt of the 3′ end of the negative strand. In all the above cases, the geneblock was amplified with EcoRI-T7 and BamHI-HDVr primers and then inserted into pUC18 using EcoRI and BamHI (Supplemental Table S1). The IVT template plasmid for the ZIKV 5′ end of the positive strand was generated by replacing the region in the ZIKV 3′ negative-strand IVT template plasmid between the T7 promoter and the HDVr ribozyme with the first 383 nt of the ZIKV PRVABC59 genome. This replacement was performed via Gibson assembly using the ZIKV 5′ (+) and T7-HDVr RPCR primer sets (Supplemental Table S1). A similar replacement was performed to generate the 3′ end of the negative-strand IVT template plasmid for DENV1, DENV3, and DENV4, where the respective terminal 163 nt of the 3′ end of the negative strand was inserted between T7 and HDVr via Gibson assembly using the T7-HDVr RPCR primer set and either the DENV1 3(−) Fwd and DENV1 3(−) Rev, DENV3 3(−) Fwd and DENV1 3(−) Rev, or DENV4 3(−) Fwd and DENV4 3(−) Rev primers, respectively (Supplemental Table S1). Plasmids were purified with the QIAprep Spin Miniprep kit (QIAGEN), according to the manufacturer's instructions and verified via Sanger sequencing (Sequencing + Bioinformatics Consortium).
Mutagenesis of SLA to disrupt the SLA′ structure via the introduction of G·U wobble base pairs was performed using the ZIKV PRVABC59 subgenomic Renilla luciferase (RLuc) reporter replicon system (Barnard et al. 2023). In brief, the replicon contains the ZIKV 5′ UTR, the first 38 amino acids of the capsid coding region fused to RLuc, the FMDV2A peptide, and the last 30 amino acids of the Envelope coding region through to the 3′ terminus of the ZIKV genome. To introduce the desired mutations into SLA, the region from PacI to ApaLI in the subgenomic replicon was subcloned into pUC18 via restriction digest and ligation with T4 DNA ligase (New England BioLabs). Mutations that introduce G·U wobble base pairs into the base stem, lower, and upper portions of the TSL, were added via reverse PCR with the SLA GU Base RPCR, SLA GU TSL Base RPCR, or SLA GU TSL Top RPCR primer sets, respectively (Supplemental Table S1). The subcloned region was amplified with the 5′ Rep SC GA primer set and reassembled with the PacI to ApaLI replicon via Gibson assembly (Table 1). Resulting replicon-containing bacterial artificial chromosomes were purified using the QIAGEN Large-Construct kit (QIAGEN), and sequences were verified by Oxford Nanopore sequencing (Plasmidsaurus).
BL21-pRARE competent cell preparation
BL21(DE3)-R3-pRARE2 was a gift from Opher Gileadi (Addgene plasmid # 26242) (Savitsky et al. 2010). These bacteria were used to inoculate a 5 mL culture of LB with 10 µg/mL chloramphenicol and incubated overnight at 37°C at 200 rpm. Next, 1 mL of the overnight culture was used to inoculate 99 mL of LB media with 10 µg/mL chloramphenicol and incubated at 37°C at 200 rpm until the culture reached an OD600 of 0.3–0.5. Bacteria were pelleted at 4500 rpm for 15 min at 4°C and then gently resuspended in 20 mL of ice-cold 1 m CaCl2. The resuspended cells were allowed to chill on ice for 15 min prior to centrifugation at 4500 rpm for 15 min at 4°C. Pelleted bacteria were then gently resuspended in 5 mL of ice-cold 0.1 m CaCl2, supplemented with 15% glycerol, and filter sterilized with a 0.22 µm filter (Millex). Aliquots of the competent cells were transferred to sterile, ice-cold microcentrifuge tubes and stored at −70°C for up to 1 year.
ZIKV NS5 production and purification
Competent BL21(DE3)-R3-pRARE2 cells were transformed with pET22b ZIKV NS5-His via a 30 sec heat shock at 42°C followed by 5 min on ice and a 45 min recovery period at 37°C and 300 rpm. Cells were plated on a LB plate containing 100 µg/mL ampicillin and 10 µg/mL chloramphenicol and incubated overnight at 37°C at 200 rpm. Multiple colonies were then used to inoculate 100 mL of LB medium containing 100 µg/mL ampicillin and 10 µg/mL chloramphenicol, and incubated overnight at 37°C at 200 rpm. Next, 10 mL of the starter culture was used to inoculate 1 L of LB medium containing 100 µg/mL ampicillin and 10 µg/mL chloramphenicol, which was incubated using the above conditions until the culture reached an OD600 of 0.6–0.8. At this point, the culture was cold shocked twice in an ice bath for 2 min with swirling before addition of isopropyl β-d-1-galactopyranoside to a final concentration of 0.5 mM. Induced cultures were incubated at 16°C at 230 rpm for 18 h and then pelleted at 4500 rpm for 15 min at 4°C. Pelleted cells were washed once with 20 mL of ice-cold phosphate buffered saline (PBS) and frozen at −70°C.
Frozen pellets were thawed on ice and resuspended in lysis buffer (20 mM Tris-HCl pH 7.5, 500 mM NaCl, 1.4 mM β-mercaptoethanol, 10% glycerol, 30 mM imidazole) with protease inhibitors (1 mM PMSF, 1 mM benzamidine, 0.7 µg/mL pepstatin A, 5 µg/mL leupeptin). Resuspended pellets were lysed via 4 × 1 min sonication pulses at 18% amplitude, and the resulting lysate was centrifuged at 18,000 rpm for 50 min at 4°C. Supernatants were then loaded onto a 5 mL HisTrap FF column (Cytiva) for initial purification via nickel affinity chromatography. The column bound ZIKV NS5-His was washed with 10 column volumes of lysis buffer and eluted with nickel affinity elution buffer (20 mM Tris-HCl pH 7.5, 500 mM NaCl, 1.4 mM β-mercaptoethanol, 10% glycerol, 300 mM imidazole). Fractions containing ZIKV NS5-His were then pooled, diluted in cation affinity loading buffer (20 mM Tris-HCl pH 7.5, 1 mM DTT, 10% glycerol) to reduce the salt concentration to 100 mM, and filtered through a 0.45 µM filter (Millex) to remove any resulting precipitate. The desalted filtrate was then loaded onto a 5 mL SP HiTrap column (Cytiva) for cation affinity chromatography and washed with 10 column volumes of cation affinity washing buffer (20 mM Tris-HCl pH 7.5, 100 mM NaCl, 1 mM DTT, 10% glycerol). ZIKV NS5-His was eluted with a linear gradient of NaCl to a final concentration of 500 mM NaCl. Fractions containing ZIKV NS5-His were then pooled and concentrated before a final size exclusion chromatography purification on a Superdex 200 Increase 10/300 column (Cytiva), where buffer was exchanged for S200 running buffer (50 mM Tris-HCl pH 7.5, 500 mM NaCl, 5 mM DTT, 10% glycerol) under RNase-free conditions. The resulting fractions containing the purified ZIKV NS5-His were pooled and concentrated to ∼2 mg/mL using 50 kDa MWCO centrifugal filters (Amicon), and then flash frozen and stored at −70°C.
RNA secondary and tertiary structure prediction
Prediction of RNA secondary structure and Gibbs free energy were performed using the RNAstructure software package (available at https://rna.urmc.rochester.edu/index.html) (Reuter and Mathews 2010). In general, RNA sequences of interest were loaded into the RNAstructure software using the “Fold RNA Single Strand” command. To generate RNA structure predictions using in vitro SHAPE data as a constraint, corresponding RNA sequences were loaded as a “Single Sequence” using the “Structure Prediction” command. Normalized SHAPE data were then input as a constraint on secondary structure prediction as a .shape file. Outputs were saved as dot bracket files and visualized via VARNA (Darty et al. 2009).
Prediction of RNA tertiary structure was performed using AlphaFold Server (available at https://alphafoldserver.com/) (Abramson et al. 2024). The first 70 nucleotides (nt) of the positive strand and the last 70 nt of the negative strand were used as templates for predictions of SLA and SLA′, respectively. The five predicted structures with the highest AlphaFold ranking score were aligned and visualized in PyMol (Schrodinger and DeLano 2020, http://www.pymol.org/pymol.).
In vitro transcription of genome termini RNA
For in vitro SHAPE and EMSA experiments, DNA templates were generated by PCR and in vitro transcribed into RNA with the T7 RiboMAX Kit (Promega), according to the manufacturer's instructions. DNA templates corresponding to the first 383 nt of the ZIKV and DENV1–4 genomes were generated from their respective IVT template plasmids using the EcoRI-T7 and BamHI-HDVr primers (Supplemental Table S1). To generate DNA templates corresponding to the first 163 nt of the ZIKV PRVABC59 genome, EcoRI-T7 and ZIKV + IVT 163 nt Rev (5′-CGTTTTAGCATATTGACAATCCGGAATC-3′) primers were used to amplify the region from the IVT template plasmids (Supplemental Table S1). Finally, amplification of DNA templates arising from the 3′ end of the negative strand of either ZIKV or DENV was performed using the EcoRI-T7 and BamHI-HDVr primers (Supplemental Table S1). All DNA templates were size-verified on a 2% agarose gel and purified using the QIAQuick PCR Purification Kit (QIAGEN), according to the manufacturer's instructions. In vitro transcribed RNA generated using the T7 RiboMAX Kit was treated with DNase I (NEB) for 20 min at 37°C, and then ethanol precipitated and purified. RNA was then gel purified on an 8% Urea-PAGE gel cast using the SequaGel UreaGel System (National Diagnostics) and run at 30 W for 2.5 h. Excised bands were incubated in 400 μL elution buffer (500 mM NH4OAc, 10 mM EDTA, and 0.1% sodium dodecylsulfate) overnight at 4°C with gentle agitation to extract the RNA. Gel-purified RNA was ethanol precipitated and stored at −20°C until use.
Selective 2′ hydroxyl acylation analyzed by prime extension (SHAPE)
In vitro SHAPE analysis was performed in quadruplicate as previously described (Chahal et al. 2019). In brief, 5 pmol of in vitro transcribed RNA was refolded and incubated in SHAPE buffer (333 mM HEPES pH 8.0, 20 mM MgCl2, 333 mM NaCl) at 37°C for 10 min. RNA was then treated with 0.01 m NAI-N3 or dimethyl sulfoxide (DMSO; treatment control) at 37°C for 5 min or 15 min for 383 and 163 nt IVT RNA, respectively. Treated RNA was then extracted using TRIzol reagent (Thermo Fisher), according to the manufacturer's instructions. Primer extensions (PEs) were performed in a 20 µL reaction volume containing 5 pmol of NAI-N3-labeled ZIKV RNA and 1 pmol of the 6-FAM-labeled PE oligo. ZIKV (+) 5′-UTR 6-FAM (5′-CAG CAT GGC AGC CAG ATC TTT CTT-3′) was used as the PE oligo for the 383 nt IVT RNAs corresponding to the 5′ ends of both the ZIKV and DENV positive strands, while the ZIKV (−) 3′ PE 6FAM (5′-AGT TGT TGA TCT GTG-3′) and DENV (−) 3′ PE 6FAM (5′-AGT TGT TAG TCT ACG-3′) oligos were used for PE of the 163 nt IVT RNA corresponding to the 3′ end of the negative strands of ZIKV and DENV, respectively. Matched sequencing ladders were generated for all reactions by performing primer extension reactions using 5 pmol of the respective untreated IVT RNA, 1 pmol of the respective NED-labeled PE oligo (ZIKV [+] 5′-UTR PE NED, ZIKV [−] 3′ PE NED, or DENV [−] 3′ PE NED), and either 0.5 mM ddCTP or ddGTP. Primer extensions were performed for 30 min at 52°C for the ZIKV (+) 5′-UTR PE reactions, and 45 min at 52°C for the ZIKV (−) 3′ PE and DENV (−) 3′ PE reactions. Following extension, 2 m NaOH was added and incubated at 95°C for 5 min to degrade the input RNA. The remaining cDNA was then diluted to a total volume of 100 µL with water and ethanol precipitated with 2.5 volumes 95% ethanol and 0.1 volumes 3 m sodium acetate pH 5.2 for storage at −70°C. Pelleted cDNA was washed three times with cold 85% ethanol and then dried at 37°C for 10 min. Dried pellets were sent for capillary electrophoresis on an ABI 3100 Genetic Analyzer at Plate-forme d'Analyses Génomiques de l'Université Laval. Raw fluorescence data were analyzed using the QuShape software package to generate normalized SHAPE reactivities for each nucleotide (Karabiber et al. 2013).
SHAPE analysis of the ZIKV IVT RNAs in the presence of recombinant NS5 protein was performed as above with the following exceptions for the NAI-N3-labeling step. Following refolding, the 5 pmol of RNA was added to a binding reaction with a total reaction volume of 20 µL containing either 1 ng of recombinant ZIKV NS5 (2 nNS5:1 nRNA) in NS5 binding buffer (20 mM Tris pH 7.4, 150 mM NaCl, 2 mM MgCl2, 10% glycerol) or NS5 binding buffer alone (0 nNS5:1 nRNA). Binding reactions proceeded at 25°C for 30 min, before treatment with 0.01 m NAI-N3 or DMSO (treatment control) at 25°C. The Wilcoxon rank test was done between the SHAPE data of RNA with NS5 and that of RNA alone to determine if there was a statistical difference between the SHAPE reactivities. The Wilcoxon test determined there are highly significant differences in SHAPE reactivity between groups for both SLA and SLA′ (P < 0.0001 for SLA; P < 0.05 for SLA). The baseline value for ΔSHAPE (NS5:RNA reactivity−RNA reactivity) was determined by finding the average of the absolute ΔSHAPE values. ΔSHAPE values above the positive baseline were considered significant increases and below the negative baseline value were considered significant decreases in SHAPE reactivity.
Electrophoretic mobility shift assay (EMSA)
First, 2.5 pmol (150 ng) of IVT RNA corresponding to the 163 nt at the 5′ terminus of the positive strand or the 3′ end of the negative strand was heated at 95°C and then snap-cooled on ice. Once cooled, RNA was added to a binding reaction with a total reaction volume of 20 µL containing one dilution of a series of ZIKV NS5-His ranging from a molar ratio (NS5:RNA) of 0–8:1 (0–2 µg of NS5) in NS5 binding buffer. Binding reactions proceeded at 25°C for 30 min before separation on a 1% agarose 0.5× TBE gel run for 1 h at 100 V on ice at 4°C. Gels were stained with 1× SYBR Gold nucleic acid gel stain (Thermo Fisher) on an orbital shaker at 120 rpm for 20 min and visualized on a ChemiDoc MP Imaging System (BioRad).
In vitro transcription of ZIKV replicon RNA
To generate full-length replicon RNAs, templates were linearized with PsrI (Creative Enzymes) and in vitro transcribed with SP6 RNA polymerase. In vitro transcription was performed in a total reaction volume of 60 µL containing 1 mM of each rNTP (New England BioLabs), 0.8 mM m7G(5′)ppp(5′)A Cap Structure Analog (New England BioLabs), 1 U/µL RiboLock RNase inhibitor (Thermo Fisher), 50 mM dithiothreitol (Thermo Fisher), and 1 U/µL SP6 RNA polymerase (New England BioLabs). Reactions proceeded at 37°C for 90 min, followed by a 20 min DNase I (New England BioLabs) treatment at 37°C. Capped firefly luciferase (FLuc) mRNAs were generated as previously described (Rheault et al. 2023). IVTs were then ethanol precipitated as described above and stored at −70°C until use.
Electroporations
BHK-21 cells were trypsinized, washed two times in 25 mL cold PBS, and then resuspended in cold PBS to a concentration of 1 × 107 cells/mL. In an ice-cold 4 mm cuvette, 400 µL of the above cell suspension (4 × 106 cells) was mixed with 10 µg of the respective ZIKV replicon RNA and 1 µg capped Fluc mRNA. Cells were then electroporated with one pulse of 270 V, 975 µF, and infinite resistance on a Bio-Rad Gene Pulser XCell (BioRad). Electroporated cells were then resuspended to a total volume of 8 mL in complete media. Two milliliters of cells were either placed in a 15 mL conical tube (2 h time point) or were plated in 6-well plates (24–72 h time points) and incubated at 37°C and 5% CO2 until harvest.
Luciferase assay
To collect samples for luciferase assays, electroporated cells were washed with 1 mL PBS, harvested in 100 µL 1× passive lysis buffer (Promega), and stored at 70°C until use. Luciferase assay was performed using the Dual Luciferase Assay kit (Promega), according to the manufacturer's instructions, except that 50 µL of each reagent was used for 10 µL of harvested sample in opaque, white 96-well plates (Greiner Bio-One). Luminescence was measured on a GloMax Explorer plate reader (Promega) with a 10 sec integration time. Luciferase activity of each sample was analyzed in duplicate, and all data are representative of three independent biological replicates.
Data and statistical analyses
Statistical analyses were performed in GraphPad 10 software. For analysis of luciferase assays, multiple unpaired Student's t-tests were performed for each time point in comparison with WT.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
We thank Young-Min Lee (Utah State University) for providing ZIKV PRVABC59 infectious cDNA that was used to generate our ZIKV luciferase replicon system. In addition, we thank John Pezacki (University of Ottawa) for assistance with the synthesis of NAI-N3 and Jasmin Chahal (McGill University) for assistance with SHAPE experimental setup and data analysis. We thank Hannah Hauch, Nathan Pilkey, and Aaron Tieu from the University of British Columbia for assistance with cloning aspects of the project. Q.H.A. thanks the Frederick Banting and Charles Best Canada Graduate Scholarships—Doctoral Award (CGS-D) and the Tomlinson Doctoral Scholarship for graduate support. Finally, we are grateful to current and past Sagan Lab members for their insightful discussions regarding this project. This research was supported by funds from the NSERC Discovery Grant (RGPIN-2020-04713) and Canadian Institutes of Health Research (PJT-162212) Operating Grant Programs to S.M.S. In addition, this research was undertaken, in part, thanks to funding from the Canada Research Chairs program.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080342.124.
-
Freely available online through the RNA Open Access option.
- Received December 1, 2024.
- Accepted April 18, 2025.
This article, published in RNA, is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Quinn H. Abram is the first author of this paper, “Structural and functional characterization of the SLA′ structure at the 3′ terminus of the Zika virus negative-strand intermediate.” Quinn is a PhD candidate at McGill University in the lab of Dr. Selena Sagan, which is now at the University of British Columbia. The main focus of Quinn's research is understanding the RNA–protein and protein–protein interactions that underlie the genome replication process in Zika virus and other flaviviruses.
What are the major results described in your paper and how do they impact this branch of the field?
Through this project, we were able to identify and confirm SLA′ as the RNA structure at the 3′ end of the negative-strand intermediate in Zika virus (ZIKV) and Dengue virus. We further confirmed that SLA′ could be bound by NS5, the viral RNA-dependent RNA polymerase, similar to the complementary SLA structure on the positive-sense genomic RNA. However, SLA′ turned out not to be necessary for genome replication in ZIKV. Thus, there must be alternative mechanisms for initiation of positive-strand synthesis in positive-sense single-stranded RNA viruses beyond the use of cis-acting RNA structural elements.
What led you to study RNA or this aspect of RNA science?
Positive-sense single-stranded RNA viruses such as ZIKV are fascinating models for RNA structure. Within the constraints of 11 kb of RNA for ZIKV, this virus needs to be able to enter cells, replicate its genome, and then form new virions to infect neighboring cells, all while evading and manipulating the cellular immune response. As such, the ZIKV genome is filled with functional and dynamic RNA structures that help facilitate all of these ends, and we are still scratching the surface of understanding them.
During the course of these experiments, were there any surprising results or particular difficulties that altered your thinking and subsequent focus?
After demonstrating that SLA′ was the mirror image of SLA and could also interact with NS5, we were certainly surprised when SLA′ turned out to not be necessary for positive-strand synthesis in ZIKV. We were especially surprised considering that other positive-sense single-stranded RNA viruses have been demonstrated to utilize cis-acting RNA structural elements to this end. As such, that result certainly opened our eyes to other possibilities for how ZIKV positive-strand synthesis is initiated, which we will hopefully be able to illuminate further moving forward.
What are some of the landmark moments that provoked your interest in science or your development as a scientist?
My scientific journey started at 7 years old, when I sought to confirm my father was masquerading as Santa Claus. However, my shoddy experimental design resulted in a false negative, demonstrating the importance of proper controls. More recently, I've been inspired by some of the recent work in the areas of single-molecule tracking and synthetic biology, which have allowed us to ask questions such as “what is the processing rate of a translating ribosome?” I think it is important to think about the dynamics occurring within a cell, and generate holistic models that both guide and provide the context for the work we do in the lab.
If you were able to give one piece of advice to your younger self, what would that be?
Always have a big-picture model to contextualize your research, but listen when the data are telling you to change it. Having a model to explain what you are studying is really helpful to both ground and guide your research. The flip side of that coin is that you can waste time going down the wrong path if you are too rigid and inflexible regarding your beliefs. So when your data doesn't fit with the overarching model, it is best to consider changing the model based on what you now know, rather than contorting yourself to force the data to conform with what you once thought to be true.








