
Repression of AGO1 by AGO2 via let-7 microRNAs facilitates embryonic stem cell differentiation
- Corresponding author: hu.wenqian{at}mayo.edu
-
Handling editor: Javier Caceres
Abstract
Argonaute (AGO) proteins are critical regulators of gene expression. Of the four AGOs in mammals, AGO1 and AGO2 are expressed in mouse embryonic stem cells (mESCs). These two proteins have opposing functions in controlling mESCs’ fate decisions between pluripotency and differentiation. AGO2 promotes differentiation predominantly via the let-7 microRNAs, whereas AGO1 maintains pluripotency via modulating protein folding independent of small RNAs. These recent findings raise the question of whether and how these two AGOs are mutually regulated in mESCs. Here, using loss-of-function and gain-of-function approaches, we show that AGO2 represses the expression of AGO1 mRNA via a conserved let-7-microRNA-binding site in its 3′ UTR. Mutating this binding site at the endogenous locus abolishes the AGO2-mediated repression of AGO1 mRNA and compromises the exit pluripotency of mESCs. These results indicate that the posttranscriptional regulation of AGO1 by AGO2 and let-7 microRNAs is important for stem cell differentiation, but also reveal a regulatory mechanism between the two AGO paralogs with opposing functions in controlling stem cell fate decisions.
Keywords
INTRODUCTION
Argonautes (AGOs) are evolutionarily conserved proteins with critical roles in the microRNA mediated gene silencing pathway (Carmell et al. 2002). There are four AGOs (AGO1–4) in mammals that can bind microRNAs (miRNAs) to posttranscriptionally repress target gene expression (Hock and Meister 2008). These AGOs and their associated miRNAs are important for mammalian cell differentiation and animal development (Ivey and Srivastava 2015; Muller et al. 2019).
AGO1 and AGO2 are expressed in mouse embryonic stem cells (mESCs) (Liu et al. 2021b). These two AGOs have opposite expression patterns during mESC differentiation: AGO2 protein levels increase, whereas AGO1 protein levels decrease (Liu et al. 2021b, 2024), suggesting that AGO1 and AGO2 have distinct roles in regulating cell fate decisions of mESCs. Consistent with this notion, recent findings indicate that AGO2 promotes differentiation predominantly via the pro-differentiation let-7 miRNAs (Liu et al. 2021a), whereas AGO1 maintains pluripotency via modulating protein folding independent of small RNAs in mESCs (Liu et al. 2024). These opposing functions of AGO1 and AGO2 in controlling mESCs self-renewal and differentiation raise the question of whether there are mutual regulations between AGO1 and AGO2 in mESCs.
Here, we show that AGO1 expression in mESCs is limited by AGO2 via a conserved let-7 miRNA-binding site in the 3′ UTR of AGO1 mRNA. Disruption of this let-7-miRNA-mediated repression increases AGO1 levels and compromises the exit pluripotency of mESCs. These results indicate that repressing AGO1 by AGO2 and let-7 miRNAs is important for the proper differentiation of mESCs, but also reveal a posttranscriptional regulatory event between the two different AGO paralogs.
RESULTS AND DISCUSSION
AGO2 represses AGO1 expression in mESCs
The recent observations of opposing functions of AGO1 and AGO2 in regulating pluripotency and differentiation in mESCs led us to examine whether there are mutual regulations between AGO1 and AGO2 (Liu et al. 2021a, 2024). We evaluated AGO1 and AGO2 protein levels in the wild-type (WT), AGO1Δ (knockout), and AGO2Δ mESCs by western blotting. Compared to those in the WT cells, AGO1 protein levels increased significantly in the AGO2Δ cells, which is consistent with previous observations (Ngondo et al. 2018), and AGO2 protein levels slightly elevated in the AGO1Δ cells (Fig. 1A). This result suggested that there are mutual regulations between AGO1 and AGO2 in mESCs. Here, we focused on the regulation of AGO2 on AGO1 due to the obvious (over twofold) increased AGO1 levels in the AGO2Δ cells.
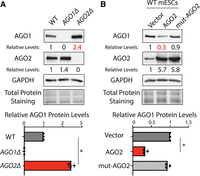
AGO2 represses AGO1 in mESCs. (A) Western blotting in the WT, AGO1Δ, and AGO2Δ mESCs cultured in the 15% FBS + Lif medium. (B) Western blotting in the WT mESCs expressing a vector, WT AGO2, and an RNA-binding mutant AGO2 in the 15% FBS + Lif medium. Data information: In A and B, representative western blots are shown, and the quantifications represent the means (±SD) of three independent experiments. GAPDH levels were used for normalization in calculating the relative protein levels. One-way ANOVA was used to determine the significance of the difference, (*) P < 0.05.
To further test whether AGO2 regulates AGO1, we performed gain-of-function experiments. When an exogenous AGO2 was expressed in the WT mESCs, AGO1 levels decreased significantly (Fig. 1B), indicating that AGO2 can repress AGO1 expression in mESCs. To examine whether this repression by AGO2 is dependent on the small-RNA pathways, we performed the same ectopic expression experiment using a mutant AGO2, in which mutations in the conserved PAZ domain abolish AGO2's binding to small RNAs (Ma et al. 2004). Although expressed at a similar level as the WT AGO2, the mutant AGO2 could not decrease AGO1 levels in the WT mESCs (Fig. 1B). Importantly, in either the WT mESCs with ectopically expressed AGO2 or the AGO2Δ mESCs, neither AGO1 mRNA levels nor protein stabilities were significantly altered (Supplemental Fig. S1), arguing that the changes in AGO1 protein levels were not due to protein stability alterations from the potential availability of free miRNAs (Johnston et al. 2010; Winter and Diederichs 2011; Martinez and Gregory 2013). These results collectively suggest that AGO2 represses AGO1 expression in mESCs via its associated small RNA(s), possibly miRNAs (e.g., translational repression).
Let-7 miRNAs can repress AGO1 expression in mESCs
To examine which miRNAs are involved in repressing AGO1 in mESCs, we identified the conserved miRNA-binding sites in the 3′ UTR of AGO1 mRNA using TargetScan (Agarwal et al. 2015) and then evaluated the expression levels of the corresponding miRNAs in mESCs using published small-RNA-seq data sets (Liu et al. 2021a). This analysis revealed that AGO1 mRNA's 3′ UTR contains a highly conserved binding site for let-7 miRNAs, which are expressed in mESCs. Let-7 miRNAs are a group of highly conserved miRNAs that promote stem cell differentiation by repressing numerous stemness factors (Bussing et al. 2008). Previous studies in human somatic cell lines show that the let-7 miRNAs can regulate AGO1 through a unique mechanism: inducing translational readthrough by binding to the let-7 site in the 3′ UTR of AGO1 mRNA, generating a C-terminus extended AGO1 (Singh et al. 2019). However, this specialized mechanism is not present in mESCs, as mESCs ribosome profiling data sets indicate that ribosomes terminate at the predicted stop codon, but not the first in-frame stop codon in the 3′ UTR of AGO1 mRNA (Supplemental Fig. S2). Moreover, cross-linking followed by immunoprecipitation and qRT-PCR indicated that AGO2 binds to AGO1 mRNA in a let-7 miRNA-dependent fashion (Supplemental Fig. S3 and see below). Altogether, these observations argue that AGO1 is regulated by let-7 miRNAs through the canonical miRNA mechanism in mESCs.
Two lines of evidence indicated that let-7 miRNAs can repress AGO1 expression in mESCs. First, when let-7a was ectopically expressed in mESCs (Fig. 2A), AGO1 protein levels decreased, which was similar to the evolutionarily conserved let-7 target, LIN28a (Fig. 2B). Second, we generated let-7Δ mESCs using genome editing, in which most of let-7 miRNA family members were specifically knocked out or knocked down (Fig. 2C). Again, similar to LIN28a (Rybak et al. 2008), AGO1 protein levels also increased in the let-7Δ mESCs compared to those in the WT mESCs (Fig. 2D). To examine whether the let-7 miRNAs are involved in the AGO2-mediated repression of AGO1, we ectopically expressed AGO2 in the let-7Δ mESCs. Different from that in the WT mESCs, the ectopically expressed AGO2 failed to repress AGO1 in the let-7Δ mESCs (Fig. 2E). These results argued that AGO2 represses AGO1 via the let-7 miRNAs in mESCs.
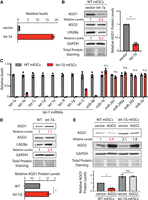
Let-7 miRNAs can repress AGO1 expression in mESCs. (A) Ectopic expression of let-7a in mESCs. The let-7a levels were determined by qRT-PCR, and U6 RNA was used for normalization. (B) Western blotting in the WT mESCs expression a vector and let-7a miRNAs. (C) qRT-PCR quantification of miRNAs in the WT and the let-7Δ mESCs. U6 RNA was used for normalization. (D) Western blotting in the WT and the let-7Δ mESCs. (E) Western blotting in the WT and the let-7Δ mESCs expressing either a vector or AGO2. Data information: In B, D, and E, representative western blots are shown, and the quantifications represent the means (±SD) of three independent experiments. GAPDH levels were used for normalization in calculating the relative protein levels. One-way ANOVA was used to determine the significance of the difference, (*) P < 0.05, (n.s.) not significant (P > 0.05).
AGO1 is repressed by let-7 miRNAs in mESCs
An important caveat in interpreting the results from let-7 overexpression (Fig. 2A) and let-7 knockout (Fig. 2D) is the potential pleiotropic effects. Let-7 miRNAs regulate hundreds of mRNAs. Overexpression or knockout let-7 miRNAs alters numerous let-7:mRNA interactions, making it difficult to determine whether the altered AGO1 protein levels are a primary effect of let-7/AGO2-mediated regulation or caused by secondary effects from altered regulations of other let-7 targets.
To address this issue and to specifically determine whether AGO1 is repressed by let-7/AGO2 in mESCs, we mutated the conserved let-7 miRNA binding site in the 3′ UTR of AGO1 mRNA into a restriction enzyme site using genome editing (Fig. 3A). Thereby, we could identify mESCs with biallelic mutations using PCR followed by restriction enzyme digestion (Fig. 3B), and then we verified the mutations in the 3′-UTR mutant mESCs using Sanger sequencing (Fig. 3C). In the 3′-UTR mutant mESCs, the levels of let-7 miRNAs were not significantly altered compared to those in the WT mESCs (Fig. 3D), whereas the AGO1 protein levels were increased (Fig. 3E). Moreover, in contrast to the results in the WT mESCs, ectopic expression of AGO2 in the 3′-UTR mutant mESCs did not significantly decrease AGO1 levels (Fig. 3F). Collectively, these results argue that AGO1 is repressed by AGO2 via the let-7 miRNAs in mESCs.
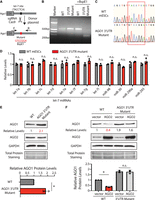
AGO1 is regulated by let-7 miRNAs in mESCs. (A) Mutating the let-7 miRNA binding site in AGO1 mRNA 3′ UTR via genome editing. (B) Genotyping of the AGO1 3′-UTR mutant. The PCR was performed using the oligos (F and R) indicated in A. (C) Sanger sequencing to verify the biallelic mutation of the let-7 miRNA-binding site in AGO1 mRNA 3′ UTR. The red box indicates the let-7 miRNA-binding site. (D) qRT-PCR quantification of miRNAs in the WT and the AGO1 3′-UTR mutant mESCs. U6 RNA was used for normalization. The results represent the means (±SD) of three independent experiments. One-way ANOVA was used to determine the significance of the difference, (n.s.) not significant (P > 0.05). (E) Western blot in the WT and the AGO1 3′-UTR mutant mESCs cultured in the 15% FBS + Lif medium. (F) Western blotting in the WT and the AGO1 3′-UTR mutant mESCs expressing either a vector or AGO2. Data information: In E and F, representative western blots are shown. All the quantifications represent the means (±SD) of three independent experiments. GAPDH levels were used for normalization in calculating the relative protein levels. One-way ANOVA was used to determine the significance of the difference, (*) P < 0.05, (n.s.) not significant (P > 0.05).
Repression of AGO1 by let-7 miRNAs is important for mESC differentiation
To determine the biological significance of let-7/AGO2-mediated repression of AGO1 in mESCs, we examined the self-renewal and differentiation of mESCs using the colony formation assay (Mulas et al. 2019) and the exit pluripotency assay (Betschinger et al. 2013), respectively. We first evaluated the let-7Δ mESCs. Consistent with the pro-differentiation activity of let-7 miRNAs, the let-7Δ mESCs displayed increased self-renewal (Fig. 4A) and decreased differentiation (Fig. 4B). Moreover, the pluripotency markers, NANOG and OCT4, remained at higher levels in the differentiating let-7Δ mESCs than those in the differentiating WT mESCs (Fig. 4C), further indicating that the let-7Δ mESCs had defects in exit pluripotency.

let-7-mediated repression of AGO1 is important for mESC differentiation. (A) Colony formation assay for the WT and the let-7Δ mESCs. (B) Exit pluripotency assay for the WT and the let-7Δ mESCs. (C) Western blotting in the differentiating WT and the let-7Δ mESCs. (D) Colony formation assay for the WT and the AGO1 3′-UTR mutant mESCs. (E) Exit pluripotency assay for the WT and the AGO1 3′-UTR mutant mESCs. (F) Western blotting in the differentiating WT and AGO1 3′-UTR mutant mESCs. Data information: In A–B and D–E, the colony morphology and AP intensity were evaluated via microscopy. In total, 100–200 colonies were examined each time to determine the percentage of undifferentiated colonies. In C and F, representative western blots are shown. All the quantifications represent the means (±SD) of three independent experiments. GAPDH levels were used for normalization in calculating the relative protein levels. One-way ANOVA was used to determine the significance of the difference, (*) P < 0.05.
To determine whether let-7/AGO2-mediated repression of AGO1 contributes to the differentiation defects in the let-7Δ mESCs, we examined the AGO1 3′-UTR mutant mESCs. Although to a lesser extent, the 3′-UTR mutant mESCs also had elevated self-renewal (Fig. 4D), compromised differentiation (Fig. 4E), and higher pluripotency markers in differentiating cells (Fig. 4F). These results strongly argue that repressing AGO1 by AGO2 via let-7 miRNAs is important for the proper differentiation of mESCs.
In sum, we show that AGO1 is repressed by AGO2 and let-7 miRNAs in mESCs, and this repression is important for the proper differentiation of mESCs. Among the four AGOs in mammals, mESCs express AGO1 and AGO2, where AGO1 protein levels are ∼2%–5% of those of AGO2 (Liu et al. 2021b; Sala et al. 2023). Consistent with this relative protein abundance, AGO2Δ inhibits global miRNA activity, whereas AGO1Δ does not (Liu et al. 2021a, 2024). Thus, at the endogenous level, AGO2 is the major AGO protein in the miRNA pathway in mESCs. AGO2Δ mESCs and mESCs with mutations in the key components (e.g., DICER, DGCR8) of the miRNA biogenesis pathway all displayed defects in exit pluripotency and differentiation (Kanellopoulou et al. 2005; Murchison et al. 2005; Wang et al. 2007; Ngondo et al. 2018), implying that AGO2 and the global miRNA activity promote differentiation in mESCs. Recent studies suggest that this pro-differentiation activity of AGO2 is largely mediated by the pro-differentiation let-7 miRNAs (Liu et al. 2024). In contrast to AGO2, our recent study showed that the endogenous AGO1 functions in a small RNA-independent manner by modulating protein folding to maintain stemness in mESCs (Liu et al. 2021a). Altogether, these previous results suggest that the two AGOs expressed in mESCs have opposing roles in controlling stem cell fate: AGO2 promotes differentiation, whereas AGO1 maintains self-renewal. The results from this study show that AGO1 mRNA is a functional target of the conserved pro-differentiation let-7 miRNAs and AGO2. This finding reveals a regulatory mechanism between the two AGO paralogs with opposing functions in controlling stem cell fate decisions. In human, different expression levels of AGO1 and AGO2 can predict distinct disease outcomes (Pantazopoulou et al. 2020). For instance, in renal cancer, high AGO2 protein levels are associated with unfavorable patient outcomes, whereas high AGO1 protein levels are associated with favorable outcomes (Liu et al. 2024). Thus, we speculate that the repression of AGO1 by let-7 miRNAs and AGO2 may also occur during the pathogenesis in human cells.
MATERIALS AND METHODS
All the cell lines, antibodies, and plasmids used in this study are listed in Table 1. The qPCR oligos and sgRNAs are listed in Supplemental Table S1. No primary data sets have been generated and deposited. The ribosome profiling data sets were from the E14 mESCs, as described previously (Ingolia et al. 2011).
Reagents and tools table
Cell culture
All the mouse ESCs used in this study were derived from ES-E14TG2a (ATCC CRL-1821), which was authenticated by STR profiling and tested negative for mycoplasma. mESCs were cultured on 0.5% gelatin-coated tissue cultured plates, in either 15%FBS + Lif medium or 2i + Lif medium.
CRISPR/Cas9-mediated genome editing in mESCs
For knockout mESCs’ generation, the targeting sgRNA was cloned onto the Bbs1 cut pWH464 [pSpCas9(BB)−2A-GFP (pX458), addgene plasmid 48138], and then 2 µg cloned plasmid was transiently transfected into mESCs using FuGENE 6. Forty-eight hours posttransfection, mESCs with the top 10% GFP intensity were single-cell sorted to 96-well plates. After 10 days, colonies were then picked and expanded for validation by genotyping PCR followed by sequencing and/or western blot analysis. The total let-7 knockout mESCs were generated by sequentially knocking out each of the let-7 miRNA family member using the sgRNA listed in the Reagent table. For generating the AGO1 3′-UTR mESCs, 2 µg of sgRNA-expressing vector (pWH464-based) and 1 µg of donor plasmid were used for the transient transfection, followed by single-cell sorting.
RNA isolation and RT-qPCR
Total RNA was extracted from mESCs using TRIzol, and genomic DNA was removed using DNase I treatment. For miRNA quantification, the cDNA was prepared using the Mir-X miRNA First Strand Synthesis Kit (Takara). For mRNA quantification, cDNA was synthesized using random hexamers and SuperScript II Reverse Transcriptase (Invitrogen), according to manufacturer instructions. qPCR was performed with the SsoAdvanced Universal SYBR Green Supermix (Bio-Rad) on a CFX96 Real-Time PCR System (Bio-Rad). The CLIP-qRT-PCR was performed in accordance with the protocols described previously (Liu et al. 2021a).
Immunoblotting analysis
Proteins were harvested in RIPA buffer (10 mM Tris-HCl pH 8.0, 140 mM NaCl, 1 mM EDTA, 1% Triton X-100, 0.5 mM EGTA, 0.1% SDS, 0.1% sodium deoxycholate, and Protease Inhibitor Cocktail) and quantified with the Pierce 660 nm Protein Assay Kit. Ten percent SDS-PAGE gel was used to resolve the proteins, and then the proteins were transferred to a PVDF membrane. The membrane was blocked with 3% nonfat dry milk and incubated with the indicated antibodies for western blot analysis. The membranes were then treated with the Clarity Western ECL Substrate (Bio-Rad), and the signal was detected by an ImageQuant LAS 500 instrument (GE Healthcare).
Colony formation assay and exit pluripotency assay
For the colony formation assay, mESCs were cultured at 500 cells/well in a gelatin-coated 12-well plate in the 15%FBS + Lif medium for 7 days. For the exit pluripotency assay, cells were seeded at 1000 cells/well in a gelatin-coated 6-well plate in the differentiation medium for 2 days, then cultured in 2i + Lif medium for another 5 days. The colonies were fixed with 4% PFA and then stained using an Alkaline Phosphatase Assay Kit (System Biosciences). The differentiation status of the colonies was manually evaluated under an Olympus CK2 microscope.
Embryoid bodies formation assay
For embryoid body formation assay, 3 × 106 mESCs harvested from 2i + Lif medium were seeded into 10 cm bacterial grade Petri dish in 10 mL differentiation medium (DMEM/F12 supplemented with 15% FBS, 1× penicillin/streptomycin, 0.1 mM nonessential amino acids, 2 mM L-glutamine, and 50 µM 2-mercaptoethanol) and maintained on a horizontal rotator with a rotating speed of 30 rpm. The medium was changed at day 3 and the resultant EBs were harvested at day 5.
Quantification and statistical analysis
Data were analyzed with Prism or R and represented as mean ± SD of three independent experiments. Data were considered significant if P < 0.05 using Student's t-tests or one-way ANOVA.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
We thank Dr. Qiuying Liu for the technical assistance. This work is supported by the Mayo Foundation for Medical Education and Research.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080426.125.
- Received February 17, 2025.
- Accepted March 10, 2025.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Gabrielle M. Schuh is the first author of this paper, “Repression of AGO1 by AGO2 via let-7 microRNAs facilitates embryonic stem cell differentiation.” Gabrielle is a postbaccalaureate student at the Mayo Clinic (Rochester, Minnesota, USA) with an interest in RNA biology and its clinical translation to improve patient outcomes.
What are the major results described in your paper and how do they impact this branch of the field?
Our study shows that AGO2 represses the expression of AGO1 via let-7 microRNAs, and this regulation plays a key role in facilitating mouse embryonic stem cell differentiation. These findings are an exciting glimpse into the importance of posttranscriptional regulation in controlling the balance between self-renewal and differentiation. By uncovering this dynamic between AGO proteins and let-7 microRNAs, we've provided a valuable framework for future research into RNA-based regulatory mechanisms and stem cell biology.
What led you to study RNA or this aspect of RNA science?
Like many of us, I gained a newfound appreciation for RNA research during the COVID-19 pandemic. I am particularly interested in understanding the mechanisms of RNA-mediated regulation and how these fundamental processes can be harnessed to address unmet medical needs. After earning my bachelor's degree, I followed this curiosity to join Dr. Wenqian Hu's lab at the Mayo Clinic to study RNA biology.
What are some of the landmark moments that provoked your interest in science or your development as a scientist?
I have always been interested in genetics research, exploring a broad range of topics during my undergraduate degree—from plant genetic engineering in biotechnology to neuron development and feeding behavior in fruit flies. However, supporting a close family member through leukemia clinical trials sparked my passion for applying genetics research to medicine. This led me to my current role researching posttranscriptional regulation at the Mayo Clinic. I am fascinated by the growing push for personalized medicine and excited to contribute to this field throughout my career.
If you were able to give one piece of advice to your younger self, what would that be?
I'd tell my younger self to take a deep breath and focus on the bigger picture, rather than letting imposter syndrome hold me back. Over time, with the support of my mentors at both North Carolina State University and the Mayo Clinic, I gradually built confidence to ask better questions and fully embrace the incredible research and mentorship opportunities I had. One of the best pieces of advice I received from my current PI and mentor, Dr. Wenqian Hu, was to expect mistakes as part of the learning process, which freed me from the self-inflicted pressure of perfection. I've learned that everyone, no matter their experience level, always has something new to learn, and that's not something to fear—it's what makes research exciting!
What are your subsequent near- or long-term career plans?
After completing my postbaccalaureate training at Mayo, I am pursuing a master's degree in genetic counseling. I am passionate about educational advocacy and increasing access to genetic information for diverse patient populations. My background in RNA research has strengthened my patience, perseverance, and analytical skills, which shape my approach to patient care and genetic test result interpretation. I am committed to staying active in research to better understand how RNA regulation influences gene expression, while fostering interprofessional collaboration to bridge research from bench to clinic and improve patient outcomes.








