
Decoding Argonaute specificity: insights from C. elegans and beyond
- Department of Biological Sciences, University of Southern California, Los Angeles, California 90089-2910, USA
- Corresponding author: cphil{at}usc.edu
Abstract
Small noncoding RNAs are versatile regulators of gene expression, capable of guiding the silencing of complementary mRNAs through their association with Argonaute proteins. This RNA-guided silencing, known as RNA interference (RNAi), is a conserved mechanism that shapes diverse biological processes, from development to genome defense. Central to the effectiveness of RNAi is the precise loading of small RNAs into their appropriate Argonaute partners, a step that ensures both specificity and fidelity in target recognition. Although most organisms harbor multiple classes of small RNAs and a corresponding repertoire of Argonautes, the rules that dictate their selective pairing remain only partially understood. Caenorhabditis elegans, with its expanded array of small RNA classes and Argonaute proteins, provides a powerful system to probe these mechanisms. In this review, we synthesize current knowledge on small RNA loading specificity, integrating insights from C. elegans with findings from other organisms. We focus on the interplay between small RNA biogenesis, biochemical properties of small RNAs, structural features of Argonautes, post-translational modifications, and the spatiotemporal coexpression patterns that together orchestrate precise Argonaute loading.
Keywords
SMALL RNAs
Small RNAs and the RNA interference pathway
RNA interference (RNAi) is a conserved gene-regulatory process mediated by small noncoding RNAs (ncRNAs) and their binding partners, Argonaute (AGO) proteins. Argonaute proteins recognize and bind transcripts that are complementary to the small RNA sequence, leading to transcript silencing. The first small RNA, the microRNA (miRNA) lin-4, and the phenomenon of RNAi were initially discovered in the nematode Caenorhabditis elegans, and later found to be conserved across diverse organisms, such as Arabidopsis thaliana, Drosophila melanogaster, Mus musculus, and Homo sapiens (Lee et al. 1993; Wightman et al. 1993; Fire et al. 1998; Pasquinelli et al. 2000; Agrawal et al. 2003).
Small RNAs can originate from both exogenous and endogenous sources. Exogenously introduced double-stranded RNA (dsRNA) is processed by the RNase III endonuclease Dicer into short dsRNA fragments, one strand of which is incorporated into the RNA-induced silencing complex (RISC), to mediate sequence-specific gene silencing (Bernstein et al. 2001; Ketting et al. 2001; Knight and Bass 2001; Schwarz et al. 2004). In the laboratory, exogenous dsRNA can be delivered into C. elegans by soaking, feeding, or direct injection into the gonad (Mello et al. 1991; Fire et al. 1998; Tabara et al. 1998), making RNAi a widely used tool for gene silencing and genetic screening.
Endogenous small RNAs play crucial roles in regulating gene expression, including silencing harmful transcripts and fine-tuning normal gene activity. For example, during C. elegans embryogenesis, activation of a single miRNA, lsy-6, can direct neuronal fate and determine cellular physiology and function (Cochella and Hobert 2012). During gametogenesis, small interfering RNAs (siRNAs) silence spermatogenesis genes to ensure normal gamete development (Han et al. 2009; Cornes et al. 2022; Ortega et al. 2024). Understanding small RNA pathways, particularly how binding and silencing specificity are achieved in model organisms like C. elegans, is essential. These insights provide a conceptual framework for human biology, shedding light on mechanisms of gene regulation and small RNA-related diseases, and may ultimately guide the development of precise RNAi-based therapies, including targeted interventions in immunotherapy.
Classification of small RNAs
Our understanding of small RNA pathways in C. elegans has expanded substantially in recent years, driven by the advances in high-throughput sequencing and CRISPR-based genome editing (Fig. 1). Small RNAs in C. elegans can be broadly categorized as either exogenous or endogenous, with each group encompassing distinct biogenesis pathways and Argonaute associations.
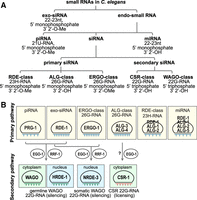
Classification of small RNAs and Argonaute proteins in C. elegans. (A) Small RNAs in C. elegans are categorized based on their Argonaute binding partners, biogenesis pathways, and molecular features including length, 5′ nucleotide identity, and terminal modifications. (B) Argonaute proteins in C. elegans are classified according to their associated small RNA partners, expression patterns, and molecular functions.
Exogenous siRNAs (exo-siRNAs) are 22–23 nt in length, possess 5′ monophosphate and 3′ 2′-O-methylation (2′-O-Me), and associate with the Argonaute protein RDE-1 (RNAi deficient-1) (Tabara et al. 1999; Billi et al. 2012; Svendsen et al. 2019). The catalytic slicing activity of RDE-1 is essential for efficient unwinding of the exo-siRNA duplex (Steiner et al. 2009). Endogenous small RNAs (endo-sRNAs) include microRNAs (miRNAs), PIWI-interacting RNAs (piRNAs), and endogenous small interfering RNAs (endo-siRNAs).
miRNAs are 21–23 nt single-stranded RNAs transcribed by RNA polymerase II (Pol II) (Lee et al. 2004; Ha and Kim 2014; Czech and Hannon 2011; Treiber et al. 2019). Primary miRNAs (pri-miRNAs) are processed by the Microprocessor complex, comprised of Drosha and DGCR8/Pasha, into precursor miRNAs (pre-miRNAs), which are then cleaved by Dicer in the cytoplasm into ∼22 nt duplexes with 2 nt 3′ overhangs (Lee et al. 2003; Denli et al. 2004). The guide strand is loaded onto Argonaute proteins (ALG-1, ALG-2, ALG-5, and RDE-1 in C. elegans) to mediate post-transcriptional gene regulation (Grishok et al. 2001; Corrêa et al. 2010; Brown et al. 2017).
piRNAs (21U-RNAs) are 21 nt long with a strong 5′ uridine bias, a 5′ monophosphate and 3′ 2′-O-Me (Ruby et al. 2006; Batista et al. 2008; Das et al. 2008; Billi et al. 2012; Kamminga et al. 2012; Montgomery et al. 2012). They associate with the PIWI Argonaute PRG-1 and recognize targets through partial complementarity (Batista et al. 2008; Wang and Reinke 2008; Bagijn et al. 2012; Lee et al. 2012). Type I piRNAs are transcribed from dedicated clusters on Chromosome IV, while Type II piRNAs arise from dispersed loci, including protein-coding genes (Ruby et al. 2006; Batista et al. 2008; Das et al. 2008; Gu et al. 2012). Maturation of piRNA precursors requires trimming of both the 5′ and 3′ ends and methylation by HENN-1 (Ruby et al. 2006; Billi et al. 2012; Kamminga et al. 2012; Montgomery et al. 2012; Tang et al. 2016; Podvalnaya et al. 2023).
26G-RNAs are a class of primary endo-siRNAs that are 26 nt in length with a 5′ guanine and 5′ monophosphate and are produced from mRNA in the antisense orientation by the RNA-dependent RNA polymerase (RdRP) RRF-3 and Dicer (Gent et al. 2009, 2010; Han et al. 2009; Conine et al. 2010). Their biogenesis depends on the ERI complex (e.g., DRH-3, ERI-5) (Thivierge et al. 2012; Almeida et al. 2018). 26G-RNAs can be classified into ERGO-class and ALG-3/4-class 26G-RNAs depending on their Argonaute partners. ERGO-class 26G-RNAs, which associate with ERGO-1 and are 2′-O-methylated, target retrotransposons, duplicated genes, pseudogenes, and long noncoding RNAs (lncRNAs) during oogenesis (Han et al. 2009; Gent et al. 2010; Vasale et al. 2010; Fischer et al. 2011; Billi et al. 2012; Kamminga et al. 2012; Montgomery et al. 2012; Seroussi et al. 2023). In contrast, ALG-3/4-class 26G-RNAs are not methylated and associate with ALG-3 and ALG-4 to target spermatogenesis genes (Conine et al. 2010, 2013; Montgomery et al. 2012; Almeida et al. 2019b).
23H-RNAs are 23 nt endogenous siRNAs with 5′ nucleotide preference for U, A, or C. They likely carry similar terminal modifications as exogenous siRNAs, including a 5′ monophosphate and 3′ 2′-O-methylation, and they exist at relatively low abundance in C. elegans. 23H-RNAs are produced primarily from long hairpin RNAs and repetitive elements through Dicer processing. They primarily associate with RDE-1, although ALG-1 and ALG-2 can also compete for binding (Knittel et al. 2024).
Secondary siRNAs (22G-RNAs) are generated by RdRPs from mRNA templates, typically acting downstream from exo-siRNA, piRNA, or 26G-RNA targeting (Sijen et al. 2001; Pak and Fire 2007). These 22G-RNAs are 22 nt long with a 5′ guanine, 5′ triphosphate, and 3′ hydroxyl group, and their mode of action depends on the identity of their Argonaute partners. WAGO-class 22G-RNAs are produced downstream from piRNAs and siRNAs and are loaded into worm-specific Argonautes (WAGOs). They primarily target pseudogenes, transposons, and other cryptic loci for silencing (Pak and Fire 2007). In contrast, CSR-class 22G-RNAs associate with CSR-1 and are linked to gene licensing or fine-tuning of protein-coding gene expression (Claycomb et al. 2009; Seth et al. 2013; Wedeles et al. 2013; Gerson-Gurwitz et al. 2016). Despite being largely biochemically indistinguishable, CSR- and WAGO-class 22G-RNAs differ by at least one detectable feature: A subset of CSR-1 22G-RNAs undergo 3′ uridylation by the terminal uridylyltransferase CDE-1, whereas WAGO-class 22G-RNAs generally lack this modification (Van Wolfswinkel et al. 2009). WAGO-class 22G-RNAs are synthesized by the RdRPs EGO-1 and RRF-1, while CSR-class 22G-RNAs rely exclusively on EGO-1 (Aoki et al. 2007; Claycomb et al. 2009; Gu et al. 2009). The mechanisms governing sorting of small RNAs between CSR-1 and WAGO Argonautes remain incompletely understood.
Although 22G-RNAs are unique to nematodes, many classes of small RNAs, including miRNAs, piRNAs, and siRNAs, are conserved across diverse organisms, each defined by distinct biogenesis pathways and Argonaute associations. In D. melanogaster, these three major classes parallel those in C. elegans and are likewise sorted into specialized Argonautes. Notably, the piRNA amplification “ping-pong” cycle in fly germ cells functionally resembles the 22G-RNA amplification system in C. elegans (Brennecke et al. 2007; Czech and Hannon 2016). Such cross-species comparisons highlight both the conservation and innovation within small RNA pathways, illustrating how diverse molecular strategies converge on the shared goal of achieving precise gene regulation. Understanding these parallels and differences provides a framework for uncovering the general principles of Argonaute–small RNA interactions, which will be explored in sections that follow.
ARGONAUTE PROTEINS
Structure and classification
Small RNAs alone are insufficient to mediate gene silencing and are unstable due to their lack of a 5′ cap and 3′ poly(A) tail (Ketting and Cochella 2021). To achieve gene silencing, small RNAs must bind to Argonaute proteins, forming a stable RISC that targets complementary transcripts. Similarly, unloaded Argonautes are prone to proteasomal and autophagic degradation (Johnston et al. 2010; Derrien et al. 2012; Martinez and Gregory 2013; Smibert et al. 2013; Kobayashi et al. 2019). This interdependence emphasizes the mutual stability provided by Argonaute–small RNA interactions, and that the Argonaute–small RNA complexes, rather than either component alone, are the functional units of silencing (Jannot et al. 2008).
Argonaute proteins have diversified in function, ranging from prokaryotic immunity systems that target mobile DNA via DNA interference (DNAi) (Swarts et al. 2014) to roles in DNA replication and repair (Sheng et al. 2014; Lisitskaya et al. 2022) and to guiding RNAi and directing DNA and histone methylation in eukaryotes (Joh et al. 2014; Adiliaghdam et al. 2020; Sigman et al. 2021). Despite this broad functional range, their structural architecture remains remarkably conserved across evolution, particularly within the seed-binding region that anchors guide RNAs (Wallmann and Van de Pette 2025). Importantly, this nucleic acid–binding interface is also a hotspot for disease-associated mutations (Wallmann and Van de Pette 2025), underscoring the critical importance of accurate Argonaute–small RNA loading.
Argonaute proteins are defined by four conserved structural domains: N, PAZ, MID, and PIWI domains (Fig. 2A). These domains are organized into two lobes, N-PAZ and MID–PIWI, connected by flexible linkers (Song et al. 2004; Elkayam et al. 2012; Nakanishi et al. 2012; Schirle and MacRae 2012). The PAZ and MID domains coordinate binding to the 3′ and 5′ ends of the small RNA, respectively. The PIWI domain resembles RNase H, and in some Argonautes, confers endonucleolytic (“slicer”) activity through a conserved DEDD/H motif. Other Argonautes lacking this catalytic activity recruit cofactors for mRNA silencing through degradation or translational repression (Eulalio et al. 2009; Iwakawa and Tomari 2013; Chen et al. 2014). The N domain is less well characterized but plays a role in small RNA duplex unwinding and loading during RISC assembly (Kwak and Tomari 2012).
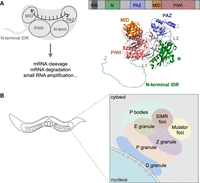
Overview of Argonaute proteins and germ granules in the C. elegans germline. (A) Domain architecture and functions of Argonaute proteins. Argonaute proteins contain four conserved domains: the N, PAZ, MID, and PIWI domains, along with an unstructured N-terminal extension. The MID and PAZ domains anchor the 5′ and 3′ ends of small RNAs, respectively. The structure of C. elegans HRDE-1 is shown (right), with domains assigned by alignment to human AGO2. (B) C. elegans germ granules are composed of multiple compartments that are spatially organized within the perinuclear structure, each supporting distinct aspects of RNA regulation.
In addition to the four conserved structural domains, Argonaute proteins also contain less well-defined regions that contribute critically to their regulation. The N-terminal extension (NTE) is an intrinsically disordered region (IDR) that varies widely in length and sequence, not only across organisms but also among different Argonaute proteins within the same species. Despite lacking stable structure, this N-terminal IDR performs several important regulatory functions. For example, in nuclear Argonautes, it can encode nuclear localization (NLS) and nuclear export (NES) signals that regulate nucleocytoplasmic shuttling (Guang et al. 2008; Ye et al. 2012; Bologna et al. 2018; Yashiro et al. 2018; Chen and Phillips 2024), and it can also influence small RNA loading efficiency and specificity (Xu et al. 2023).
Immediately following the IDR is a short, structured linear element known as the N-coil (∼14 amino acids in Arabidopsis AGO1 and human AGO2), which connects the N domain to the MID–PIWI lobe (Poulsen et al. 2013). Recent studies show that the N-coil functions as a conformational switch distinguishing unloaded from loaded Argonaute proteins: In the unloaded state, the N-coil is exposed and serves as a platform for recognition by the autophagy cargo receptors, targeting the Argonaute for degradation (Bressendorff et al. 2023, 2025). Although this mechanism has not been tested in C. elegans, the N-coil may offer a useful framework for understanding why only a subset of C. elegans Argonautes are destabilized when unloaded, while others remain stable (Zhao et al. 2025). These findings highlight how structural elements beyond the conserved domains can function as conformational checkpoints, offering new insight into how Argonautes are monitored and maintained.
In C. elegans, 26 Argonaute genes encode 19 functional Argonaute proteins (Table 1). These proteins can be classified into three clades: the AGO clade, PIWI clade, and WAGO (worm-specific Argonaute) clade. Members of each clade bind subsets of small RNAs with distinct features (Yigit et al. 2006; Seroussi et al. 2023).
Summary of Argonaute proteins and their loaded small RNAs
In other model organisms, Argonaute repertoires are typically smaller but still show functional diversification. D. melanogaster has five Argonaute proteins, Piwi, Aubergine, Ago1, Ago2, and Ago3. The PIWI clade, Piwi, Aubergine, and Ago3, binds to piRNAs (Saito et al. 2006; Vagin et al. 2006), while the AGO clade, Ago1 and Ago2, binds to miRNAs and siRNAs, respectively (Hammond et al. 2001; Okamura et al. 2004; Förstemann et al. 2007). Small RNA sorting depends on multiple factors, which will be discussed in more detail later in this review (Förstemann et al. 2007; Tomari et al. 2007). A. thaliana encodes 10 Argonaute proteins, divided into three clades based on sequence and functional characteristics (Vaucheret 2008). Clade I (AGO1, AGO5, and AGO10) is primarily involved in post-transcriptional gene silencing (PTGS) in the cytoplasm (Bohmert et al. 1998; Lynn et al. 1999; Morel et al. 2002; Vaucheret 2008). Clade II (AGO2, AGO3, and AGO7) functions in DNA repair, antiviral defense, and endogenous RNAi in both the nucleus and cytoplasm (Adenot et al. 2006; Vaucheret 2008). Clade III (AGO4, AGO6, AGO8, and AGO9) is primarily nuclear and mediates transcriptional silencing (TGS) via siRNAs (Zilberman et al. 2003; Zheng et al. 2007; Vaucheret 2008). In humans, four Argonaute proteins (AGO1–4) bind both miRNAs and siRNAs (Petri et al. 2011; Dueck et al. 2012); only AGO2 retains slicer activity (Liu et al. 2004; Meister et al. 2004). The four PIWI proteins (PIWIL1–4) are involved in piRNA-mediated gene silencing (Cox et al. 1998; Qiao et al. 2002). The mechanisms determining small RNA sorting in humans remain elusive.
Overall, Argonautes are a conserved class of nucleic acid–binding proteins whose core structure is maintained across evolution, yet has diversified to accommodate distinct small RNA classes and gene-silencing mechanisms in different organisms. Recent advances in AlphaFold-based structure modeling provide unprecedented opportunities to link Argonaute structure to the molecular mechanisms that underlie small RNA loading, target recognition, and gene-regulatory specificity.
Argonaute-mediated regulation
Once the appropriate small RNAs are loaded into Argonaute proteins, they form functional RNA-induced silencing complexes (RISCs) that exert diverse downstream regulatory effects. Argonautes bound to miRNAs typically promote translational repression and transcript degradation. miRNAs usually recognize sequences within the 3′ untranslated region (3′UTR) of target mRNAs, where they recruit complexes, including deadenylation and decapping enzymes, resulting in translational inhibition and mRNA decay (Duchaine and Fabian 2019). Notably, miRNAs do not require perfect complementarity to their targets. Instead, the degree of base-pairing, particularly in the seed region (nucleotides 2–8 or 2–9 from the 5′ end), influences the silencing outcome (Schirle et al. 2014; Salomon et al. 2015; Duan et al. 2022). In D. melanogaster, for example, perfect seed pairing often leads to transcript degradation, while imperfect pairing typically results in translational repression (Ameres et al. 2011; Fuchs Wightman et al. 2018; Pinhal et al. 2025).
In plants, however, miRNA-directed silencing operates through a distinct logic. Plant miRNAs primarily recognize protein-coding sequences (CDSs) rather than 3′ UTRs and generally exhibit near-perfect complementarity to their targets, which promotes direct mRNA cleavage (Jones-Rhoades and Bartel 2004; Millar and Waterhouse 2005). This contrast between animal and plant miRNA targeting highlights how the same fundamental Argonaute–miRNA machinery can be adapted across lineages to produce distinct regulatory modes.
In C. elegans, Argonautes loaded with primary siRNAs and piRNAs act primarily by initiating the production of secondary 22G-RNAs. These primary Argonautes recognize targets with some tolerance for mismatches, particularly outside the seed region (Rual et al. 2007; Bagijn et al. 2012; Lee et al. 2012; Montgomery et al. 2012; Zhang et al. 2018). Upon target engagement, they recruit RdRPs to synthesize 22G-RNAs directly from the target transcripts, yielding siRNAs with perfect complementarity to their mRNA templates. Once produced, these secondary 22G-RNAs are loaded into downstream Argonautes that execute distinct regulatory outcomes.
Secondary siRNA-bound Argonautes fall into two major functional categories: silencing and licensing. WAGO-class Argonautes mediate post-transcriptional silencing via RNA degradation or translational repression in the cytoplasm, or cotranscriptional gene silencing in the nucleus. In the nucleus, WAGOs such as NRDE-3 (in somatic cells) and HRDE-1 (in the germline) recruit chromatin-modifying enzymes that deposit repressive histone marks, notably H3K9me3, at target loci (Guang et al. 2008; Buckley et al. 2012). This process, known as nuclear RNAi, contributes to transcriptional silencing and epigenetic memory (Buckley et al. 2012).
Conversely, the Argonaute CSR-1 promotes gene licensing rather than silencing (Claycomb et al. 2009). CSR-1 tethering in vivo can safeguard transcripts from silencing and even reactivate previously silenced transcripts (Wedeles et al. 2013). Endogenously, CSR-1 protects germline-expressed genes from piRNA-mediated silencing, a role thought to involve counteracting PRG-1 recognition, preventing recruitment of repressive chromatin modifiers, and promoting sense transcription by RNA polymerase II (Claycomb et al. 2009; Seth et al. 2013; Cecere et al. 2014). Through these activities, CSR-1 ensures robust germline expression and fertility.
Together, these downstream effects highlight the versatility of Argonaute proteins as effectors of both gene repression and gene expression, dictated by the type of small RNA they are loaded with and the context of their action—cytoplasmic or nuclear, post-transcriptional or epigenetic.
RNAi pathway components are organized in compartmentalized granules
The RNAi pathway is a multistep gene-silencing mechanism in which many of its components are spatially organized within compartmentalized perinuclear germ granules. These granules are phase-separated biomolecular condensates, typically formed through weak, multivalent interactions between proteins that contain IDRs (Brangwynne et al. 2009; Peran and Mittag 2020; Vazquez et al. 2022; Zheng and Zhang 2024). This physical organization allows for efficient coordination of the molecular events required for small RNA production, Argonaute loading, and gene silencing.
In C. elegans, at least six distinct subdomains of the germ granule have been identified: P granules, Z granules, Mutator foci, SIMR foci, D granules, and E granules (P, Z, M, S, D, and E compartments) (see review by Phillips and Updike 2022; Fig. 2B). P granules, the first germ granule compartment to be discovered in C. elegans, can be identified by the presence of PGL-1 and PGL-3 and play key roles in RNA storage, processing, and surveillance (Brangwynne et al. 2009; Updike and Strome 2010; Hanazawa et al. 2011). Z granules, defined by the presence of ZNFX-1 and WAGO-4, are essential for the inheritance of gene-silencing signals across generations (Wan et al. 2018). Mutator foci, scaffolded by the proline-, glutamine-, and asparagine-rich protein MUT-16, serve as sites for secondary small RNA amplification by recruiting key components of the small RNA biogenesis machinery (Phillips et al. 2012). SIMR foci, characterized by SIMR-1, are found adjacent to Mutator foci and promote piRNA-dependent secondary small RNA production and loading of small RNAs into nuclear Argonaute proteins (Manage et al. 2020; Chen and Phillips 2024, 2025). Recently, D granules, containing DDX-19 and CSR-1, were found to be between nuclear pore complexes and P granules, perhaps serving as sites where CSR-1 licenses germline mRNAs prior to screening by piRNAs and siRNAs in the P and Z granules (Sheth et al. 2010; Huang et al. 2025). E granules, another recently described germ granule compartment, are marked by EGO-1, EGC-1, and ELLI-1 and are specifically required for the biogenesis of a subset of CSR-class 22G-RNAs, particularly at the 5′ ends of target transcripts (Chen et al. 2024b; Li et al. 2025). Lastly, processing bodies (P bodies), well-known condensates involved in mRNA turnover and storage of translationally repressed transcripts, are also closely associated with germ granules and have been linked to transgenerational gene silencing (Du et al. 2023).
Germ granules undergo extensive reorganization during C. elegans embryogenesis. P granules were first observed to accumulate selectively in germline blastomeres, a pattern maintained by autophagy-mediated clearance of somatic P granules (Strome and Wood 1982; Zhang et al. 2009). More recent studies reveal that multiple germ granule subtypes display dynamic developmental localization patterns, including the formation of transient somatic counterparts during embryogenesis. For example, in early embryos, P granules and Z granules initially form a merged compartment that separates only when the Z2/Z3 germ cells are specified and germline transcription begins (Seydoux and Strome 1999; Wan et al. 2018). Mutator and SIMR foci undergo similar partitioning during this developmental window (Chen and Phillips 2025), highlighting that granule remodeling is tightly coupled to transcriptional onset. Additional studies have revealed specialized somatic granules marked by distinct RNAi pathway components. These include somatic granules containing CSR-1, SIMR-1, RDE-12, RSD-6, and ZNFX-1 (Shirayama et al. 2014; Yang et al. 2014; Wan et al. 2018; Ouyang et al. 2019; Chen and Phillips 2025). Like P granules, CSR-1, SIMR-1, and ZNFX-1 somatic granules are regulated by autophagy (Chen and Phillips 2025), emphasizing the importance of precise spatiotemporal granule specification in coordinating gene-regulatory programs during embryogenesis.
Germ granules also play important roles in gene regulation in other organisms (Thomas et al. 2023). For example, in D. melanogaster, germ granules form during oogenesis and accumulate in embryonic germ cells, where they are essential for germ cell specification. Mutations in core components such as oskar and tudor disrupt RNA localization within these granules and impair proper germline formation (Trcek et al. 2015; Trcek and Lehmann 2019). These examples highlight the conserved importance of phase-separated RNA–protein granules in germline development and small RNA-mediated regulation.
Taken together, the C. elegans germline provides an unusually rich and complex environment for studying small RNA pathways, with an expanded repertoire of Argonaute proteins and noncoding RNAs spatially organized across distinct germ granule domains. This organization facilitates precise regulation of gene expression during development and inheritance. In the following sections, we will explore how Argonaute proteins recognize and bind to their corresponding small RNA targets to execute silencing or licensing functions.
ARGONAUTE–SMALL RNA BINDING SPECIFICITY AND ITS REGULATION
Precise binding between small RNAs and Argonaute proteins is essential for accurate gene regulation, and this specificity is determined by multiple interconnected factors (Fig. 3). First, the biochemical characteristics of the small RNA, shaped by its mode of biogenesis and subsequent processing, play a central role in Argonaute selection. Second, the properties of the Argonaute proteins themselves, including their structural features, post-translational modifications, and interacting protein partners, contribute to small RNA binding specificity. Third, spatial and temporal patterns of both the small RNAs and Argonautes must be appropriately aligned; they must be expressed in the correct tissues, at the right time, and in the correct stoichiometric balance. No single factor is sufficient to fully explain Argonaute specificity; rather, it is the coordinated integration of all of these layers that ensures accurate small RNA loading and downstream gene-regulatory effects.
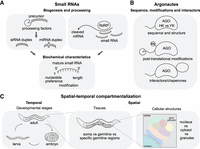
Mechanisms underlying Argonaute–small RNA loading specificity. (A) Biogenesis factors and biochemical features of small RNAs influence their selective loading into specific Argonaute proteins. During biogenesis, precursor structure, the use of particular RNA-dependent RNA polymerases, and the choice of duplex strand shape the properties of mature small RNAs. These properties, including length, 5′ nucleotide preference, and terminal modifications, collectively affect Argonaute loading specificity. (B) Post-translational modifications of Argonautes, together with interactions with accessory proteins, further refine small RNA binding specificity. (C) Spatial and temporal coexpression of small RNAs and their Argonaute partners constrains loading specificity. Effective pairing requires coexpression in developmental stage, tissue, and subcellular compartment, including the nucleus, cytoplasm, or specialized germ granules.
SMALL RNAs AND THEIR BIOCHEMICAL CHARACTERISTICS
Small RNAs carry distinct biochemical signatures that determine their incorporation into specific Argonaute proteins. The importance of small RNA biochemical signatures in sorting is clearly demonstrated in multiple systems. Early studies in Drosophila and human cell lines suggest that the guide strand selection is influenced by thermodynamic asymmetry: The strand with its 5′ end at the thermodynamically less stable terminus of the duplex is preferentially selected as the guide and loaded into an Argonaute protein (Khvorova et al. 2003; Schwarz et al. 2003). Subsequent work in A. thaliana revealed that the 5′ nucleotide is another dominant determinant of sorting. AGO1 preferentially binds small RNAs with a 5′ uridine, AGO2 and AGO4 favor 5′ adenosine, and AGO5 selects for those with 5′ cytosine. Remarkably, altering the 5′ nucleotide of a small RNA can redirect it to a different Argonaute, thereby changing its regulatory function (Kim 2008; Mi et al. 2008; Montgomery et al. 2008; Takeda et al. 2008). These findings demonstrate how biochemical features of small RNAs dictate their functional outcomes.
Small RNA length also influences Argonaute sorting in Arabidopsis. AGO1 and AGO2 typically bind 21 nt small RNAs, including most miRNAs and some endogenous siRNAs. AGO6 also incorporates short 21–22 nt siRNAs derived from transposable elements to mediate RNA-directed DNA methylation (McCue et al. 2015). In contrast, AGO4 binds 24 nt small RNAs, which often participate in RNA-directed DNA methylation and chromatin modification. AGO5 is less selective for length, and can bind small RNAs of various sizes, provided they carry the preferred 5′ cytosine (Mi et al. 2008; Zha et al. 2012). Consistent with these observations, in vitro RISC assembly assays incubating AGO4 and AGO6 with siRNAs of varying lengths (19–25 nt) demonstrated distinct binding preferences: Both AGO4 and AGO6 bind long siRNAs (23–25 nt), with highest efficiency for 24 nt siRNAs, while AGO6 also retains the capacity to bind shorter 21–22 nt siRNAs, indicating a broader length tolerance (Liu et al. 2022).
Similarly, the biochemical properties of small RNAs influence the distinct binding patterns of Drosophila Ago1 and Ago2: Ago1 almost exclusively associates with miRNAs, whereas Ago2 predominantly binds siRNAs. Distinct 5′ biases have been observed: Ago1-bound small RNAs are highly enriched for 5′ U, while Ago2-bound small RNAs generally lack this preference (Czech et al. 2008). Moreover, certain subsets of Ago2-associated RNAs exhibit a 5′ C bias (Ghildiyal et al. 2008). This differential sorting between siRNAs and miRNAs is mediated by the chaperone R2D2 (discussed later) and further influenced by small RNA sequence preferences. An in vitro randomized small RNA loading assay demonstrated that Ago2 favors G-rich sequences, whereas Ago1 preferentially loads small RNAs enriched in GAC nucleotides near the 5′ end. These preferences are also observed in vivo, suggesting that Argonaute loading is shaped not only by the canonical 5′ U bias but also by additional sequence motifs (Goh and Okamura 2019).
Interestingly, although Drosophila Ago2 is considered a canonical siRNA effector, it can also load the passenger (miRNA*) strand of miRNA duplexes (Czech et al. 2009; Okamura et al. 2009). Normally, the miRNA* strand is degraded during biogenesis, but it can instead be incorporated into Ago2 through the canonical siRNA loading machinery. This unusual sorting between miRNA and miRNA* relies on distinct biochemical features. Specifically, Ago2 preferentially loads miRNA* with 3′ 2′-O-Me and favors base-pairing at positions 9 and 10, whereas unpaired or G:U wobble at these positions correlates with Ago1 loading. These findings in Arabidopsis and Drosophila highlight how small RNA features, such as 5′ nucleotide identity, length, modifications, and sequence context, act as powerful determinants of Argonaute specificity, a principle likely conserved but adapted in other organisms, including C. elegans.
In C. elegans, primary small RNAs (e.g., miRNAs, piRNAs, exo-siRNAs, 26G-RNAs) have distinct biochemical characteristics, including length, 5′ nucleotide identity, and 3′ modification, which may influence their sorting into different Argonaute proteins, although this has not been tested directly (Fig. 3A). In contrast, secondary siRNAs generated by RdRPs are highly uniform: Although different classes show distinct patterns of distribution along their target transcripts, they remain consistently 22 nt in length, strongly 5′ guanine-biased, and carry a 5′ triphosphate group (Ambros et al. 2003; Gu et al. 2009; Ishidate et al. 2018; Singh et al. 2021; Seroussi et al. 2023). Nonetheless, WAGO-class and CSR-class 22G-RNAs are generally sorted into the correct Argonaute proteins, although some degree of cross-loading can occur. For example, WAGO-4 binds to both WAGO- and CSR-class 22G-RNAs under wild-type conditions (Seroussi et al. 2023), while WAGO-1 and HRDE-1, which normally associate with WAGO-class 22G-RNAs, can misload CSR-class 22G-RNAs in glh-1 and hrde-2 mutants, respectively. Similarly, CSR-1 isoforms normally bind distinct subsets of small RNAs, but this specificity can be altered by the loss of arginine methylation (Nguyen and Phillips 2021). These observations indicate that biochemical features alone are insufficient to explain sorting specificity, pointing to additional regulatory layers that act alongside small RNA structure.
In addition to 5′ nucleotide identity and length, the secondary structure of the small RNA duplex is a key determinant of Argonaute selection. In C. elegans, both ALG-1/2 and RDE-1 bind to small RNAs derived from dsRNA precursors, but they can discriminate based on structural features. Pre-miRNA duplexes, which contain mismatches and bulges, are preferentially loaded into ALG-1/2, whereas perfectly base-paired pre-siRNA duplexes are selectively loaded into RDE-1 (Steiner et al. 2007). This structural distinction underlies differential recognition by specific Argonautes and possibly chaperone proteins, paralleling the well-characterized loading preferences of Drosophila Ago1 and Ago2 described above. More broadly, structural elements such as G:U wobble pairs, internal mismatches, and terminal loops influence the thermodynamic properties of small RNA duplexes, thereby shaping their Argonaute loading fate (Tomari et al. 2007; Czech et al. 2009).
Together, these observations emphasize that the molecular features of small RNAs, including their sequence, chemical modifications, length, and precursor structure, are central determinants of Argonaute binding specificity.
ARGONAUTE PROPERTIES AND INTERACTIONS
Small RNA features contribute substantially to Argonaute loading, but they do not fully account for how distinct regulatory outcomes are achieved. For example, in C. elegans, biochemically indistinguishable 22G-RNAs can be partitioned between CSR-1 and WAGO proteins, producing opposite effects on gene expression. Such examples highlight the importance of Argonaute-encoded features and cofactor interactions in shaping loading specificity (Fig. 3B).
Sequence and structure of Argonaute proteins
The small RNA binding pockets of Argonaute proteins are central to their function. The 5′ end of the small RNA is anchored by the MID domain, while the 3′ end is recognized by the PAZ domain (Lingel et al. 2003; Yan et al. 2003; Song et al. 2004; Ma et al. 2005; Boland et al. 2010; Frank et al. 2010). Specific structural features within these binding pockets, together with neighboring domains, determine selective small RNA recognition.
Key mechanistic insight came from the crystal structure of an Archaeoglobus fulgidus Piwi protein bound to dsRNA (Ma et al. 2005). This study revealed that the phosphorylated 5′ end of the guide RNA is anchored within a basic pocket formed at the MID–PIWI interface, where the first nucleotide stacks against a conserved tyrosine residue in a sequence-independent manner. The small RNA binding pocket is highly conserved, particularly the YKQK motif (Tyr123, Lys127, Gln137, and Lys163). Mutational analysis showed that disrupting this motif, for example, by substituting two conserved residues in human AGO2, or in Drosophila Ago3, Aub, and Ago1, abolishes small RNA loading and downstream silencing (Ma et al. 2005; Djuranovic et al. 2010; Webster et al. 2015).
Interestingly, nematodes, including C. elegans, display a clear divergence of this pocket: While primary Argonautes retain the conserved YKQK motif, all secondary Argonautes (WAGOs) in clades III-V of the Nematoda phylum carry a divergent HKQK motif (Fig. 4; Parker and Barford 2006; Buck and Blaxter 2013). This divergence has been hypothesized to reflect a specialization for small RNAs with distinct 5′ phosphorylation states (Faehnle and Joshua-Tor 2007). Primary small RNAs, generated by Dicer, carry a 5′ monophosphate, whereas secondary 22G-RNAs generated by RdRPs carry a 5′ triphosphate and preferentially associate with WAGOs containing the HKQK pocket (Pak and Fire 2007). Supporting this idea, 5′ triphosphorylated RNAs, the EGO-1 and RRF-1 RdRPs that generate them, and WAGO Argonautes bearing the HKQK motif are all absent from Nematoda clades I and II, but present in clades III-V (Dalzell et al. 2011; Buck and Blaxter 2013; Sarkies et al. 2015; Tu et al. 2015; Almeida et al. 2019b). Consistent with the critical role of YKQK in other organisms, functional studies in C. elegans have shown that substituting the first two residues of the HKQK motif (HK→AA) disrupts small RNA binding, alters Argonaute localization, and abolishes silencing (Chen and Phillips 2024, 2025). Although the precise contribution of histidine versus tyrosine to determining binding specificity remains unresolved, these findings underscore that a precise structural match between Argonaute proteins and their small RNA partners is essential for recognition.
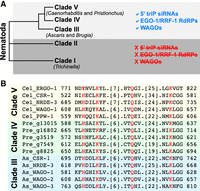
Conservation of the HKQK motif across Nematoda WAGO Argonautes. (A) Cladogram of the Nematoda phylum, with representative genera indicated. 5′ Triphosphorylated RNAs, EGO-1/RRF-1 RdRPs, and WAGO Argonautes are present in clades III-V but absent from clades I and II. (B) Protein sequence alignment of WAGO Argonautes from clade III (Ascaris suum), clade IV (Panagrellus redivivus), and clade V (C. elegans) shows conservation of the divergent HKQK motif in the MID domain. C. elegans ERGO-1, a primary AGO-clade Argonaute, is included for comparison and retains the canonical YKQK motif (Buck and Blaxter 2013; Zagoskin et al. 2022). Sequence alignment was generated using T-coffee (Notredame et al. 2000).
Additional crystallographic studies highlight the role of the MID domain in 5′ nucleotide recognition. In human AGO2, a rigid nucleotide specificity loop within the MID domain interacts directly with uridine or adenosine, explaining the 5′ nucleotide bias in miRNA loading (Frank et al. 2010). In Arabidopsis, AGO1, AGO2, and AGO5 each harbor distinct specificity loops that form precise side-chain interactions with the 5′ nucleotide, providing a direct structural mechanism for Argonaute sorting (Frank et al. 2012). This loop is conserved among miRNA-pathway Argonautes across organisms, including human AGO1-4, C. elegans ALG-1/2, and Drosophila Ago1 (Frank et al. 2010), underscoring its central role in 5′ nucleotide-mediated sorting. However, the specificity loop is not the sole determinant of 5′ nucleotide choice. Sequence preferences established during small RNA processing can further shape Argonaute loading. In Drosophila Piwi, for example, the loop disfavors 5′C piRNAs, but upstream processing steps are enriched for pyrimidines (U/C), resulting in the characteristic 5′U bias of mature piRNAs (Stein et al. 2019).
Beyond the MID domain, other regions also influence small RNA binding specificity. In Arabidopsis, AGO4 and AGO6, key players in RNA-directed DNA methylation (Zilberman et al. 2003; Chan et al. 2004; Zheng et al. 2007), both preferentially load 24 nt siRNAs with a 5′ adenine (Matzke and Mosher 2014). In vitro RISC assembly revealed subtle distinctions in their preference: AGO6 shows a strict requirement for 24A siRNAs, whereas AGO4 can accommodate other nucleotides with lower efficiency, with additional sensitivity to the thermodynamic stability of the siRNA duplex. Domain-swap experiments showed that cooperative contributions from both the MID and PIWI domains fine-tune 5′ nucleotide specificity (Liu et al. 2022).
Overall, studies across systems reveal that diverse structural features of Argonautes contribute uniquely to small RNA recognition, adding an additional layer of regulation beyond small RNA biochemistry.
Post-translational modifications of Argonaute proteins
Argonaute proteins are extensively regulated by post-translational modifications (PTMs), including methylation and N-terminal processing. These modifications influence Argonaute stability, localization, protein–protein interactions, and the efficiency and specificity of small RNA binding (Table 2).
Summary of Argonaute protein PTM
Arginine methylation, typically catalyzed by protein arginine methyltransferases (PRMTs) in arginine-glycine-rich (RGG/RG) motifs, also regulates Argonaute specificity (Thandapani et al. 2013; Blanc and Richard 2017). In C. elegans, CSR-1A, the spermatogenic-specific isoform of the Argonaute CSR-1, has an extended N-terminal exon enriched in RGG/RG motifs that undergo methylation. This modification helps CSR-1A predominantly bind spermatogenic 22G-RNAs; loss of the RGG/RG motifs broadens its small RNA repertoire to include oogenic 22G-RNAs (Nguyen and Phillips 2021). Similarly, in Arabidopsis, the AGO1 n-terminal extension is symmetrically dimethylated by PRMT5. Loss of methylation does not impair general miRNA binding but selectively affects loading of a subset of siRNAs (Martín-Merchán et al. 2024).
Methylation can also influence small RNA loading and specificity through indirect mechanisms. In Drosophila, methylation of the PIWI-clade Argonaute Aubergine (Aub) is closely linked to its role in piRNA processing. Although methylation is not required for the initial loading of piRNAs into Aub, piRNA loading triggers methylation by PRMT5. This post-loading modification facilitates Aub's interaction with Tudor domain–containing proteins, which is essential for initiating ping-pong amplification and expanding the piRNA pool (Huang et al. 2021).
N-terminal processing is another post-translational mechanism that regulates Argonaute–small RNA interactions. The unstructured N-terminal regions of Argonaute proteins remain one of the least well-understood regions, yet they contribute to small RNA loading in several organisms. In C. elegans, the dipeptidase DPF-3 cleaves the proline-rich N termini of WAGO-1 and WAGO-3. Loss of this processing in dpf-3 mutants reduces WAGO-1 specificity for WAGO-class 22G-RNAs, leading to inappropriate binding of CSR-class 22G-RNAs and defects in gene regulation and fertility (Gudipati et al. 2021). DPF-3 also interacts with ALG-1 and cleaves the N-terminal peptide of ALG-2 in vitro. Though a direct role in small RNA loading has not yet been established, in dpf-3 mutants, total miRNA levels are restored in alg-1 mutants, rescuing miRNA-related defects (Harvey et al. 2025). These findings underscore the importance of N-terminal processing in modulating Argonaute activity and small RNA specificity.
Phosphorylation is one of the most extensively studied post-translational modifications of Argonaute proteins and can influence their subcellular localization, small RNA binding, and RISC assembly. In human AGO2, phosphorylation at S387 within the linker region between PAZ and MID domains is required for proper P-body localization and translational repression (Horman et al. 2013), while phosphorylation of Y529 within the MID domain 5′-phosphate binding pocket impairs P-body localization and small RNA association (Rüdel et al. 2011). In C. elegans, ALG-1 is heavily phosphorylated, including a hyperphosphorylated serine/threonine cluster (S988:S998) in the PIWI domain that promotes efficient AGO-miRISC assembly and target silencing (Quévillon Huberdeau et al. 2022; Shah et al. 2023). Additionally, phosphorylation of ALG-1 at S642 in the MID domain by KIN-1 reduces miRNA binding (Quévillon Huberdeau et al. 2022). Whether phosphorylation more broadly fine-tunes AGO binding specificity remains an open question.
Overall, these studies illustrate that PTMs provide a versatile mechanism for regulating Argonaute function, fine-tuning Argonaute activity and specificity across diverse biological contexts.
Chaperones and other interacting proteins
Argonaute proteins associate with a variety of cofactors that influence small RNA production, loading, and RISC assembly. These factors contribute to small RNA loading specificity through multiple mechanisms: by modulating small RNA biogenesis, by directly assisting in loading, or by affecting Argonaute localization or binding affinity (Table 3).
Summary of Argonaute protein interactors
Molecular chaperones directly facilitate Argonaute loading. In vitro studies show that HSP70, HSP90, and the co-chaperone HSP40 drive ATP-dependent conformational changes in human AGO2 and Drosophila Ago1/Ago2, enabling efficient siRNA binding (Iwasaki et al. 2010; Miyoshi et al. 2010; Yoda et al. 2010; Naruse et al. 2018). Single-molecule FRET experiments with purified Drosophila Ago2 further revealed that HSP70 and HSP40 initiate the conformational change, while HSP90 stabilizes an open state that promotes small RNA loading (Tsuboyama et al. 2018). Similar in vitro studies across plants, flies, and mammals confirm that Argonautes and chaperones alone are sufficient for RISC assembly (Iki et al. 2010; Iwasaki et al. 2015).
In C. elegans, HSP40 co-chaperones also contribute to miRNA loading. DNJ-12, a member of the HSP40 family, interacts specifically with the N-terminal domain of ALG-1, but not with ALG-2 or other siRNA-binding Argonautes. This interaction facilitates ALG-1 association with HSP70 and enables efficient loading of a subset of miRNAs. Loss of DNJ-12 selectively reduces binding of these miRNAs while increasing binding of others, leading to incomplete miRISC assembly and associated miRNA-related phenotypes (Frédérick et al. 2024). These findings identify DNJ-12 as a critical factor directing particular miRNAs to ALG-1. The C. elegans genome encodes over 90 chaperone and co-chaperone proteins, including HSP40 family members DNJ-1 through DNJ-30, yet the roles of most remain uncharacterized in the context of small RNA pathways (Brehme et al. 2014; Frumkin et al. 2014). This diversity hints that additional chaperones may act in concert with other Argonautes to fine-tune loading specificity.
A parallel mechanism is observed in plants. In Arabidopsis, AGO10 employs the HSP70/HSP90 chaperone system to selectively load miR166/165 duplexes. This selectivity is influenced by a central PIWI loop in AGO10 (residues 714–720), underscoring how individual Argonaute domains can shape chaperone-mediated loading specificity (Xiao and MacRae 2022).
Beyond chaperones, other protein interactors can also directly influence Argonaute loading. A clear example comes from Drosophila, where distinct small RNA processing and loading factors recognize different duplex structures, directing them to the appropriate Argonaute. Pre-miRNAs are processed into miRNAs by Dicer-1, whereas long dsRNAs are cleaved into siRNAs by Dicer-2, which then forms a stable heterodimer with the cofactor R2D2. This Dicer-2/R2D2 complex specifically loads siRNAs onto Ago2 and prevents misloading onto Ago1 (Liu et al. 2003; Förstemann et al. 2007; Tomari et al. 2007; Okamura et al. 2011). Strand selection and loading are dictated by the thermodynamic asymmetry of siRNA duplexes: R2D2 binds the end with higher base-pairing stability, while Dicer-2 recognizes the other end, orienting the guide siRNA strand for Ago2 incorporation (Tomari et al. 2004). A similar mechanism occurs in C. elegans, where the dsRNA-binding protein RDE-4 mediates siRNA processing by Dicer/DCR-1, and selectively promotes loading into RDE-1, but does not participate in miRNA loading into ALG-1 (Knittel et al. 2025).
Another layer of regulation comes from RNA helicases of the SF2 superfamily, whose ATP-dependent remodeling activities help shape small RNA loading and Argonaute specificity. In C. elegans, the germ granule proteins GLH-1 and GLH-4, DEAD-box helicases related to Drosophila VASA, physically interact with WAGO-1 and its target mRNAs (Marnik et al. 2019; Chen et al. 2020; Dai et al. 2022). Loss of GLH-1 causes misloading of some CSR-class 22G-RNAs into WAGO-1 and HRDE-1, likely due to defects in retaining primary target mRNAs within germ granules or in promoting WAGO-class 22G-RNA biogenesis and loading (Dai et al. 2022). Similarly, the FG repeats of GLH-family proteins, including GLH-1, GLH-2, and GLH-4, were recently shown to regulate the subcellular localization and loading specificity of WAGO-4. In FG-repeat mutants, WAGO-4 mislocalizes from germ granules to the cytoplasm and binds 22G-RNAs that more closely resemble the CSR-1 profile (Jelenic et al. 2025). These studies demonstrate that RNA helicases may act as critical modulators of Argonaute loading fidelity, ensuring that the correct small RNAs associate with their appropriate Argonaute partners.
In the C. elegans siRNA pathway, the nuclear Argonaute proteins HRDE-1 and NRDE-3 depend on interactions with their respective paralogous cofactors HRDE-2 and ENRI-2 (Lewis et al. 2020; Chen and Phillips 2024, 2025). These interactions occur specifically with unloaded Argonautes in granules marked by the Tudor domain–containing protein SIMR-1: HRDE-1 and HRDE-2 in germ cells, and NRDE-3 and ENRI-2 in early embryos. Disruption of the HRDE-2–HRDE-1 interaction causes unloaded HRDE-1 to relocalize from germ granules to the cytoplasm, resulting in misloading. HRDE-1 switches from binding WAGO-class 22G-RNAs to CSR-class 22G-RNAs, leading to ectopic H3K9me3 deposition at CSR targets (Chen and Phillips 2024). In contrast, loss of ENRI-2 has only a subtle effect on NRDE-3 loading, suggesting the involvement of additional factors in ensuring its specificity (Chen and Phillips 2025). These specific examples illustrate the diverse ways in which cofactors influence Argonaute loading.
Overall, Argonaute-interacting proteins can shape binding specificity through multiple mechanisms. They can act directly, by guiding the loading of particular small RNAs, or indirectly, by influencing upstream small RNA biogenesis, Argonaute subcellular localization, or the availability of binding partners. In this way, interactors provide an additional layer of regulation, ensuring that Argonautes are loaded with the correct small RNA populations with high specificity and fidelity.
SPATIAL AND TEMPORAL COMPARTMENTALIZATION
In addition to small RNA biochemical features, Argonaute structure, and interacting partners, the spatial and temporal expression of both Argonaute proteins and small RNAs is a critical determinant of binding specificity (Fig. 3C). Unraveling the complexity of this regulation is particularly tractable in C. elegans, where the expanded Argonaute repertoire and specialized granules provide a powerful system to dissect mechanisms of small RNA sorting and function across both time and space.
Temporal regulation
Distinct Argonaute proteins and their preferred small RNAs are expressed at specific developmental stages in C. elegans. Temporally regulated miRNAs in the heterochronic pathway provide classic examples of how precise temporal control of small RNAs determines cell lineage fate. For example, lin-4, let-7, miR-48, miR-84, and mir-241 are expressed in sequential waves during larval development, regulating distinct transcription factors to drive cell fate transitions. Consistent with this, miRNA-binding Argonaute proteins, such as ALG-1 and ALG-2, are highly expressed during larval stages (Pasquinelli et al. 2000; Großhans et al. 2005; Resnick et al. 2010; Sokol 2012).
Many siRNAs exhibit similar temporal regulation alongside their Argonaute partners. CSR-1A, for example, is expressed in the germline during the L4 stage and binds spermatogenic 22G-RNAs, whereas CSR-1B is expressed in adults and binds oogenic 22G-RNAs (Nguyen and Phillips 2021). Expression of either isoform under a ubiquitous promoter in a csr-1 null mutant rescues fertility (Charlesworth et al. 2021), suggesting that developmental timing is a key determinant of small RNA binding specificity. Similarly, the nuclear Argonaute NRDE-3 switches small RNA partners during embryogenesis, from CSR-class 22G-RNAs in early stages to ERGO-class 22G-RNAs later (Chen and Phillips 2025), likely reflecting changing small RNA availability across developmental time.
Spatial regulation
Argonautes must colocalize with their small RNA partners in tissue and subcellular compartments. In C. elegans, eight of 19 Argonautes are germline-specific, three are soma-restricted (SAGO-1, SAGO-2, and ALG-1), and the remainder are expressed in both tissues (Seroussi et al. 2023). Within the gonad, expression is regionally restricted: ERGO-1 and NRDE-3 are expressed only in the oocyte region, whereas CSR-1A, ALG-3, ALG-4, and WAGO-10 are confined to the spermatogenic region (Seroussi et al. 2023). Small RNA partners are presumed to follow similar spatial restrictions, although this hypothesis remains to be formally tested.
Small RNAs also differ in where they are produced. Whereas many small RNAs are processed in the cytoplasm or in germ granules, piRNA biogenesis occurs on the mitochondrial surface in multiple organisms (Weick and Miska 2014; Ding et al. 2019; Gao et al. 2025). In C. elegans, disruption of the mitochondrial tethering of the PUCH processing complex severely reduces mature piRNAs and likely impairs their subsequent loading into PRG-1 (Podvalnaya et al. 2023). Although PRG-1 loading itself has not been shown to occur at mitochondria, these findings raise the possibility that the spatial organization of piRNA biogenesis may influence Argonaute loading by modulating where and when mature piRNAs become available. Whether similar spatial constraints apply to other small RNA classes remains unknown.
Spatial compartmentalization within germ granules further refines specificity. CSR-class 22G-RNAs are produced in the cytosol and in E granules, while WAGO-class 22G-RNAs are synthesized in Mutator foci (Phillips et al. 2012; Singh et al. 2021; Chen et al. 2024b). This compartmentalized biogenesis contributes to HRDE-1 and WAGO-4 loading specificity: HRDE-1 is recruited to SIMR foci by HRDE-2 to ensure correct loading with WAGO-class 22G-RNAs. In hrde-2 mutants, HRDE-1 mislocalizes to the cytosol and inappropriately loads CSR-class 22G-RNAs (Chen and Phillips 2024). WAGO-4 shows a similar sensitivity to granule positioning; mutations that disrupt its anchoring within germ granules cause cytoplasmic mislocalization and a shift toward CSR-like 22G-RNA binding (Jelenic et al. 2025). By extension, mislocalization to the wrong granule may lead to improper small RNA loading, although this remains to be directly tested.
While analogous granule subdomains have not yet been clearly defined in other organisms, germ granules in Drosophila and zebrafish exhibit dynamic spatial organization. Super-resolution microscopy and single-molecule imaging reveal that ribosomes and translating mRNAs localize to the granule periphery, with untranslated mRNAs stored centrally (Westerich et al. 2023; Ramat et al. 2024). Moreover, in Drosophila, translating mRNAs further adopt a defined orientation in granules: The coding sequence and associated polysomes are at the granule surface, while the 3′ UTR is anchored internally (Chen et al. 2024a); disrupting this organization reduces translation efficiency (Ramat et al. 2024). Similar toroidal organization occurs in P and Z compartments of C. elegans germ granules (Uebel et al. 2023), though the relationship to mRNA translation remains unclear. Together, these findings suggest that mRNA orientation, compartmentalized small RNA biogenesis, and selective Argonaute loading may rely on conserved organizational principles across species.
CONCLUSION
Achieving specific Argonaute–small RNA pairing requires multilayered regulation: Argonautes and small RNAs must be expressed at the correct developmental stage, in the appropriate tissue, and within the correct subcellular compartment. Structural domains of Argonautes, post-translational modifications, and interaction with binding partners further refine small RNA incorporation. Collectively, these mechanisms ensure that Argonautes engage the correct small RNA populations, directing precise downstream effects on gene expression. While many mechanistic insights have been elucidated in C. elegans, emerging parallels across diverse organisms suggest that the principles governing Argonaute specificity and granule organization are broadly conserved in shaping development and maintaining genome integrity.
OPEN QUESTIONS
-
Spatiotemporal regulation of RNAi: How does small RNA expression change across development? Are different classes of small RNAs in C. elegans regulated in time and space in coordination with their mRNA targets? Can emerging high-resolution microscopy methods directly visualize their spatial and subcellular distributions? Are individual steps of RNAi, including small RNA biogenesis, Argonaute loading, and Argonaute-mediated silencing, spatially organized within germ granules?
-
Argonaute expression patterns: How is the precise spatial and temporal expression of individual Argonaute proteins established, maintained, and coordinated with small RNA populations?
-
Determinants of WAGO specificity: Why do WAGO Argonautes harbor a divergent HKQK motif within the conserved small RNA binding pocket, and does this residue underlie their specificity for triphosphorylated 22G-RNAs? How do highly similar WAGO Argonautes discriminate among distinct small RNA classes?
-
Role of N-terminal regions: What is the molecular contribution of intrinsically disordered N-terminal regions to Argonaute binding specificity? How conserved are the functions of the N-terminal regions among different Argonautes in C. elegans and across species?
-
Post-translational modifications: How do Argonaute PTMs influence small RNA binding specificity? Do they act by altering Argonaute structure, expression, localization, or interactions with binding partners?
Footnotes
-
Abbreviations: (sRNA) small RNA, (ncRNA) noncoding RNA, (miRNA) microRNA, (pri-miRNA) primary miRNA, (pre-miRNA) precursor miRNA, (piRNA) PIWI-interacting RNA, (siRNA) small interfering RNA, (dsRNA) double-stranded RNA, (lncRNA) long noncoding RNA, (nt) nucleotide, (Pol II) RNA polymerase II, (AGO) Argonaute, (WAGO) worm-specific Argonaute, (RNAi) RNA interference, (RISC) RNA-induced silencing complex, (RdRP) RNA-dependent RNA polymerase, (3' UTR) 3' untranslated region, (CDS) protein-coding region, (DNAi) DNA interference, (TGS) transcriptional gene silencing, (PTGS) post-transcriptional gene silencing, (NLS) nuclear localization signal, (NES) nuclear export signal, (NTE) N terminal extension, (PTM) post-translational modification, (PRMT) protein Arginine methyltransferase, (RG motif) arginine-glycine-rich motif, (IDR) intrinsically disordered region
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080816.125.
-
Freely available online through the RNA Open Access option.
This article, published in RNA, is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵








