
RNase L produces tRNA-derived RNAs that contribute to translation inhibition
- 1Laboratory of Oncology, Pharmacy Practice and Sciences, Tohoku University Graduate School of Pharmaceutical Sciences, Sendai 980-8578, Japan
- 2Division of Rheumatology, Inflammation and Immunity, Brigham and Women's Hospital, Boston, Massachusetts 02115, USA
- 3Department of Medicine, Harvard Medical School, Boston, Massachusetts 02115, USA
- 4Harvard Initiative for RNA Medicine, Boston, Massachusetts 02115, USA
- Corresponding authors: yasutoshi.akiyama.b4{at}tohoku.ac.jp; pivanov{at}bwh.harvard.edu
-
↵5 These authors contributed equally to this work.
-
Handling editor: Eric Phizicky
Abstract
Ribonuclease L (RNase L) is an RNase which is activated by viral double-stranded RNAs (dsRNAs). RNase L cleaves not only viral RNAs but also host RNAs, including mRNAs and tRNAs, which contributes to innate immune defense against viruses. While it has been reported that RNase L-mediated bulk mRNA cleavage induces rapid translation repression independently of the integrated stress response, the significance of RNase L-mediated tRNA cleavage remains largely unknown. Here we show that RNase L cleaves various tRNA species in the anticodon loops, generating transfer RNA-derived RNAs (tDRs) similar to tRNA-derived stress-induced RNAs (tiRNAs) that are generated by a stress-responsive RNase angiogenin (ANG). Three tRNA species (tRNALeu, tRNASeC, and tRNASer) were cleaved within the variable loops as well as in the anticodon loops by RNase L, generating noncanonical tDRs. As RNase L-induced 5′-tDRAla/Cys were similar in length to 5′-tiRNAAla/Cys that possess a translation inhibitory effect, we examined whether RNase L-induced 5′-tDRAla also inhibited translation. In vitro translation analysis showed that RNase L-induced 5′-tDRAla significantly inhibits mRNA translation like 5′-tiRNAAla, suggesting that the production of 5′-tDRAla may be involved in the mechanism of RNase L-mediated stress response during viral infection. Our data shed new light on the potential roles of tDRs in innate immunity against viral infection.
Keywords
INTRODUCTION
Transfer RNA-derived RNAs (tDRs) are small RNAs produced by endonucleolytic cleavage of transfer RNAs (tRNAs) (Akiyama and Ivanov 2023; Holmes et al. 2023). There is accumulating evidence that tDRs, especially those produced in response to stress, can play essential roles in cellular stress response (Akiyama and Ivanov 2023). Under stress conditions, tDRs are produced predominantly by a stress-responsive ribonuclease (RNase), angiogenin (ANG). ANG is a vertebrate-specific RNase, which belongs to RNase A superfamily (Lyons et al. 2017a). Under physiological conditions, ANG is inactivated by forming a complex with RNH1, an endogenous inhibitor of RNase A superfamily enzymes. When cells encounter a variety of stress stimuli such as UV irradiation, heat shock, oxidative stress, and sodium arsenite treatment, ANG becomes activated by dissociating from RNH1. A recent study has revealed that the active ANG binds to the A site of the 80S ribosome, which further activates the RNase catalytic center. Then the fully active ANG cleaves incoming tRNAs at the A site (Loveland et al. 2024). ANG cleaves tRNAs in the anticodon loops, resulting in the production of nicked tRNAs (Costa et al. 2023). These nicked tRNAs are either repaired to their original form by the RTCB ligase complex (Akiyama et al. 2022) or separated into two kinds (5′-side and 3′-side) of tDRs by ATP-dependent RNA helicases (Drino et al. 2023). These stress-induced tDRs are called tRNA-derived stress-induced RNAs (tiRNAs) (Yamasaki et al. 2009).
Among a variety of tiRNAs, 5′-tiRNAs derived from tRNAAla and tRNACys (5′-tiRNAAla and 5′-tiRNACys) specifically possess a translation inhibitory effect through a stretch of five guanosines at their 5′ ends, termed the terminal oligoguanine (TOG) motif (Ivanov et al. 2011). Mechanistically, these TOG-containing 5′-tiRNAs form tetrameric G-quadruplex (G4) structures (Ivanov et al. 2014; Lyons et al. 2017b). G4-tiRNAs displace the eukaryotic initiation factor eukaryotic initiation factor 4F (eIF4F) complex from the cap structure of mRNAs through direct binding to eIF4G (Lyons et al. 2020), one of the subunits of eIF4F complex, which represses the scanning step of translation initiation. This global translation inhibition contributes to cell protection by conserving energy for recovery from stress-induced damage (Advani and Ivanov 2019). ANG-induced tDRs are now recognized as an important determinant of cell fate such as stem cell differentiation (Goncalves et al. 2016; Flores et al. 2017), cancer progression (Honda et al. 2015; Garcia-Vilchez et al. 2023), neurodegenerative disorders (Ivanov et al. 2014), and host defense through myeloid differentiation (Kfoury et al. 2021). At least part of them are likely mediated by translational regulatory effects of tDRs.
When cells are infected with viruses, the innate immune defense system is stimulated in response to double-stranded RNAs (dsRNAs), a hallmark of viral replication. This antiviral response is mediated by a variety of dsRNA-sensing molecules such as RIG-I-like receptors (RLRs) for type I interferon production, protein kinase R (PKR) for triggering the integrated stress response, and oligoadenylate synthetases (OASs) for activation of ribonuclease L (RNase L) (Hur 2019). Translational repression is one of the key stress responses in viral infection as well as other stress conditions. During viral infection, PKR, one of the four eIF2α kinases that trigger the integrated stress response, is activated through the interaction with viral dsRNAs, which causes global translational inhibition via eIF2α phosphorylation (Pakos-Zebrucka et al. 2016; Costa-Mattioli and Walter 2020). In addition to PKR, recent studies have shown that RNase L also plays a pivotal role in translational repression, especially in early phase of viral infection before the phosphorylation of eIF2α (Donovan et al. 2017; Burke et al. 2019; Rath et al. 2019).
RNase L is a cytoplasmic ribonuclease which contributes to innate immune response against viruses (Karasik and Guydosh 2024). Under normal conditions, RNase L remains inactive as a monomer. In response to viral dsRNAs, OASs synthesize 2′-5′-linked oligoadenylates (2-5A), which in turn binds to inactive RNase L, resulting in dimerization and activation of RNase L (Schwartz and Conn 2019). Once activated, RNase L rapidly degrades bulk mRNAs except antiviral genes, resulting in global translational repression independently of the integrated stress response (Burke et al. 2019; Rath et al. 2019). As RNase L has a promiscuous recognition sequence (UN^N) (Han et al. 2014), small noncoding RNAs, such as tRNAs and Y-RNAs, are also known to be cleaved into fragments by RNase L (Donovan et al. 2017; Rath et al. 2019). It has been reported that three kinds of tRNAs, tRNAHis, tRNAPro, and tRNAGln, are preferentially cleaved by RNase L into tDRs (Donovan et al. 2017). However, it has yet to be determined whether other tRNA species can be cleaved into tDRs by RNase L. In addition, although it is unlikely that RNase L-mediated tRNA cleavage causes significant depletion of functional tRNAs (Donovan et al. 2017), it is still unclear whether RNase L-induced tDRs contribute to translational regulation during viral infection.
As ANG-induced 5′-tiRNAs function as stress-responsive molecules through their inhibitory effects on protein synthesis, we hypothesized that RNase L-induced tDRs also play a role in translational regulation under viral infection. Here we show that RNase L cleaves various kinds of tRNAs into tiRNA-like tDRs. We also show that RNase L-induced 5′-tDRAla, like 5′-tiRNAAla, is capable of repressing translation, which may be involved in the stress response during viral infection.
RESULTS
RNase L-mediated tRNA cleavage induced by poly(I:C) transfection
Although a previous study reported that RNase L potently cleaves three tRNA species (tRNAHis, tRNAPro, and tRNAGln) into tDRs (Donovan et al. 2017), it remains unclear whether RNase L cleaves other tRNAs. First, we sought to identify tRNAs that are cleaved by RNase L. Polyinosinic-polycytidylic acid [poly(I:C)], a synthetic analog of dsRNA, has been widely used as a viral mimetic to activate RNase L. Poly(I:C) transfection induced the production of RNA fragments of ∼30–50 nt in length, suggesting that they were originated from tRNAs (Fig. 1A). Northern blotting showed that poly(I:C) transfection induced 5′-tDRs derived from all three tRNA species that were reported to be targets of RNase L (Donovan et al. 2017) in A549 cells, while 5′-tDRGln-CTG was barely detected in U2OS cells (Fig. 1B). These data suggest that under our conditions, poly(I:C) transfection induced RNase L activation, resulting in tRNA cleavage as previously reported (Donovan et al. 2017). It is worth noting that the amount of poly(I:C)-induced tDRs was higher in A549 cells than in U2OS cells (Fig. 1A,B). RNase L as well as OAS3, one of the RNase L activators (Schwartz and Conn 2019), are more highly expressed in A549 cells than in U2OS cells (Supplemental Fig. S1A), explaining more active tDR production in A549 cells. These data also support previous data from the Korennykh laboratory that RNase L is responsible for the tRNA cleavage (Donovan et al. 2017).
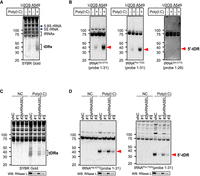
RNase L-mediated tRNA cleavage induced by poly(I:C) transfection. (A, B) Poly(I:C) transfection induces tRNA cleavage in U2OS/A549 cells. (A) SYBR Gold staining and (B) northern blotting for tRNAHis-GTG, tRNAPro-TGG, and tRNAGln-CTG. (C, D) RNASEL knockdown decreases the amount of poly(I:C)-induced tDRs in A549 cells. (C) SYBR Gold staining and (D) northern blotting for tRNAHis-GTG and tRNAPro-TGG. RNase L expression levels evaluated by western blotting are also shown. Red arrowheads indicate 5′-tDRs generated by RNase L-mediated cleavage.
Next, we examined the effect of RNASEL knockdown on poly(I:C)-induced tRNA cleavage to confirm that RNase L was solely responsible for the poly(I:C)-induced tRNA cleavage. We generated three shRNA constructs for RNASEL knockdown (Fig. 1C,D; Supplemental Fig. S1B). The construct #1, which showed the highest knockdown efficiency (∼80% reduction), accordingly most strongly inhibited poly(I:C)-induced tDR production in A549 cells (Fig. 1C,D). On the other hand, the effect of construct #2, which showed relatively low knockdown efficiency (Supplemental Fig. S1B), was modest compared to the construct #1 (Fig. 1C,D), which suggests that the amount of poly(I:C)-induced tDRs is dependent on intracellular RNase L expression levels. We also examined the amount of poly(I:C)-induced tDRs in RNASEL knockout (ΔRNASEL) cells (Supplemental Fig. S1C–E). RNase L depletion completely abolished poly(I:C)-induced tDRs production in A549 cells (Supplemental Fig. S1D,E), further supporting that poly(I:C)-induced tDRs are solely due to RNase L-mediated tRNA cleavage.
Since RNase L is activated by 2′-5′-linked oligoadenylate (2-5A) molecules synthesized by 2′-5′-OASs (Schwartz and Conn 2019), we investigated whether the amount of RNase L-induced tDRs also depends on the expression level of OAS3, the major OAS isoform responsible for RNase L activation (Li et al. 2016). We confirmed that OAS3 knockdown did not affect the expression level of RNase L and vice versa (Supplemental Fig. S1F). Under the condition, OAS3 knockdown decreased the amount of poly(I:C)-induced tDRs like RNASEL knockdown (Supplemental Fig. S1G,H), suggesting that OAS3 expression level is also a determinant of poly(I:C)-induced tDR production.
RNase L cleaves various tRNAs into tDRs
While various tRNA sequencing approaches exist, most of them still have some technical limitations (Cabrelle et al. 2024; Padhiar et al. 2024). Next, we sought to identify tRNA species that are cleaved by RNase L in A549 cells using traditional northern blotting approach. Among 22 tRNA species we evaluated, 20 tRNAs were cleaved by RNase L, resulting in the detection of 5′-tDRs, while 5′-tDRs originating from tRNAArg and tRNAThr were not detected by northern blotting (Fig. 2A,B). As indicated by red arrowheads in Figure 2B, the length of RNase L-induced 5′-tDRs was ∼30–40 nt, suggesting that RNase L cleaves tRNAs in the anticodon loops.
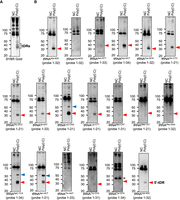
RNase L cleaves various tRNA species into tDRs in A549 cells. RNase L activated by poly(I:C) transfection induces tDRs originating from various tRNA species. (A) SYBR Gold staining and (B) northern blotting. Red and blue arrowheads indicate 5′-tDRs likely generated by cleavage in the anticodon loops and variable loops, respectively. Asterisk indicates 5′-tDR derived from pre-tRNATyr-GTA (Akiyama et al. 2022).
ANG cleaves the anticodon loops of tRNAs in response to stress stimuli, generating tDRs called tiRNAs (Yamasaki et al. 2009). As RNase L was suggested to cleave tRNAs in the anticodon loops similar to ANG, we compared ANG-induced with poly(I:C)-induced cleavage patterns using U2OS cells (Supplemental Fig. S2). SYBR Gold staining showed that both poly(I:C) transfection and ANG treatment induced RNA fragments of ∼30–50 nt in length (Supplemental Fig. S2A). Northern blotting revealed that poly(I:C)-induced tDRs showed a similar, but different pattern from ANG-induced tiRNAs in length (Supplemental Fig. S2B). For example, the lengths of 5′-tDRAla-AGC, 5′-tDRCys-GCA, and 5′-tDRGln-CTG were different from those of 5′-tiRNAAla-AGC, 5′-tiRNACys-GCA, and 5′-tiRNAGln-CTG, respectively, suggesting that RNase L cleaves such tRNAs at different sites from that by ANG. Importantly, these data show that ANG remained inactive under the condition where poly(I:C) was transfected, suggesting stress-specific specialization of various RNases in cells. In addition, ANG efficiently cleaved tRNAArg-ACG unlike RNase L (Supplemental Fig. S2B). Taken together, we concluded that RNase L cleaves various tRNAs in the anticodon loops, generating similar but different profiles of tDRs than that of ANG-mediated tiRNAs.
RNase L also cleaves variable loops of tRNAs
We noticed that, in addition to 5′-tDRs that were supposed to be generated by cleavage in the anticodon loops, longer 5′-tDRs were also detected in three tRNA species, namely tRNALeu-CAG, tRNASeC-TCA, and tRNASer-GCT, as indicated by blue arrowheads in Figure 2B. The longer 5′-tDRs were ∼50 nt in length. As these three tRNAs are known to possess long variable loops (Chan and Lowe 2016), we hypothesized that RNase L cleaved the variable loops of these tRNAs, generating 5′-tDRs of ∼50 nt in length. If these tRNAs were cleaved in the variable loops, 3′-tDRs of ∼35 nt would be predicted to be produced, in addition to those of ∼50 nt specific to cleavage within the anticodon loops. All three tRNA species contain UN^N motifs within the variable loops (Fig. 3), the consensus sequence of RNase L-mediated cleavage (Han et al. 2014). By northern blotting using 3′-probes that hybridize the sequences downstream from the variable loops (Fig. 3), 3′-tDRs of ∼35 nt were detected in all the three tRNAs, supporting the cleavage in the variable loops by RNase L. Thus, RNase L targets the variable loops as well as the anticodon loops in tRNAs that possess long variable loops.
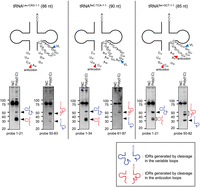
RNase L can target variable loops of tRNAs. To identify how the longer 5′-tDRs were generated from tRNALeu-CAG, tRNASec-TCA, and tRNASer-GCT, northern blotting was performed using both 5′ and 3′ probes for these tRNAs. The sequences around anticodon loops and variable loops in these tRNAs are shown. Predicted cleavage sites in anticodon loops and variable loops are indicated by red and blue arrowheads, respectively. (VL) Variable loop.
RNase L-induced 5′-tDRAla inhibits translation
tDRs are now widely recognized to play diverse roles in cellular functions especially under stress conditions through translational regulation (Akiyama and Ivanov 2023). One of the most well-studied subclasses of tDRs are ANG-induced tiRNAs. Several tiRNAs, such as 5′-tiRNAAla, 5′-tiRNACys, 5′-tiRNAGly, and 3′-tiRNAPro, have been shown to exert translational inhibitory effects (Ivanov et al. 2011). As we revealed that RNase L cleaves various tRNA species into tiRNA-like tDRs including 5′-tDRAla and 5′-tDRCys, we sought to examine whether such RNase L-induced tDRs also play roles in stress response especially by inhibiting translation.
First, we compared the length of RNase L-induced 5′-tDRAla with ANG-induced 5′-tiRNAAla, one of the most intensively studied tDRs that show a translation inhibitory effect (Ivanov et al. 2011, 2014; Lyons et al. 2017b). Northern blotting showed that ANG produces two kinds of 5′-tiRNAAla, whereas RNase L produces only one kind of 5′-tDRAla (Fig. 4A). The mobility of poly(I:C)-induced 5′-tDRAla was between those of two ANG-induced 5′-tiRNAsAla, suggesting that RNase L cleaves tRNAAla-AGC between the two cleavage sites specific to ANG. As ANG cleaves the 3′ side of pyrimidine nucleotides (Lyons et al. 2017a), while RNase L cleaves RNAs at UN^N motif (Han et al. 2014), we predicted cleavage sites by ANG/RNase L (Fig. 4B). We predicted that RNase L produces 5′-tDRAla of 33 nt, while ANG produces two kinds of 5′-tiRNAAla (32 and 35 nt). Synthetic 5′-tDRsAla of 32 and 33 nt showed the same mobility as that of the shorter 5′-tiRNAAla and RNase L-induced 5′-tDRAla, respectively, which is consistent with the prediction (Fig. 4C,D). These data suggest that RNase L produces 5′-tDRAla of 33 nt with 1 nt difference in length from 5′-tiRNAAla. According to the tDRnamer (Holmes et al. 2023), this RNase L-induced 33 nt tDRAla was named tDR-1:34-Ala-AGC-2-M4, while ANG-induced 5′-tiRNAsAla of 32 and 35 nt were named tDR-1:33-Ala-AGC-2-M5 and tDR-1:36-Ala-AGC-2-M4, respectively. We also compared the length between RNase L-induced 5′-tDRCys and 5′-tiRNACys (Supplemental Fig. S3). As northern blotting showed that 5′-tDRCys is slightly shorter than 5′-tiRNACys (Supplemental Fig. S3A), we predicted the cleavage sites by RNase L/ANG (Supplemental Fig. S3B). Based on the prediction, RNase L-induced 5′-tDRCys and 5′-tiRNACys were named tDR-1:34-Cys-GCA-2-M2 and tDR-1:35-Cys-GCA-2-M2, respectively (Holmes et al. 2023). There is only 1 nt difference in the estimated length between 5′-tDRCys (33 nt) and 5′-tiRNACys (34 nt), suggesting that they may have similar biological activities at least in terms of mRNA translation inhibition.
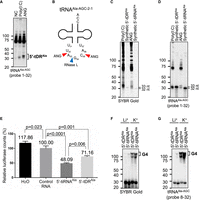
RNase L-induced 5′-tDRAla shows a translation inhibitory effect. (A–D) Comparison between RNase L-induced 5′-tDRAla and ANG-induced 5′-tiRNAAla. (A) Northern blotting for tRNAAla-AGC. (B) Predicted cleavage sites of tRNAAla-AGC-2-1 by ANG/RNase L. Red and blue arrowheads indicate the predicted sites of ANG-mediated and RNase L-mediated cleavage, respectively. (C, D) Estimation of the length of RNase L-induced 5′-tDRAla and ANG-induced 5′-tiRNAAla. Mobility of the tDRs was compared with that of synthetic 5′-tDRsAla (32 and 33 nt). (C) SYBR Gold staining and (D) northern blotting for tRNAAla-AGC. (E) 5′-tDRAla (33 nt) as well as 5′-tiRNAAla (32 nt) inhibit translation of mRNA reporters in vitro. Means and standard deviation were obtained from three independent experiments. Mean values and P-values are shown. (F, G) 5′-tDRAla as well as 5′-tiRNAAla can form G-quadruplex structure. (F) SYBR Gold staining and (G) northern blotting for tRNAAla-AGC.
Next, we evaluated whether RNase L-induced 5′-tDRAla has a translational inhibitory effect by in vitro translation assay using rabbit reticulocyte lysates (RRLs), which we used previously (Ivanov et al. 2011; Lyons et al. 2017b; Akiyama et al. 2020). RNase L-induced 5′-tDRAla (33 nt), as well as ANG-induced 5′-tiRNAAla (32 nt), significantly reduced the translation of luciferase mRNA reporter in vitro, although the inhibitory effect of RNase L-induced 5′-tDRAla was slightly, yet statistically significantly, weaker than that of 5′-tiRNAAla (Fig. 4E). It has been reported that 5′-tiRNAAla exerts translation inhibition through the formation of G4 structure (Lyons et al. 2017b), a noncanonical four-stranded RNA structure formed through guanine-rich sequences (Kharel and Ivanov 2024). We examined whether RNase L-induced 5′-tDRAla is also capable of forming G4 structures like 5′-tiRNAAla. In the presence of K+ ions, which are known as a G4-permissive condition, both 5′-tiRNAAla and 5′-tDRAla assembled G4 structures, whereas G4 assembly was inhibited under a G4-nonpermissive condition (in the presence of Li+) (Fig. 4F,G). These results suggest that RNase L-induced 5′-tDRAla may also contribute to translational repression through the G4 formation as 5′-tiRNAAla does. Taken together, these results suggest that RNase L-induced 5′-tDRAla may contribute to stress response under viral infection through its translational inhibitory effect.
DISCUSSION
It is now widely accepted that tDRs are not simple degradation products, but at least some of them contribute to cellular homeostasis and stress response as biologically active molecules (Akiyama and Ivanov 2023). Accumulating evidence has shown that in particular, tiRNAs (Yamasaki et al. 2009) (or tRNA halves [Fu et al. 2009]) play essential roles in cellular physiology and pathology including cell protection against a variety of stress stimuli (Akiyama and Ivanov 2024). Here, we revealed that RNase L cleaves various tRNA species into tDRs as observed with ANG-induced tiRNAs (Fig. 2), which leads us to speculate that RNase L-induced tDRs may play diverse roles under viral infection.
In this study, we used poly(I:C) transfection as a model of RNase L activation under viral infection. Under our conditions, RNASEL knockdown specifically decreased the amount of poly(I:C)-induced tDRs depending on the knockdown efficiency (Fig. 1C,D). In addition, poly(I:C)-induced tDR production was completely repressed in RNASEL knockout (ΔRNASEL) cells (Supplemental Fig. S1C–E). These data suggest that poly(I:C)-induced tRNA cleavage was solely due to RNase L activities. In other words, other RNases including ANG were not activated by poly(I:C) transfection, implying that dsRNA-responsive RNase L pathway is completely independent of ANG activation. It should be noted that while poly(I:C) transfection activated only RNase L, some viruses can also activate ANG. For instance, ANG-mediated tiRNA production is triggered by some viral infections such as respiratory syncytial virus (RSV) (Wang et al. 2013; Deng et al. 2015) or hepatitis B and C virus (HBV and HCV) (Selitsky et al. 2015), which suggests that tDRs can be simultaneously produced by both ANG and RNase L under viral infection. Therefore, the profile of tDRs under viral infection is likely more complex than that of our simple experimental model. Further investigation is needed to clarify the expression profile and functions of tDRs in virus-infected cells.
We evaluated RNase L-mediated tRNA cleavage using two cell lines, U2OS cells and A549 cells. Although poly(I:C) transfection induced tRNA cleavage in both cell lines, tDRs were more actively produced in A549 cells than in U2OS cells (Fig. 1A,B). This directly corresponds to expression levels of RNase L and OAS3, which are higher in A549 cells compared to U2OS cells (Supplemental Fig. S1A). Thus, our data suggest that the expression levels of RNase L as well as its activators OASs are determinants of tDR production levels in response to dsRNA. Importantly, these data suggest that even if tDRs were not detected, it does not necessarily mean that RNase L did not cleave their parental tRNAs. For example, the number of detected tDRs was smaller in U2OS cells (Supplemental Fig. S2) than in A549 cells (Fig. 2), which was likely because these tDRs were not efficiently produced in U2OS cells due to lower RNase L activity. In addition, it is possible that cell line–specific tRNA modifications may have affected the efficiency of tDR production, as it was reported that tRNA modification in the anticodon loops can inhibit RNase L-mediated cleavage (Donovan et al. 2017). These differences likely contribute to cell type- or tissue-specific responsiveness to dsRNAs, which in turn could be biologically relevant to immunity.
Translation inhibition by ANG-induced 5′-tiRNAAla is one of the well-studied mechanisms of tDR-mediated stress response (Ivanov et al. 2011; Lyons et al. 2017b, 2020; Akiyama and Ivanov 2023). We showed that RNase L-induced 5′-tDRAla also possesses a translational inhibitory effect like ANG-induced 5′-tiRNAAla (Fig. 4E), which suggests that RNase L-induced tDR production may contribute to stress response through translational repression under viral infection. It should be noted that the translational inhibitory effect of 5′-tDRAla (33 nt) was slightly but significantly weaker than that of 5′-tiRNAAla (32 nt), although there is only 1 nt difference in length at their 3′ ends (Fig. 4E). While both possess the same 5′-TOG motif (Ivanov et al. 2011), which is required for G4 assembly (Lyons et al. 2017b), 5′-tiRNAAla (32 nt) seemed to form G4 structure more efficiently than 5′-tDRAla (33 nt) (Fig. 4F,G). There is a possibility that additional determinant outside of the TOG motif such as the adenine nucleotide at position 33 in 5′-tDRAla may affect the G4-forming efficiency, weakening the translational inhibitory effect. However, it is difficult to directly evaluate which tDR more strongly inhibits translation in cellulo. We found that ANG-mediated tRNAAla cleavage produced two kinds (likely 32 and 35 nt) of 5′-tiRNAAla (Fig. 4A,B), which suggests that 5′-tiRNAAla likely functions as a mixture of them. As we performed in vitro translation assay using only the shorter (32 nt) 5′-tiRNAAla, we could not evaluate the net effect of 5′-tiRNAAla compared with that of RNase L-induced 5′-tDRAla. Nevertheless, our data highlight length of tDRs as a contributing determinant of the tDR functions.
Our data suggest that RNase L can cleave some tRNAs not only in the anticodon loops but also in the variable loops (Fig. 3). To our best knowledge, there has been no report so far regarding tRNA cleavage within the variable loops. We identified three tRNA species (tRNALeu-CAG, tRNASeC-TCA, and tRNASer-GCT) whose variable loops are cleaved by RNase L. These tRNAs are known to possess long variable loops (Chan and Lowe 2016). Northern blotting analysis showed that the cleavage in the variable loops produces two types of “noncanonical” tDRs, 5′-tDRs consisting of the sequence from 5′ termini to the variable loops, and 3′-tDRs consisting of the sequence from the variable loops to 3′-CCA termini (Fig. 3). In addition, it may be possible that noncanonical intermediate tDRs consisting of the sequence from the anticodon to the variable loops (∼20 nt in length) are also generated by the cleavage at both anticodon and variable loops in a single tRNA molecule. Whereas northern blotting is not suitable for detection of such short RNAs due to low sensitivity and specificity, high-throughput RNA sequencing may be able to detect the intermediate tDRs. As these noncanonical tDRs might possess unknown biological functions, RNA sequencing–based comprehensive profiling is warranted to clarify the entire effect of RNase L-induced tDRs.
There is accumulating evidence that tRNA modifications affect the biological functions of tDRs. For instance, 1-methyladenosine (m1A) modification at position 58 in 3′-tDRs attenuates their gene-silencing activity by inhibiting annealing between the tDRs and target mRNAs (Su et al. 2022). In contrast, pseudouridine modification at position 8 is necessary for the translational inhibitory effect of 5′-tDRAla, 5′-tDRCys, and 5′-tDRVal (Guzzi et al. 2018). In addition, we have also reported that endogenous 5′-tiRNAGly more strongly inhibits translation than its synthetic counterpart (Akiyama et al. 2020). Therefore, synthetic RNAs, which do not possess any modified nucleotides, may not necessarily reflect the exact biological functions of endogenous tDRs, although can be used as tDR mimics in some applications including potential therapeutics. In this study, we used synthetic RNAs to examine the translational inhibitory effect of 5′-tDRsAla, which is one of the limitations of our study. Although we tried to purify endogenous 5′-tDRAla by hybridization-based pulldown (Akiyama et al. 2020; Takenaka et al. 2025b), the yield from confluent poly(I:C)-transfected A549 cells in twenty 10 cm dishes was <8 pmol, which was too low to perform in vitro translation analysis. Thus, it is still challenging to prepare enough tDRs for functional analyses because of low efficiency of RNase L-mediated tRNA cleavage. We previously developed a method termed “in lysate RNA digestion” which enables to efficiently cleave tRNAs by ANG in cell lysate, which significantly improve the yield of endogenous tiRNA pulldown (Akiyama et al. 2021). Development of the method to reconstitute active RNase L in vitro may bring a breakthrough in efficient preparation of endogenous RNase L-induced tDRs by combination with our in lysate RNA digestion method. Also, development of more sensitive experimental methods requiring smaller amount of tDRs are also strongly desirable for further establishing functions of RNase L-induced tDRs.
It is also worth noting that it remains challenging to investigate biological functions of RNase L-induced tDRs under viral infection because RNase L activation completely alters intracellular conditions, especially in terms of RNA metabolism and mRNA translation. Although ANG-mediated cleavage is highly specific to tRNAs (Akiyama et al. 2021; Loveland et al. 2024), RNase L cleaves various RNAs including host mRNAs (Prangley and Korennykh 2022). Once activated, RNase L rapidly cleaves up to 90% of host mRNAs, while some RNAs important for innate immune responses such as IFNB (coding interferon-β) mRNAs escape from RNase L-mediated cleavage (Burke et al. 2019; Rath et al. 2019). This widespread mRNA decay causes dynamic reprogramming of translation, such as permissive translation of mRNAs’ noncoding regions that are not efficiently translated under normal conditions (Karasik et al. 2021), nuclear retention of mRNAs, as well as nuclear translocation of RNA-binding proteins (Burke et al. 2021). Therefore, physiological functions of RNase L-induced tDRs should be evaluated under very specific cellular conditions. Despite these limitations, as translation initiation complexes and ribosomes remain functional under RNase L activation (Rath et al. 2019), it is theoretically possible that RNase L-induced 5′-tDRAla contributes to translational repression by displacing eIF4F complex from the cap structure of mRNAs.
In conclusion, we revealed that RNase L produces a variety of tDRs by cleaving most tRNAs in the anticodon loops. Some tRNAs are also cleaved in the variable loops, producing noncanonical tDRs. The profile of RNase L-induced tDRs is completely different from that of ANG-induced tiRNAs in terms of their length, abundance, and preferentially cleaved tRNA species (Supplemental Fig. S2). Among the tDRs, 5′-tDRAla showed a translational inhibitory effect likely through the same mechanism as that of ANG-induced 5′-tiRNAAla. Whereas further work is required to establish functions of each tDR, our findings hint at potential roles of RNase L-induced tDRs in innate immunity against viral infection.
MATERIALS AND METHODS
Cell culture and treatment
The human osteosarcoma-derived U2OS cells and lung adenocarcinoma-derived A549 cells were grown in 5% CO2 incubator at 37°C in Dulbecco's modified Eagle's medium (DMEM) (Nacalai Tesque) supplemented with 10% fetal bovine serum (Corning) and 1% of penicillin–streptomycin mixed solution (Nacalai Tesque).
For RNase L activation, poly(I:C) (HMW) (InvivoGen) was transfected into U2OS cells or A549 cells (5 × 105 cells) at a final concentration of 2 µg/mL using lipofectamine 3000 (ThermoFisher) according to the manufacturer's protocol. After 6 h incubation, cells were collected using Sepasol-RNA I Super G (Nacalai Tesque) for RNA purification.
In lysate ANG digestion method was performed as previously described (Akiyama et al. 2021). Recombinant human ANG (R&D Systems) was added into the lysate at indicated concentrations. Total RNA in the lysate was purified using Sepasol-RNA II Super (Nacalai Tesque).
Western blotting
Western blotting was performed as we previously reported (Akiyama et al. 2022). Antibodies used in this study are shown in Supplemental Table S1.
Knockdown of OAS3/RNASEL
Knockdown was performed as we previously reported (Akiyama et al. 2022). DNA oligonucleotides for shRNA targeting RNASEL or OAS3 were designed using siDirect (Naito et al. 2009), synthesized by Integrated DNA Technologies (IDT), and subcloned into pLKO.1 vector (Addgene) according to the manufacturer's instructions. The sequences of the oligonucleotides are shown in Supplemental Table S2. The lentiviral particles were produced by cotransfection of the pLKO.1 construct with pMD2.G and psPAX2 (Addgene) into Lenti-X 293T cells (Clontech) using lipofectamine 3000 (ThermoFisher). Cells were infected with lentiviral particles in the presence of 8 μg/mL polybrene (Sigma-Aldrich), then selected 1 day after viral transduction with 2 μg/mL of puromycin (Santa Cruz). Cells were collected 7 days after transduction, then subjected to experiments such as poly(I:C) transfection. Knockdown efficiencies of RNASEL or OAS3 were evaluated by western blotting, using anti-RNase L antibody or anti-OAS3 antibody, respectively. Western blotting against anti-β-actin antibody or Ponceau S staining (Nacalai Tesque) was used as loading control.
Generation of RNASEL knockout (ΔRNASEL) cells
DNA oligonucleotides for single-guide RNAs (sgRNAs) were designed using CRISPRdirect (Naito et al. 2015), synthesized by IDT, then subcloned into LentiCRISPRv2 vector (Addgene), according to the manufacturer's instructions. The sequences of sgRNAs are shown in Supplemental Table S3. The lentiviral particles were produced by cotransfection of the LentiCRISPRv2 construct with pMD2.G and psPAX2 (Addgene) into Lenti-X 293T cells (Clontech) using lipofectamine 3000 (ThermoFisher). The lentiviral particles (multiplicity of infection [MOI] = 0.3) were transduced into A549 cells in the presence of 8 μg/mL polybrene (Sigma-Aldrich), then selected with 2 μg/mL of puromycin (Santa Cruz) 1 day after viral transduction. Cells were cloned by limiting dilution and screened by western blotting using anti-RNase L antibody. LentiCRISPRv2 was a gift from Feng Zhang (Addgene, plasmid #52961) (Sanjana et al. 2014).
Northern blotting
Northern blotting was performed as we previously reported (Akiyama et al. 2022; Takenaka et al. 2025a). The sequences of the probes used in this study are shown in Supplemental Table S4. Each probe hybridizes to corresponding tRNAs at the position indicated as “probe position” in figures.
Generation of synthetic RNAs
All synthetic RNAs used in this study were synthesized by IDT. The sequences of the RNAs are shown in Supplemental Table S5.
G-quadruplex formation of 5′-tiRNAAla and 5′-tDRAla
The evaluation of G-quadruplex formation was performed as we previously described (Lyons et al. 2017b; Akiyama et al. 2020). Synthetic 5′-tDRAla (32 and 33 nt) was diluted to 10 μM in 100 mM KCl or LiCl solution, heated to 95°C for 10 min, and allowed to cool gradually to room temperature. Twenty picomoles each of 5′-tDRAla were run through a 15% polyacrylamide nondenaturing gel and subjected to northern blotting. After electrotransfer and UV crosslink, the membrane was hybridized with a biotinylated oligonucleotide probe that recognizes the 5′ side of tRNAAla (position 8–32), as shown in Supplemental Table S4. Then, the membrane was probed with HRP-conjugated streptavidin (BioLegend, 1:3000 dilution). Signals were visualized using Chemi-Lumi One L reagent (Nacalai Tesque) and detected with ChemiDoc imaging system (Bio-Rad), according to the manufacturer's instructions.
In vitro translation assay
In vitro translation was performed by the established protocols as we previously reported (Ivanov et al. 2011; Lyons et al. 2017b; Akiyama et al. 2020). Synthetic RNAs (150 pmol) were added to translation reactions, then incubated in 15 μL reaction volume at 30°C for 30 min. Luciferase activity was measured using Luciferase Assay System (Promega) on SpectraMax i3x (Molecular Devices). For statistical analyses, one-way ANOVA was performed for comparisons between groups. The Tukey–Kramer test was used for multiple comparisons.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
This work was supported by JST SPRING (JPMJSP2114 to Y.Ta.), the Japan Society for the Promotion of Science, Grants-in-Aid for Scientific Research (PS KAKENHI) (21K06865 and 23KK0290 to Y.A.; 23K06736 to Y.To.), and the National Institutes of Health R01 GM126150 and R01 GM146997 to P.I.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080419.125.
- Received February 9, 2025.
- Accepted March 26, 2025.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHORS


Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Yoshika Takenaka and Asuka Yamada are the co-first authors of this paper, “RNase L produces tRNA-derived RNAs that contribute to translation inhibition.” Yoshika is a third-year PhD student in the group of Professor Yoshihisa Tomioka at the Graduate School of Pharmaceutical Sciences, Tohoku University in Sendai, Japan. Yoshika's research focuses on cellular stress response during viral infection, especially that which is mediated by RNase L. Asuka graduated from the Faculty of Pharmaceutical Sciences, Tohoku University, last year after three years of research in the lab. Asuka is now pursuing a career as a pharmacist.
What are the major results described in your paper and how do they impact this branch of the field?
In this paper, we show that RNase L, a ribonuclease that is activated in response to viral dsRNAs, cleaves various tRNAs into tRNA-derived RNAs (tDRs). Among RNase L-induced tDRs, 5′-tDR derived from tRNAAla (5′-tDRAla) showed a translational inhibitory effect in vitro. We also reveal that the efficiency of RNase L-mediated tDR production is dependent on expression levels of both RNase L and OAS3, one of the activators of RNase L, likely determining cell type- and tissue-specific responsiveness to dsRNAs. Further, our results demonstrate that some tRNA species can be cleaved not only in the anticodon loops but also in the variable loops, which results in the production of noncanonical tDRs. Our findings shed new light on the potential roles of RNase L-induced tDRs in innate immune responses against viral infections.
What led you to study RNA or this aspect of RNA science?
YT: To be honest, I applied to Tomioka lab because I wanted to study antibody-based cancer therapy. But due to the personnel change of the staff in Tomioka lab, I unexpectedly started research about the regulation of tDRs by the RTCB ligase complex under Dr. Yasutoshi Akiyama who just came back to Japan from the Ivanov lab in Boston. Although I dove into the RNA world by accident, I became more and more interested in RNA biology day by day. Now I am very happy to specialize in RNA science!
AY: The first event that attracted me to RNA science was the lectures in biology at the university that I took before being assigned to the lab. Through the lectures, I was fascinated by diverse and complex functions of RNAs such as coordinated gene expression, maintenance of cellular homeostasis, and stress response. As I strongly desired to study RNA science, I applied to the Tomioka lab where Dr. Akiyama, the former postdoc in the Ivanov lab, is working on RNA biology.
During the course of these experiments, were there any surprising results or particular difficulties that altered your thinking and subsequent focus?
Since it was unclear how many tRNA species are cleaved by RNase L, the finding that RNase L targets most tRNA species was surprising for us. We were also intrigued that RNase L can target the variable loops of tRNAs as well as the anticodon loops, because there had been no report about the cleavage in the variable loops of tRNAs. These findings taught us that there are still so many unknowns in our research field and also taught us how exciting new discoveries by our own are. What was the most interesting is the finding that only 1 nt difference in length affects the translational inhibitory effect of 5′-tDRAla. Now we want to further probe the hypothesis that slight differences in length alter functions of tDRs.
What are some of the landmark moments that provoked your interest in science or your development as a scientist?
YT: I participated in the 29th Annual Meeting of the RNA Society in Edinburgh. That was one of the most important moments in my development. Fortunately, many researchers studying RNase L and antiviral immunity came to my poster. Discussion with researchers from all over the world was very exciting and valuable for me. This experience further inspired me to reveal potential roles of RNase L-induced tDRs in antiviral immunity.
AY: The experience of a poster presentation at the Annual Meeting of the RNA Society of Japan in 2023 was my landmark moment. Because this meeting was the first academic conference I attended, everything was new and exciting to me. During my presentation, I got the opportunity to discuss my data with many experts in the field of RNA biology. This experience was motivating to me to further investigate RNase L-induced tDR production and its effects on cellular stress response in viral infections.
What are your subsequent near- or long-term career plans?
YT: My next immediate plan is to defend my PhD thesis to obtain a doctorate in pharmaceutical science based on this study. I want to further explore this fascinating RNA world in the future.
What were the strongest aspects of your collaboration as co-first authors?
The strong aspect of our collaboration was that we trust each other as best friends as well as collaborators. Throughout our three-year collaboration, we always shared every single experimental result and discussed hypotheses and ideas to achieve the same goal. Because we had built the best relationship of trust, we were able to enjoy our research even when we were faced with challenges.








