
Investigating the role of RNA-binding protein Ssd1 in aneuploidy tolerance through network analysis
- Center for Genomic Science Innovation, University of Wisconsin-Madison, Madison, Wisconsin 53706, USA
- Laboratory of Genetics, University of Wisconsin-Madison, Madison, Wisconsin 53706, USA
- Corresponding author: agasch{at}wisc.edu
Abstract
RNA-binding proteins (RBPs) play critical cellular roles by mediating various stages of RNA life cycles. Ssd1, an RBP with pleiotropic effects, has been implicated in aneuploidy tolerance in Saccharomyces cerevisiae but its mechanistic role remains unclear. Here, we used a network-based approach to inform on Ssd1's role in aneuploidy tolerance, by identifying and experimentally perturbing a network of RBPs that share mRNA targets with Ssd1. We identified RBPs whose bound mRNA targets significantly overlap with Ssd1 targets. For 14 identified RBPs, we then used a genetic approach to generate all combinations of genotypes for euploid and aneuploid yeast with an extra copy of chromosome XII, with and without SSD1 and/or the RBP of interest. Deletion of 10 RBPs either exacerbated or alleviated the sensitivity of wild-type and/or ssd1Δ cells to chromosome XII duplication, in several cases indicating genetic interactions with SSD1 in the context of aneuploidy. We integrated these findings with results from a global overexpression screen that identified genes whose duplication complements ssd1Δ aneuploid sensitivity. The resulting network points to a subgroup of proteins with shared roles in translational repression and P-body formation, implicating these functions in aneuploidy tolerance. Our results reveal a role for new RBPs in aneuploidy tolerance and support a model in which Ssd1 mitigates translation-related stresses in aneuploid cells.
Keywords
INTRODUCTION
RNA-binding proteins (RBPs) are critical mediators of cellular responses, with highly interconnected functions often spanning multiple stages of RNA life cycles (Hogan et al. 2008; Müller-Mcnicoll and Neugebauer 2013; Goswami et al. 2024). Many RBPs are both complex in sequence—with many functional domains—and pleiotropic in either functions or phenotypes they affect (Hentze et al. 2018; Corley et al. 2020). As a group, they are integral to all facets of posttranscriptional regulation, including but not limited to RNA transcription, nuclear export, processing, subcellular localization, translation, stability, and decay (Dreyfuss et al. 2002; Mazumder et al. 2003). It is not uncommon for RBPs to operate across more than one of these regulatory realms. The pleiotropic nature of RBP functions often presents a challenge in elucidating their biochemical mechanisms and how those mechanisms impact cellular phenotypes.
Previous work in our laboratory found that RBP called Ssd1 enables budding yeast to tolerate extra chromosomes, a state known as aneuploidy (Hose et al. 2020; Rojas et al. 2024). Despite the clear implication of Ssd1 in aneuploidy tolerance, the mechanistic basis for its involvement has remained unknown. Many different studies identified Ssd1 in suppressor screens, linking it to processes ranging from cell wall integrity and thermotolerance to tRNA modifications, aging, quiescence, and now aneuploidy (Kaeberlein and Guarente 2002; Li et al. 2009; Hu et al. 2018; Miles et al. 2019; Xu et al. 2019; Hose et al. 2020). Ssd1 is important for the delivery of cell wall-associated mRNAs to sites of polarized growth under normal conditions or to RNA–protein granules called P-bodies (PBs) during stress (Kurischko et al. 2011a,b). In fact, Ssd1 is a component of PBs, although found in only a subset of PBs in the cell (Xing et al. 2020). Ssd1 has also been implicated in translational repression, based on global polysome profiles as well impacts on protein abundance from specific directly bound transcripts (Jansen et al. 2009; Wanless et al. 2014; Hu et al. 2018; Hose et al. 2020). Unlike its mammalian ortholog Dis3L2 that degrades poly-uridylated transcripts, Ssd1 is catalytically inactive but directly binds several hundred mRNAs at sequences primarily in the 5′ UTR and coding regions (CDSs) (Hogan et al. 2008; Jansen et al. 2009; Ohyama et al. 2010; Wanless et al. 2014; Hose et al. 2020; Bayne et al. 2022). The RNAs bound by Ssd1 are enriched for those encoding specific functional groups, including cell cycle regulation, cell wall integrity, RNA metabolism, budding, and sterol transport, but they also include many mRNAs outside of these functional groups (Hogan et al. 2008; Hose et al. 2020; Bayne et al. 2022). Although Ssd1 has been linked to mRNA localization and translational control (Hogan et al. 2008; Jansen et al. 2009; Kurischko et al. 2011b; Wanless et al. 2014; Hu et al. 2018; Xu et al. 2019; Khonsari et al. 2021), how it acts mechanistically is not known. Furthermore, it is unclear which of its functions, bound transcripts, or their related processes influence aneuploidy sensitivity.
One strategy to pinpoint which of Ssd1's functions are important is to explore other RBPs that may work with Ssd1 by binding the same transcripts. Previous studies in Saccharomyces cerevisiae and humans revealed that subsets of RBPs show substantial overlap in mRNA targets (Hogan et al. 2008; Achsel and Bagni 2016; Kershaw et al. 2023). This interconnectivity is an asset to elucidate RBP networks, which can be constructed based on the overlap in RBP targets and on physical interactions between RBPs. The resulting networks can reflect on regulatory principles and shared regulation across specific mRNAs. For example, yeast mRNAs can be effectively categorized into discrete modules based on the RBPs that bind them (Costello et al. 2015; Kershaw et al. 2023). Features of translational control can be inferred by relating these regulatory modules to the biophysical properties of the affected mRNAs (Costello et al. 2015; Kershaw et al. 2023). Thus, a network perspective provides greater biological insights into the regulation of mRNA targets than if their interactions were examined piecemeal.
Here, we used a network-based approach to better understand the role of Ssd1 in aneuploidy tolerance. Under the hypothesis that Ssd1 may function in this role with other proteins that bind the same RNAs, we first identified a network of RBPs whose bound mRNAs overlap with Ssd1's known targets. We then experimentally perturbed this network by deleting RBPs in euploid and aneuploid cells, alone and in an ssd1Δ background. This allowed us to investigate the influence of each RBP on aneuploidy tolerance, alone and in the context of SSD1 deletion. We integrated these findings with recent results from a global gene-duplication screen to further refine hypotheses about Ssd1's functions in aneuploidy tolerance across chromosome duplications. Our results implicate a subnetwork of RBPs and translation factors whose deletion or overexpression influences aneuploidy tolerance in yeast. This work adds to a growing body of evidence that Ssd1 and its broader RBP network mediate aneuploidy tolerance through functions related to translational suppression. We discuss models for how translational repression could influence the tolerance of extra chromosomes.
RESULTS
Ssd1 is part of a highly interconnected RBP network
We previously used RNA-immunoprecipitation and sequencing (RIP-seq) to identify 286 mRNAs bound by Ssd1 (Hose et al. 2020). To identify other RBPs that might act with Ssd1 including in mediating aneuploidy tolerance, we identified RBPs that bind more Ssd1 targets than expected by chance. We first collated from public data sets available at the time the mRNAs bound by 65 different RBPs that share at least one Ssd1 target (Fig. 1, see Materials and Methods and Supplemental Table S2). We then identified the subset of RBPs whose bound mRNAs statistically significantly overlap Ssd1-bound mRNAs (FDR < 0.05, hypergeometric test, see Materials and Methods).
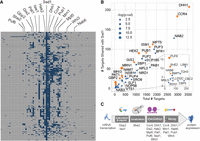
Fourteen RBPs show significant overlap in targets with Ssd1. (A) The matrix shows 280 mRNAs bound by Ssd1 (rows) and bound status (dark blue box) by each of 65 RBPs (columns). RBPs whose targets significantly overlap Ssd1 targets (FDR < 0.05) are labeled along the top. (B) A plot showing the number of mRNAs bound by each of 39 RBPs that share at least three RNA targets with Ssd1 (x-axis, see Materials and Methods) and the fraction of those targets that overlap Ssd1-bound targets (y-axis). Significant RBPs are shown in orange circles, where the size of the circle is proportionate to the enrichment P-value as indicated in the key. (C) A summary of 14 enriched RBPs and the functional processes they participate in.
This approach identified 14 RBPs as part of the Ssd1 network; this network extends beyond a recent analysis of RBPs that bind cell wall transcripts, which represent a subset of the mRNAs analyzed here (Hall and Wallace 2022). Several of these proteins are implicated in multiple processes related to RNA biogenesis (Fig. 1C), and together, the network spans functions related to most stages of mRNA life cycles, including transcript maturation and nuclear export (Isw1 and Gbp2), subcellular localization (She2), translational regulation (Gis2, Mpt5, Puf6, Whi3, and Hek2, among others), and roles in mRNA decay and stability (Ccr4, Dhh1, Pat1, Pub1, Mrn1, Nab6, and Whi3). Like Ssd1, many of these RBPs were previously found to bind cell wall-related transcripts (She2, Hek2, Gbp2, Pub1, Mpt5, Mrn1, and Nab6) or have been otherwise implicated in cell wall integrity (Mpt5, Ccr4, and Hek2) (Kaeberlein and Guarente 2002; Hogan et al. 2008; Ito et al. 2011; Hall and Wallace 2022; Bresson et al. 2023). However, overlap with Ssd1 targets remained significant (FDR < 0.05) for all of these proteins except Pub1 and Nab6 when transcripts corresponding to the GO term “cellular wall” were removed, suggesting that Ssd1's network is not restricted to cell wall transcripts alone. We previously reported that Ssd1-bound transcripts are enriched for mRNAs with structure and mRNAs encoding proteins with an elevated proportion of intrinsic disorder (Hose et al. 2020). We investigated these features in targets bound by the 14 RBPs identified here (Fig. 2). These RBPs varied in their propensity to bind structured mRNAs (Kertesz et al. 2010). However, we found that, like Ssd1, 10 of the 14 RBPs bind mRNAs that encode proteins with an elevated amount of predicted disorder (Dosztányi 2018; Mészáros et al. 2018). The significance of this is unclear but could reflect the regulation of proteins with specific biophysical properties.
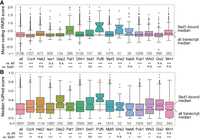
RBPs share enriched features among bound targets. Distributions of PARS scores (Kertesz et al. 2010) representing RNA secondary structure (top) or encoded-protein IUPred scores (Dosztányi 2018) representing predicted disorder (bottom) for RNAs bound by each RBP; see Materials and Methods for details. The number of mRNAs plotted is listed above each RBP name. (***) FDR < 0.00001, (**) 0.00001 ≤ FDR < 0.001, (*) 0.001 ≤ FDR < 0.05, two-sided Mann–Whitney U test with Benjamini and Hochberg FDR correction (Benjamini and Hochberg 1995).
Interestingly, in addition to sharing mRNA targets with Ssd1, many of these RBPs interact physically and/or genetically with Ssd1 or other RBPs in the network (Fig. 3). Ccr4 in particular, along with Ssd1 itself, exhibits many genetic interactions with other RBPs in the network. Furthermore, 11 of the 14 proteins can localize to membraneless condensates including PBs or stress granules (SG) that form after stress (for review, see Escalante and Gasch 2021). Indeed, Ssd1 is known to localize to a subset of PBs and SG during various stresses (Kurischko et al. 2011a; Xing et al. 2020). Together, these results suggest that Ssd1 and its targets are part of a highly interconnected network of proteins that act on similar targets.
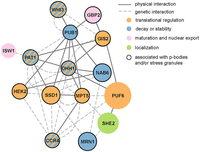
Ssd1 shares a regulatory network with 14 other RBPs. Each circle represents an RBP and each line represents a physical or genetic interaction according to the key. With the exception of Ssd1, nodes are scaled to reflect the proportion of that RBP's targets that are also bound by Ssd1. Colors depict the primary functional groups of each RBP as shown; since many RBPs are involved in multiple functions, this figure depicts simplified annotations.
Many of the RBPs in Ssd1's regulatory network exhibit synthetic genetic interactions with ssd1Δ and aneuploidy
We were especially interested if any of these RBPs, like Ssd1, play a role in aneuploidy tolerance. To investigate this, we devised a genetic strategy to sensitively measure the interaction of each RBP deletion with SSD1 deletion and chromosome XII (Chr12) duplication, alone and in combination. We chose this chromosome duplication because it is a good representative in our strain background and was available at the time of our analysis. Like in many other aneuploid strains, Chr12 duplication is tolerated in wild-type cells but very deleterious in an ssd1Δ background (Rojas et al. 2024). At least part of the dependence on Ssd1 is due to the amplification of specific mRNAs on Chr12 that are very deleterious in a euploid strain lacking SSD1 (Dutcher et al. 2024).
We engineered a diploid parent in our wild strain background, oak-soil strain YPS1009, that is trisomic for Chr12 (3nChr12) and hemizygous at both the SSD1 locus and the locus of each RBP of interest. We then dissected meiotic products to isolate discrete genotypes that all grow side-by-side in the same environment (Fig. 4A). Colony size was determined after a defined growth period and used as a proxy for growth rate. Each colony's genotype was subsequently determined using drug markers to identify SSD1 and RBP knockouts, and euploid or aneuploid status was identified by flow cytometry to infer DNA content (Chen et al. 2012). Progeny were diploid due to mating-type switching in this strain background, with the exception of she2Δ and whi3Δ. These two mutants harbored DNA content consistent with mixed ploidy, likely caused by impaired mating or mating-type switching (Jansen et al. 1996; Nash et al. 2001) and were thus reconstructed as haploids for further analysis. We compared colony sizes of all genotypes from a single parental strain grown in parallel under identical conditions, including strains with and without SSD1, each RBP of interest, an extra copy of Chr12 (hereafter referred to simply as “aneuploidy”), and every combination thereof. Statistically significant genetic interactions were identified if the measured colony size in the double mutant was more or less than expected based on the independent action of each RBP and of aneuploidy (through a multiplicative model, see Materials and Methods). This method implicated RBP deletions that alleviate or exacerbate aneuploidy stress in wild-type cells and in the context of ssd1Δ (see Materials and Methods).
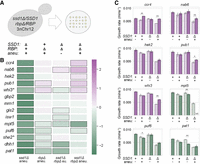
RBPs with shared targets exhibit synthetic genetic interactions with ssd1Δ and aneuploidy. (A) Dissection of a parent strain trisomic for chromosome 12 (3nChr12) and hemizygous at SSD1 and each RBP locus of interest yields colonies with all possible genotypes, a subset of which are represented in B. (B) The log2(fold-difference) in colony size comparing mean observed size versus mean expected size across batches (see Materials and Methods), according to the color scale, for genotypes indicated along the top in A. Black outlined boxes identify significant effects (FDR ≤ 0.05). Asterisk indicates haploids. (C) Average and standard deviation of exponential growth rates of the denoted strains under standard conditions; all strains that are shown for each mutant lack the RBP in question. Measured growth rates are represented as solid bars; expected growth rates are represented by light colors and dashed outlines. The average growth rate of the wild-type parent is 0.0081 min−1. Mutants with primarily a positive (alleviating) effect are shown in purple; mutants that mostly exacerbated defects are shown in green. (**) P-value ≤ 0.01, (*) P ≤ 0.05, (+) P ≤ 0.1, two-tailed, replicate-paired t-test comparing observed versus expected values. (UC) Unculturable strain.
Our strategy revealed a remarkably high hit rate, with 10 of the 14 RBPs having a significant impact aneuploidy and/or aneuploidy in the context of ssd1Δ (Fig. 4B). The approach confirmed the expected interaction between ssd1Δ and Chr12 duplication in every single dissection, recapitulating the very strong negative genetic interaction between ssd1Δ and aneuploidy (see Fig. 4B, column 1). No other RBP deletion sensitized cells to chromosome amplification to nearly the same extent as ssd1Δ; however, mpt5Δ, puf6Δ, she2Δ, dhh1Δ, and pat1Δ all sensitized cells to aneuploidy to some degree in wild-type cells (Fig. 4B, column 2) (FDR < 0.05). Of these, a subset (mpt5Δ, dhh1Δ, and pat1Δ) also had negative interactions with ssd1Δ in a euploid context (Fig. 4B, column 3), consistent with previous reports (Moriya and Isono 1999; Kaeberlein and Guarente 2002; Costanzo et al. 2016). Only mpt5Δ and puf6Δ had significant negative interactions with ssd1Δ in an aneuploid background, although the effect of dhh1Δ just missed the significance cutoff (Fig. 4B, column 4). Thus, several mutants displayed negative genetic interactions with aneuploidy, with ssd1Δ, or both, such that they exacerbate the sensitivity to extra Chr12.
Interestingly, a large group of significant effects comprised RBPs whose deletion resulted in larger than expected colonies for aneuploid double-mutants (ssd1Δ rbpΔ) (Fig. 4B, column 4). Deletion of CCR4, NAB6, HEK2, PUB1, or WHI3 in combination with ssd1Δ and Chr12 duplication all produced colonies larger than expected from a multiplicative model (see Materials and Methods), to varying degrees. In all cases, this positive genetic interaction was limited to or stronger in the aneuploid double mutant, producing a larger effect than seen in the euploid double mutant. Thus, deletion of over a third of the queried RBPs in Ssd1's network produces significant, reproducible differences in colony size that are specific to the context of ssd1Δ and aneuploidy.
One possibility is that cell size and/or colony morphology, rather than growth rate, explain the observed differences in colony size. To measure effects that are truly due to growth rate differences, we measured the growth rate of select RBP deletions in liquid culture, this time in a haploid background to identify ploidy-independent effects (Fig. 4C). All five positive genetic interactions between RBP deletion, ssd1Δ, and aneuploidy were validated in liquid culture (although the ssd1Δ hek2Δ aneuploid missed the cutoff for significance). (We noted substantial colony size variability in streaks of whi3Δ and ssd1Δ whi3Δ strains, which could suggest populations of mixed ploidy, consistent with reports of increased rates of ploidy increase in whi3Δ mutants from a previous study [Schladebeck and Mösch 2013].) Growth rate measurements likewise validated most negative genetic interactions between ssd1Δ and aneuploidy: In the most striking example, ssd1Δ mpt5Δ aneuploids proved entirely unculturable, consistent with the dramatic growth defect observed from our colony size assay. Interestingly, puf6Δ and pat1Δ liquid growth rates produced mixed results: both deletions exacerbated the sensitivity of SSD1+ cells to Chr12 duplication, but slightly alleviated the growth defect of ssd1Δ aneuploid cells (though missed the cutoff for significance). In summary, most phenotypes from our colony-size assay were validated in liquid growth, confirming that the genetic interactions affect growth rate. Thus, we identified several new proteins in Ssd1's network that also exhibit intriguing genetic interactions with aneuploidy.
Incorporating beneficial gene duplications expands the Ssd1-aneuploidy network
Through other work, we recently identified gene duplications that alleviate the growth defect of ssd1Δ aneuploids carrying different chromosome amplifications, including Chr12. Genes that benefited multiple ssd1Δ aneuploids were enriched for genes involved in RNA metabolism and translation (Dutcher et al. 2024). To further refine hypotheses about Ssd1's role in aneuploidy tolerance, here we integrated those results with the RBP network presented here. We started with 316 gene duplicates that specifically benefit the ssd1Δ Chr12 aneuploid strain (FDR < 0.05) (see Dutcher et al. 2024). This group represented diverse functions but was enriched for proteins involved in mRNA processing (P = 7.2 × 10−5, hypergeometric test) and regulation of translation (P = 8.2 × 10−3). We identified the subset of 66 genes that belonged to these and related functional groups (namely, rRNA processing and ribosome biogenesis, splicing, and tRNA-related, see Materials and Methods) and combined them into a joint network with RBPs identified above. We considered gene duplicates beneficial to ssd1Δ cells with a Chr12 duplication as well as the subset of those genes that also benefit other ssd1Δ aneuploids.
The resulting network suggested functional connections among beneficial gene duplicates and impactful RBPs (Fig. 5). First, the network clarified the functions of beneficial genes: most of the genes related to rRNA, tRNA, and ribosome biogenesis were only scored as beneficial upon duplication of Chr12, which contains the rDNA locus. This suggests that some of the genes may complement Ssd1-dependent effects related to ribosome biogenesis or function. In contrast, duplication of other genes, including those linked to splicing, mRNA processing, and especially translation, were beneficial to multiple chromosome duplications, strongly suggesting more generalizable effects in ssd1Δ aneuploids. Interestingly, several of the proteins encoded by beneficial gene duplicates interact physically with impactful RBPs (Fig. 5, orange edges). Several interacted with more RBPs in our original network than expected by chance (based on their total number of protein interactions, FDR < 0.05, hypergeometric test) (Fig. 5, large nodes). Many of these are translational regulators. In fact, the group of 15 beneficial genes that interact with more RBPs than expected was statistically significantly enriched for translational regulators (P = 2.4 × 10−5, hypergeometric test). These enrichments further suggest that Ssd1's role in aneuploidy tolerance may be linked to translation (see Discussion).
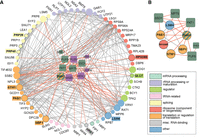
RBPs that genetically interact with aneuploidy physically interact with duplicated proteins that complement aneuploid ssd1Δ defects. (A) Sixty-five peripheral nodes (circles) represent genes related to translation and RNA biology (colored with simplified annotations according to the legend) whose duplication is especially beneficial to Chr12 ssd1Δ aneuploids. Central nodes (squares) represent significant RBPs from Figure 4, colored purple or green if the deletion alleviates or exacerbates ssd1Δ Chr12 sensitivity. Nodes with a black border (label with colored background) indicate gene duplications that confer a benefit in multiple ssd1Δ aneuploids carrying different chromosome amplifications. Edges represent physical interactions between encoded peripheral proteins (gray) or with an RBP (orange). (B) Complementary network of gene duplicates beneficial to multiple ssd1Δ aneuploids (circles) and RBP deletions (squares) that exacerbate the aneuploid growth defect. DHH1 belongs to both groups.
To further investigate, we generated a subnetwork of genes linked to complementary impacts on ssd1Δ aneuploidy: a benefit when overproduced and/or a defect when deleted. We reasoned that the complementary effects of their genetic perturbation would tap into a network of gene products that function together in the same required process. We took six proteins that are beneficial to multiple ssd1Δ aneuploid strains (Dutcher et al. 2024) and show more physical interactions with RBPs studied here (FDR < 0.05). We then combined this list with the six RBPs whose deletion exacerbates growth defects in the presence of Chr12 duplication (Fig. 4B). The DEADBOX RNA helicase Dhh1 was identified in both studies: its duplication alleviates the ssd1Δ aneuploidy growth defects, whereas its deletion sensitizes cells to Chr12 duplication.
The resulting subnetwork implicated deep connections between RBPs (all of which share targets with Ssd1) and genes whose duplication benefits aneuploidy tolerance in the absence of SSD1. Remarkably, nearly all of these proteins can be related to translational control. This includes several proteins implicated in translation or ribosome functions (Pab1, Rps28b, Stm1), translational repression specifically (Sbp1, She2 and Puf6, Mpt5, Pab1, Dhh1, along with Ssd1), or RNA decay (Lsm4, Pat1, Dhh1). Strikingly, most of these proteins either localize to PBs or influence their function (Pat, Dhh1, Lsm4, Mpt5, Ssd1, Rps28b, and others). Together, our computational and experimental analyses support a role for translational control in mediating aneuploidy tolerance. Implications and models are discussed below.
DISCUSSION
While Ssd1 is clearly involved in aneuploidy tolerance, its precise function in mediating stress associated with chromosome amplification has remained elusive. Ssd1 has been linked to translational regulation (including but not exclusively translational repression), mRNA localization, and PBs (Jansen et al. 2009; Ohyama et al. 2010; Kurischko et al. 2011a; Wanless et al. 2014). It is known to impact cell wall biology, aging, and quiescence among other phenotypes (Kaeberlein and Guarente 2002; Miles et al. 2019; Xu et al. 2019). The apparent complexity of Ssd1's role is not unique: many RBPs are pleiotropic, either in function (e.g., multiple roles within the cell) or in the phenotypes they impact. For instance, it is common for RBPs to participate in multiple, different stages of mRNA life cycles, or even to have secondary functions unrelated to their RNA-binding activity (for review, see Hentze et al. 2018). Consequently, pinpointing the specific functions of Ssd1 that contribute to aneuploidy tolerance presents a significant challenge.
This study took a network approach to identify RBPs whose mRNA targets are shared with Ssd1, under the hypothesis that they may function together to regulate those mRNAs and impact aneuploidy tolerance. We found that Ssd1 shares more targets than expected with 14 other RBPs. Remarkably, 10 of these 14 RBPs (over 70%) influenced the growth of cells with an extra copy of Chr12, when the RBP is deleted in wild-type aneuploids or in the context of SSD1 deletion. We identified several RBPs whose deletion sensitized cells to Chr12 duplication in their own right, including Dhh1, Pat1, and to a lesser extent Mpt5 and others. Thus, this work identified new players in the modulation of Chr12 aneuploidy tolerance, and identified a refined network that suggests associated functions for future investigation. It is notable, however, that none of the new RBPs impacts aneuploidy tolerance to the extent of Ssd1.
Combining experimental results from several assays, interrogating Chr12 duplication but also others, presented a subnetwork of proteins that influence aneuploidy tolerance in ssd1Δ cells. One of the most striking commonalities among these proteins is their connections to translational repression and/or PB function. Poly(A) binding protein Pab1, whose duplication benefits multiple ssd1Δ aneuploids (Dutcher et al. 2024), is important for efficient translation initiation but also plays a role in regulating repression and can also localize to PBs (Brengues and Parker 2007; Chritton and Wickens 2010; Brambilla et al. 2019). The helicase Dhh1 and decapping factor Pat1 are also important for translational repression and are essential PB components: cells lacking both proteins have pronounced defects in PB formation and translational suppression (Coller and Parker 2005; Marnef and Standart 2010; Zeidan et al. 2018; Vindry et al. 2019; Vijjamarri et al. 2023). We found that deletion of either factor sensitizes cells to Chr12 duplication, even in the presence of Ssd1. Deletion of another PB factor, MPT5, also produced a mild sensitivity to Chr12 aneuploidy that was severe in the absence of SSD1. Mpt5 recruits specific mRNAs to PB, impacting either their decay or storage, depending on the target (Wang et al. 2018). Additional PB proteins were implicated by our network analysis (Fig. 5B), including decapping and PB factor Lsm4, translational regulator Stm1 (which associates with nontranslating ribosomes, can promote decapping, and is a suppressor of pat1Δ temperature sensitivity) (Balagopal and Parker 2009, 2011), ribosomal protein Rps28b whose 3′ UTR serves as a scaffold for PB assembly (Fernandes and Buchan 2020), and translational repressor Sbp1 that also has a role in PB disassembly (Roy et al. 2022). Thus, the majority of proteins represented in the subnetwork shown in Figure 5B are directly linked to PB physiology. The implication of this result is not clear, since the exact function of PB remains under investigation (for review, see Escalante and Gasch 2021). PBs have long been associated with translational silencing, although their formation may be a consequence, rather than a driver, of translational repression (Eulalio et al. 2007). Many PB components function outside of PBs in roles related to translational repression and/or mRNA decay. We hypothesize that their implication here underscores the importance of translational repression for aneuploidy tolerance and a specific role for Ssd1 in that process. Given that many of these RBPs function during times of stress, an interesting avenue for future work is to test their role in mediating aneuploid cells’ response to suboptimal conditions.
In contrast, several genes whose deletion ameliorates ssd1Δ aneuploidy sensitivity are associated with mRNA stability, especially at specific classes of mRNAs including cell wall transcripts and mRNAs with upstream open reading frames (uORFs), both of which are enriched among Ssd1 targets. Nab6 stabilizes mRNAs encoding cell wall proteins (Bresson et al. 2023), while Pub1 stabilizes uORF-containing mRNAs by preventing nonsense-mediated decay (NMD) (Ruiz-Echevarría and Peltz 2000). Deletion of these RBPs is thus predicted to destabilize bound transcripts, which could counteract a loss of translational repression in the ssd1Δ strain. Interestingly, Ssd1 targets are enriched for both cell wall mRNAs (Hogan et al. 2008; Hose et al. 2020) and transcripts with uORFs (P = 9.4 × 10−3, see Materials and Methods) (Ingolia et al. 2009; Spealman et al. 2023). Several genes encoding cell wall proteins (but also many others) are especially deleterious in ssd1Δ euploid cells and are encoded on Chr12, perhaps contributing to its toxicity (Dutcher et al. 2024). Another gene whose deletion alleviated the ssd1Δ Chr12 aneuploid growth defect was Ccr4, a core component of the Ccr4–Not deadenylase complex that influences mRNA decay, including of some cell wall mRNAs (Tucker et al. 2001; Ito et al. 2011).
Integrating these details suggests several models through which Ssd1 could function. Our results point to a role for Ssd1 in translational repression to mediate aneuploidy tolerance, of Chr12 studied here and other chromosomes investigated elsewhere (Hose et al. 2020; Dutcher et al. 2024; Rojas et al. 2024). We previously ruled out a model in which Ssd1 globally silences all amplified mRNAs encoded on the extra chromosome (Hose et al. 2020). Past proteomic work identified few proteins whose abundance increased in the ssd1Δ Chr12 aneuploid compared to the wild-type aneuploid in a way that could not be explained by underlying mRNA changes; only eight of the affected proteins come from Ssd1 targets, and only one of those is encoded on the amplified chromosome. Furthermore, we found no strong evidence for a singular role in mRNA stability that affected all of the Ssd1-bound targets. Roughly 40% of Ssd1-bound mRNAs (117 out of 280) showed statistically significant expression differences in ssd1Δ versus wild-type Chr12 aneuploid cells (Hose et al. 2020); however, among these, there was a similar number of transcripts with higher versus lower abundance, and both of those groups were strongly enriched for functional categories that may reflect a regulated physiological response. Thus, if Ssd1 plays a role in mRNA stability, that role must be different at different mRNAs. Most Ssd1-bound transcripts are not more toxic when duplicated in isolation in ssd1Δ euploids, indicating that loss of repression of single transcripts is unlikely to drive aneuploidy sensitivity of ssd1Δ cells (Dutcher et al. 2024).
Instead, Ssd1 may help to mitigate the indirect effects of aneuploidy on translational efficiency or fidelity. If chromosome amplification taxes translation, perhaps through an overabundance of many translated mRNAs, this could produce a cellular state that is sensitized to further translational stress. Several lines of evidence support a role for Ssd1 in managing that stress. First, aneuploid yeast are sensitive to the aminoglycoside nourseothricin (NTC), which binds the ribosome to disrupt elongation and cause tRNA misincorporation (Haupt et al. 1978; Ling et al. 2012; Hose et al. 2020; Dutcher et al. 2024). Deletion of SSD1 renders aneuploid cells—but not euploids—extremely sensitive to NTC (Dutcher et al. 2024). Thus, the combination of SSD1 deletion and chromosome amplification exacerbates sensitivity to this translational inhibitor. Second, SSD1 deletion sensitizes euploid cells to mutation of the Elongator tRNA-modification complex. Loss of Elongator function increases tRNA misincorporation at specific codons (Karlsborn et al. 2014; Xu et al. 2019, 2020). Third, this connection between Ssd1 and translational fidelity may explain a recent result from other work in our laboratory that showed that overexpression of tRNAs improves the growth of multiple ssd1Δ aneuploid strains (Rojas et al. 2024). Increased abundance of specific tRNAs can enhance translation through cognate codons, which may explain why tRNA up-regulation alleviates growth defects in several ssd1Δ aneuploids (Kramer and Farabaugh 2007; Frumkin et al. 2018; Rak et al. 2018; Rojas et al. 2024). Finally, we recently found that SSD1+ aneuploids in the YPS1009 strain background can tolerate extra chromosomes during log-phase growth, but they have a major defect entering quiescence and maintaining normal life span—these premature aging defects are due in part to defects in the ribosome quality control (RQC) pathway that responds to stalled ribosomes (Sitron and Brandman 2020; Escalante et al. 2024). Remarkably, simply overexpressing stoichiometrically limiting RQC subunits partly alleviates aneuploid defects (Escalante et al. 2024).
One possibility is that Ssd1, and potentially other factors identified here, regulate mRNAs that are either error prone or difficult to translate. If aneuploid cells are already taxed, perhaps if overabundant mRNAs from the amplified chromosome titrate key translation factors, deletion of Ssd1 and misregulation of specific targets could exacerbate problems. Interestingly, Kershaw et al. (2023) recently identified seven RNA regulons based on shared RBP interactions. Ssd1-bound mRNAs are overrepresented (P = 5.4 × 10−15, hypergeometric test) in a regulon characterized by long, relatively structured mRNAs with low ribosome occupancy under standard conditions (Cluster 3). Indeed, Ssd1 targets are more structured relative to all yeast mRNAs (Kertesz et al. 2010) (P = 2.0 × 10−6, see Materials and Methods). Thus, Ssd1 targets could have unique requirements for efficient translation, such that proper regulation of these mRNAs (or a subset thereof) is particularly important in aneuploids. Further studies will be required to elucidate the mechanistic underpinnings of these interactions, as well as to better understand the fundamental liabilities associated with aneuploidy stress. In all, this study identified new RBPs that influence aneuploidy tolerance, uncovered novel genetic interactions with SSD1 in the context of aneuploidy, and implicated functions important for tolerance of chromosome duplication.
MATERIALS AND METHODS
Strains and growth conditions
Strains used in this study are listed in Supplemental Table S1. Gene knockouts were generated by homologous recombination of HYG-MX into the designated locus, followed by diagnostic PCR to confirm the absence of one copy of the target RBP gene. Colony-based growth was scored as described below. Liquid growth curves were performed in YPD (1% yeast extract, 2% peptone, 2% dextrose) in test tubes at 30°C with shaking. Specifically, liquid cultures were inoculated, grown for 3–4 h to minimize extra chromosome loss, and optical density at 600 nm (OD600) was measured over time to calculate exponential growth rates. Genetic interactions were determined as described below.
mRNA target overlap analysis
We identified 65 proteins that shared targets with our 286 previously identified Ssd1-bound mRNAs (Hose et al. 2020) using the BioGRID database (Oughtred et al. 2016) accessed via YeastMine (data obtained October 2019) (Balakrishnan et al. 2012) along with several other direct studies available at the time (Hogan et al. 2008; Shahbabian et al. 2014; Delaveau et al. 2016). Of these 65 RBPs, 39 shared at least three RNA targets with Ssd1, where SSd1 targets were defined as those measured in our laboratory previously (Hose et al. 2020). We identified the subset of these RBPs that bound more Ssd1 targets than expected by chance, using hypergeometric tests (assessing a total of 5944 mRNAs) with Benjamini–Hochberg FDR correction (Benjamini and Hochberg 1995), taking FDR < 0.05 as significant. This identified the 14 RBPs shown in Figure 1. Each RBP's bound RNA targets were analyzed for secondary structure using the PARS (parallel analysis of RNA structure) data set from Kertesz et al. (2010), comparing the distributions of scores of bound targets (if available) to the whole transcriptome data set (n = 3196) using a two-sided Mann–Whitney U (MWU) test, with FDR calculated using the Benjamini and Hochberg procedure (Benjamini and Hochberg 1995). For protein analysis, the protein sequences of all genome features designated as “ORF” were downloaded from YeastMine (Balakrishnan et al. 2012), uploaded in batches into the IUPred2A tool (Mészáros et al. 2018), and the median score was computed for each protein. Distributions of median scores were compared using MWU with Benjamini and Hochberg (Benjamini and Hochberg 1995) correction for multiple testing.
Genetic interaction analysis
A single copy of each RBP was deleted from diploid HO+ parental strain YPS1009 that was hemizygous for SSD1 and harbored three copies of chromosome 12. The resulting double hemizygous strains (SSD1/ssd1Δ::KanMX; RBP/rbpΔ::HygMX) were sporulated and dissected to produce evenly spaced colonies of all desired genotypes (average of 22 colonies per genotype). Cells were grown on YPD plates for 3 days at 30°C, imaged, scaled by 200%, and colony size was quantified using the R package gitter v 1.1 (Wagih and Parts 2014) using a reference image (see R package documentation) and plate.format = c(4,8), with otherwise default parameters. Colony genotype was subsequently determined based on the segregation of drug markers (G418 resistance indicating ssd1Δ::KANMX or hygromycin resistance indicating rbpΔ::HYGMX). Flow cytometry was used to assess aneuploidy (described below). Tetrads with missing colonies or aberrant segregation patterns were excluded from downstream analysis.
The presence of synthetic genetic interactions was assessed as follows: for each independent dissection batch, the distribution of colony sizes was determined for each genotype. Genotype means were used to calculate defects for single mutants and aneuploids relative to wild-type controls. These defects were then used to calculate the distribution of expected colony sizes using a combinatorial model. For example, to generate the distribution of expected colony sizes for a given RBP mutant aneuploid, we first defined the defect due to Chr12 duplication using the mean size of the wild-type Chr12 colonies versus the mean size of wild-type euploid colonies, for colonies measured in the same batch. We then multiplied this defect by the observed size of each RBP mutant in the absence of aneuploidy, again for colonies in the same batch. The results generated a distribution of expected colony sizes that represent the expected combinatorial effect of aneuploidy and of RBP deletion in a euploid background. We then identified a statistically significant genetic interaction if the observed rbpΔ aneuploid colony sizes were greater, or smaller, than the expected colony sizes. A similar procedure was performed for other comparisons, estimating expected growth defects for each ssd1Δ rbpΔ euploid (ssd1Δ euploid defect × rbpΔ euploid defect) and ssd1Δ rbpΔ aneuploid (ssd1Δ aneuploid defect × rbpΔ aneuploid defect). Observed versus expected colony sizes were compared using a two-sided independent t-test with Benjamini–Hochberg FDR correction (Benjamini and Hochberg 1995).
Synthetic genetic interactions were assessed from liquid growth rates using a similar procedure. Growth rates were measured in at least biological triplicate for each strain, with strains being compared grown side-by-side in each replicate to enable replicate-paired t-tests to assess statistical significance.
Flow cytometry to assess aneuploidy
To determine euploid/aneuploid status, a portion of each colony was scraped, resuspended in 70% EtOH and stored at 4°. To process for flow cytometry, cells were incubated in 50 µg/mL RNase A in 0.05 M sodium citrate for at least 3 h or overnight, then incubated with 50 µg/mL proteinase K in 50 mM TRIS pH 8.0 and 10 mM CaCl2 for at least 3 h or overnight. Processed cells were stained with 1 µM Sytox Green for 15 min at room temperature, then run at a medium flow rate on Guava EasyCyte flow cytometer (Millipore). We defined differences in Sytox Green signal for known euploid and aneuploid control colonies. Aneuploidy genotyping was then done using these guidelines to assess all colonies of a given genotype from the same plate.
Protein–protein interaction network and enrichments
Interaction data were obtained from BioGRID via YeastMine as above and visualized using Cytoscape v. 3.9.1 (Shannon et al. 2003). Beneficial gene duplications were taken from Dutcher et al. (2024), restricted to the 316 genes that were especially beneficial in Chr12 aneuploids. We identified the subset of genes related to RNA functions, starting with genes mapped to the following GO SLIM process terms: mRNA processing, rRNA processing, RNA splicing, regulation of translation, cytoplasmic translation, translational elongation, RNA modification, sno(s)RNA processing, tRNA processing, mitochondrial translation, and tRNA aminoacylation for protein translation, according to the Saccharomyces Gene Database (Cherry et al. 2012). We then manually curated annotations and added genes with related functions that were missing from these categories, then simplified categorization for visualization purposes. This process yielded the 67 genes depicted in Figure 5A (peripheral nodes plus DHH1). All genes shown in Figure 5A, including the RBPs with significant genetic interactions (central nodes), were then analyzed to determine if they had more interactions with central nodes than expected by chance as follows: first, we identified the subset of encoded proteins that interacted with at least two central node RBPs (see Fig. 5A). For each protein that met this criterion, overrepresentation of interactions was then assessed using a hypergeometric test in R: phyper(x, m, n, k, lower.tail = FALSE), where x = the number of central node interactions for the given protein; m = the total number of interactors for that protein; n = 6400 (estimated size of total pool of interactors) − m; and k = 11 (number of central RBP nodes). Benjamini–Hochberg correction was then used to calculate FDR, taking FDRs < 0.05 as significant. Functional enrichments were assessed using the hypergeometric function and GO annotations defined in setRank version 1.0 (Simillion et al. 2017). Genes shown in network figures were manually categorized according to their primary functions, as listed in the key. Of the 286 Ssd1 RNA targets identified in Hose et al. (2020), 57 transcripts with uORFs were identified using data from Ingolia et al. (2009), combining a list of predicted uORFs with translated uORFs identified in that study. Analysis of the secondary structure of Ssd1-bound RNAs compared to the whole transcriptome was conducted using numerical scores representing secondary structure from Kertesz et al. (2010). Distributions of all available scores corresponding to the CDSs of Ssd1-bound mRNAs from Hose et al. (2020) were compared to the total available CDS scores of the yeast transcriptome using a two-sided MWU test. Ssd1 target overlap with clusters identified in Kershaw et al. (2023) was determined using a hypergeometric test: 96 of 267 Ssd1 targets (Hose et al. 2020) present in the data set are in “Cluster 3,” out of a total of 5050 RNAs across all clusters (Kershaw et al. 2023).
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
This work was funded by National Institutes of Health (NIH) grants R01CA229532 and R01GM148975 to A.P.G. H.A.D. was supported by training grants T32GM007133 and T32HG002760 to the Genomic Sciences Training Program.
Footnotes
-
Handling editor: John Woolford
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080199.124.
- Received July 19, 2024.
- Accepted October 15, 2024.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Auguste Dutcher is the first author of this paper, “Investigating the role of RNA-binding protein Ssd1 in aneuploidy tolerance through network analysis.” Auguste completed this work while a graduate student in Audrey Gasch's lab at the University of Wisconsin-Madison. Her paper investigates the RNA-binding network of Ssd1, a protein implicated in aneuploidy tolerance in wild yeast.
What are the major results described in your paper and how do they impact this branch of the field?
Our previous work showed that an RNA-binding protein (RBP) called Ssd1 is a key mediator of aneuploidy tolerance in a wild strain of budding yeast. In this paper, we interrogated a broader network by deleting other RBPs that share RNA targets with Ssd1 in cells carrying an extra copy of Chr12. We integrated these results with data from a global gene duplication screen that included other chromosome duplications, and ultimately identified an influential subgroup of proteins with shared functions, including translational repression and P-body formation. Our results suggest that these functions may play a role in aneuploidy tolerance, highlighting the roles of specific RBPs in either exacerbating or alleviating sensitivity to chromosome duplication.
What led you to study RNA or this aspect of RNA science?
I entered graduate school with a strong interest in RNA biology, having spent several years studying the evolution of small noncoding RNAs in bacteria. When I learned that studying an RNA-binding protein could help us understand how cells tolerate extra chromosomes, I was immediately drawn to the project.
What are some of the landmark moments that provoked your interest in science or your development as a scientist?
I never expected to become a biologist. Honestly, my interest in the field came partially from selfish motivations to avoid boredom. Scrolling through job postings after moving to a new city, I felt uninspired by many of the roles that aligned with my formal qualifications. I wanted autonomous, curiosity-driven work that was going to be intensely challenging and constantly evolving. Fortunately, I knew I liked research—my favorite role to date had been on a team studying the geography of health disparities, following my training as a social scientist in college. That experience instilled in me somewhat of a bias against deterministic thinking: Thorough investigation often revealed that people's choices were far more complex (and shaped by many more variables) than initially assumed.
Around the time I was considering a new career, advances in next-gen sequencing were propelling the study of the human microbiome into the popular science spotlight. I thought, “I've studied human communities, why not microbial ones instead?” Though my research interests evolved over time, it was that idea that ultimately put me back into college classrooms as a returning student, in front of a computer as a novice programmer, and eventually at the bench too. As in my previous research experience, I was humbled by both the immense body of knowledge generated by scientists before me and the unfathomable magnitude of what we still have to learn. The systems that surround us—whether in our cities or in our cells—are incredibly intricate. While this complexity makes our work challenging, it's also what makes it fascinating. To me, being a scientist is a commitment to a lifetime of continuous learning and growth: If we're not constantly reevaluating our ideas, reshaping our questions, and asking better ones based on what we have learned, we aren't doing it right.
What are your subsequent near- or long-term career plans?
I defended my dissertation in June 2024 and recently accepted a position in wastewater surveillance, where I'll leverage sequencing approaches to improve monitoring for pathogens and emerging antimicrobial resistance. I'm eager to apply my experience in both social and biological sciences to benefit public health, and I look forward to the challenges and opportunities in this rapidly growing field.








