
Cellular translational enhancer elements that recruit eukaryotic initiation factor 3
- Jiří Koubek1,
- Jaswinder Kaur1,
- Shivani Bhandarkar1,
- Cole J.T. Lewis1,
- Rachel O. Niederer1,3,
- Andrei Stanciu2,
- Colin Echeverría Aitken2 and
- Wendy V. Gilbert1
- 1Department of Molecular Biophysics and Biochemistry, Yale University, New Haven, Connecticut 06520, USA
- 2Biology Department and Biochemistry Program, Vassar College, Poughkeepsie, New York 12604, USA
- Corresponding authors: wendy.gilbert{at}yale.edu, caitken{at}vassar.edu
-
Handling editor: Marina Rodnina
Abstract
Translation initiation is a highly regulated process that broadly affects eukaryotic gene expression. Eukaryotic initiation factor 3 (eIF3) is a central player in canonical and alternative pathways for ribosome recruitment. Here, we have investigated how direct binding of eIF3 contributes to the large and regulated differences in protein output conferred by different 5′-untranslated regions (5′ UTRs) of cellular mRNAs. Using an unbiased high-throughput approach to determine the affinity of budding yeast eIF3 for native 5′ UTRs from 4252 genes, we demonstrate that eIF3 binds specifically to a subset of 5′ UTRs that contain a short unstructured binding motif, AMAYAA. eIF3-binding mRNAs have higher ribosome density in growing cells and are preferentially translated under certain stress conditions, supporting the functional relevance of this interaction. Our results reveal a new class of translational enhancers and suggest a mechanism by which changes in core initiation factor activity enact mRNA-specific translation programs.
Keywords
INTRODUCTION
mRNA-specific translational activity—the number of protein molecules produced per mRNA—varies by orders of magnitude under normal growth conditions and is extensively regulated in response to a wide range of physiological signals (Ghazalpour et al. 2011; Schwanhäusser et al. 2011; Vogel and Marcotte 2012; Lahtvee et al. 2017). 5′-untranslated regions (5′ UTRs) directly contact the translation initiation machinery and can strongly influence the rate of translation (Hinnebusch et al. 2016). For example, sequence differences between native yeast 5′ UTRs are sufficient to drive >100-fold differences in translation initiation (Rojas-Duran and Gilbert 2012). Despite great progress toward illuminating the fundamental mechanisms by which eukaryotic mRNAs recruit ribosomes to initiate translation, many quantitatively large mRNA-specific differences in translation initiation remain unexplained.
For most cellular messages, translation initiation requires the concerted action of many eukaryotic initiation factors (eIFs). Cellular mRNAs begin with a 5′-m7G cap that is recognized by the eIF4E subunit of the cap-binding complex, eIF4F. Ribosomes are recruited to mRNA as 43S preinitiation complexes (PIC) that consist of a 40S small ribosomal subunit bound to eIF3, a ternary complex of eIF2•GTP•Met-tRNAi, and additional factors. During cap- and scanning-dependent initiation, the assembled PIC scans from 5′ to 3′ to find the start codon, at which point the 60S large ribosomal subunit joins and protein synthesis begins (Dever et al. 2016). Eukaryotic initiation factor 3 (eIF3) is a central player in this canonical pathway for translation initiation. eIF3 consists of five core subunits that are conserved from yeast to man with seven additional subunits present in filamentous fungi and multicellular eukaryotes (Cate 2017; Valášek et al. 2017). Consistent with its large size and conservation, biochemical and structural studies reveal an extensive network of eIF3 interactions within the PIC, which stabilize the PIC and are required for mRNA recruitment (Aitken et al. 2016; Valášek et al. 2017).
eIF3 is also required for several noncanonical or cap-independent modes of translation. In vitro, eIF3 can alone drive initiation on uncapped mRNAs (Mitchell et al. 2010). Four of the conserved subunits, eIF3a, b, c, and g, contain RNA-binding domains that may bind mRNA as well as 40S ribosomes during initiation (Sun et al. 2013). Consistent with this possibility, crosslinking and immunoprecipitation of eIF3 from human 293T cells identified hundreds of specific crosslink sites mostly within 5′ UTRs (Lee et al. 2015). Further characterization of one eIF3 direct target, c-JUN, showed that eIF3 bound to a structured 5′-UTR element and enhanced cap-dependent translation, although the majority of efficiently translated mRNAs did not crosslink to eIF3. Certain RNA viruses, such as hepatitis C, initiate translation without mRNA caps using structured 5′-UTR elements that bind to host eIF3 with high affinity (Filbin et al. 2009). Thus, direct binding of eIF3 to the 5′ UTR is sufficient to initiate downstream steps in translation initiation. Because this mode of ribosome recruitment is insensitive to regulatory mechanisms that target the activity of the cap-binding complex, cap-independent, and eIF3-dependent initiation is thought to play an important role in selective protein synthesis during times of stress (Gilbert 2010; Shatsky et al. 2018). And yet, how pervasive eIF3 targeting of specific mRNA sequences is and whether it contributes to regulating translation more broadly is not clear, in part because a systematic and quantitative mapping of its targets across the transcriptome does not exist.
Here, we have used an unbiased high-throughput approach to determine the affinity of the yeast eIF3 toward 5′ UTRs from 4252 genes and then compared direct eIF3 binding to translation activity in cells. We incubated purified yeast eIF3 with a synthetic pool of 5′ UTRs at a range of protein concentrations and sequenced the bound RNAs to identify hundreds of specific binders. Quantitative filter-binding assays validated specific 5′ UTRs as high-affinity binders. We identified a sequence motif, AMAYAA, that was significantly enriched within unstructured regions of 5′ UTRs that bound eIF3 and found that mRNAs containing this 5′ UTR motif show higher translation activity in rapidly growing yeast. High-throughput analysis and luciferase reporter assays show that mRNAs which bind eIF3 in vitro maintain higher translation activity under conditions of limiting eIF3 and also during glucose starvation. Together, these results suggest a broad role for direct eIF3 binding to specific 5′-UTR elements during normal and stress-responsive translation.
RESULTS
A high-throughput assay for direct binding of eIF3 to specific 5′ UTRs
Because of the emerging role of eIF3 in mediating the translation of specific transcripts, we asked whether specific yeast mRNAs bind eIF3 with high affinity. Such direct binding to eIF3 could promote efficient translation in growing cells or contribute to selective protein synthesis during stress conditions where cap-binding activity is downregulated. To survey the binding specificity of individual transcripts for eIF3 across the transcriptome, we performed RNA bind-n-seq (RBNS), a quantitative high-throughput assay for RNA-binding affinity in vitro (Lambert et al. 2014). We designed an RNA library of sequences derived from deep sequencing of full-length yeast 5′ UTRs (Pelechano et al. 2014) (Materials and Methods). This pool contained 5′-UTR sequences from 4252 genes and included their annotated alternative 5′-UTR isoforms.
eIF3 was purified from yeast cells and its activity confirmed by biochemical complementation of translation extracts from temperature-sensitive prt1-1 cells lacking functional eIF3 as previously described (Supplemental Fig. S1A,B; Phan et al. 1998). Purified eIF3 at various concentrations (0–1500 nM) was incubated with the 5′-UTR pool, and bound RNA:eIF3 complexes were separated from free RNA by passing through a nitrocellulose filter and used to prepare RNA sequencing libraries together with input RNA (Fig. 1A). For each concentration, we determined an eIF3 enrichment score in the bound fraction as the relative frequency of a given RNA in the bound library compared to the input. The normalized reads for each eIF3 concentration were highly reproducible between replicates (Fig. 1B; Supplemental Fig. S1C). The replicate scores were therefore averaged in all subsequent analyses. Averaged eIF3 enrichment scores from adjacent eIF3 concentrations were highly correlated above 55 nM eIF3 (Spearman R > 0.9, Fig. 1C), which is an indicator of good quality RBNS libraries (Lambert et al. 2014).
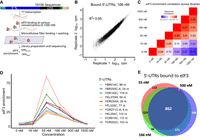
Hundreds of natural yeast 5′-UTRs bind to eIF3 specifically. (A) Scheme of RBNS assay. Designed DNA oligos encoding ∼10,000 yeast 5′-UTRs, plus at least 36 nt of endogenous coding sequence (CDS) are transcribed in vitro. The resulting RNA pool is incubated with various concentrations of purified eIF3, and protein-bound RNAs are captured on nitrocellulose and sequenced. Enrichment is the frequency of a 5′ UTR in the bound sample normalized to input RNA. (B) RBNS results are reproducible. Shown are bound reads in two replicates of 166 nM eIF3. (C) Enrichment between 55 and 500 nM eIF3 was well correlated. (D) Enrichment of the top 10 eIF3 binders from the 166 nM library traced across all libraries. Peak enrichment at 55 nM eIF3 is consistent with high-affinity binding. Less enrichment at higher (eIF3) is expected because low-affinity RNAs consume more of the sequencing lane. (E) Overlap of 5′-UTRs bound in 55, 166, and 500 nM libraries (red, green, and purple, respectively). See also Supplemental Figure S1.
Yeast eIF3 binds hundreds of natural 5′ UTRs with high affinity
Next, we examined which 5′ UTRs are bound with high specificity to eIF3. We focused initially on RNAs that were enriched in the 166 nM eIF3 library, which was selected as a representative of specific binding because this concentration of eIF3 was both intermediate and well correlated with adjacent concentrations (55 and 500 nM) (Fig. 1C). Tracking the 10 most enriched 5′ UTRs from the 166 nM library across all other libraries revealed an enrichment score peak in the 55 nM library, which is consistent with high-affinity binding of these RNAs to eIF3 (Fig. 1D). Reduced enrichment at higher concentrations of protein is expected as lower-affinity binders take up more of the sequencing space (Lambert et al. 2014). 5′ UTRs were defined as “bound” or “not bound” at each concentration of eIF3 using a standard deviation-like cutoff, enrichment > (1 +range 33rd to 66th percentile), as previously described (Taliaferro et al. 2016). 5′ UTRs that bound nonspecifically to nitrocellulose in the absence of eIF3 were eliminated from consideration (Supplemental Fig. S1D). Overall, we identified 1164 5′-UTRs bound in two or more adjacent concentrations of eIF3, with most of these (74%) showing binding at all three intermediate concentrations (55, 166, and 500 nM) (Fig. 1E; Supplemental Table 1).
Representative 5′ UTRs, which included eIF3 binders and nonbinders, were tested by nitrocellulose filter binding with purified eIF3 at concentrations ranging from 15 nM to 1 μM to validate the results from RBNS and determine the Kd of binding (Materials and Methods). Out of 11 tested binders, nine bound to eIF3 with Kd values from 100 to 250 nM and two did not bind tightly (Kd > 1000 nM) (Fig. 2; Supplemental Fig. S2A–G). In parallel, we tested five nonbinding 5′ UTRs (enrichment < 1.59 in 166 nM library) from the RBNS assay, all of which bound weakly or not at all with affinities above the limit for reliable quantification (Kd > 1000 nM) (Supplemental Fig. S2H–L). These results validate specific high-affinity binding of individual targets identified from comprehensive testing of eIF3 binding to yeast 5′ UTRs and show that there are hundreds of different mRNAs whose 5′ UTRs confer preferential eIF3 binding.
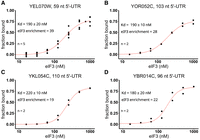
eIF3 binds to specific 5′ UTRs with nanomolar affinity. Purified eIF3 (15–1000 nM) was incubated with individual 32P-labeled 5′ UTRs from YEL070W (A), YOR052C (B), YKL054C (C), and YBR014C (D). Experimental data points (black dots) were fit (red lines) to determine dissociation constants (Kd). Error reflects standard error of the fit of nonlinear regression using Hill's equation. Enrichment score at 166 nM eIF3 is shown for comparison. See also Supplemental Figure S2.
Bound 5′ UTRs are enriched in AMAYAA motifs in unstructured regions
Yeast eIF3 is composed of five distinct subunits, several of which contain known or potential RNA-binding domains, including two RNA recognition motifs (RRM) and two helix–loop–helix (HLH) domains (Valášek et al. 2017), which have been found to mediate recognition of specific RNA sequence motifs by eIF3 and other proteins (Sun et al. 2013; Schuetz et al. 2014). Additionally, certain viral internal ribosomal entry sites (IRES) bind mammalian eIF3 with high affinity by forming an intricate RNA structure (Filbin et al. 2009; Walker et al. 2020). To investigate the mechanisms underlying specific binding of yeast eIF3 to a subset of 5′ UTRs, we first searched the bound RNAs from the 55, 166, and 500 nM libraries for short sequence motifs using DREME (Bailey et al. 2015). All three libraries yielded AMAYAA (where M = A or C and Y = C or U) as the most significantly enriched eIF3 binding motif with additional A-rich sequences identified in each (Supplemental Fig. S3A–C). Therefore, we focused further analysis on the AMAYAA sequence.
If the AMAYAA motif influences eIF3 binding, we reasoned that mRNAs containing more copies of this motif would display higher affinities for eIF3. We counted the motifs in each RNA and divided the pool based on the number of motifs in the sequence. We observed that the presence of the AMAYAA motif in the RNA significantly increases enrichment score in a dose-dependent manner for sequences with 0, 1, 2, or 3+ motifs (P < 1.5×10−5, Fig. 3A). This behavior is similar to other RNA-binding proteins with known binding motifs that were analyzed using RBNS (Taliaferro et al. 2016), suggesting a bona fide sequence preference of eIF3 for AMAYAA. To directly test the contribution of AMAYAA to eIF3 binding, we deleted 2 AMAYAA motifs from a structured 5′ UTR that bound eIF3 with a Kd of 230 nM (YOR092W_51_SL31s, Table 4) and observed at least a fourfold decrease in eIF3 binding (Kd > 1000 nM, Fig. 3B). However, the converse manipulation, mutating a nonbinding YCR054C 5′ UTR (R-score < 1) to introduce an extra AMAYAA motif (C46A, A47C, A49C, G53A), only trended toward increased binding but the difference was not statistically significant (Supplemental Fig. S3J). A single AMAYAA motif may require additional sequence context to significantly alter binding by eIF3.
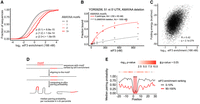
eIF3 recognizes AMAYAA motifs in unstructured regions. (A) eIF3-binding increases with increasing numbers of AMAYAA motifs. Distribution of observed enrichment (166 nM eIF3) for 5′ UTRs with 0, 1, 2, and 3 or more AMAYAA motifs, P-adjusted (Mann–Whitney) for selected pairwise comparisons. (B) eIF3 filter-binding assay of YOR092W_51_SL31s_7 which contains three consecutive AMAYAA motifs (red) and a 10 nt deletion mutant lacking AMAYAA motifs (gray). The best fit of experimental data points (dots) is shown as a line. (C) eIF3 preferentially binds unfolded 5′ UTRs. Enrichment correlates (Pearson) with 5′-UTR folding energies calculated in RNAfold. (D) Overview of RNA motif structure analysis. (E) AMAYAA motifs in tight-binding 5′ UTRs (top decile, red line) are more likely to be presented in unpaired contexts compared to motifs in weak binders (bottom decile, gray line). Median nucleotide pairing probability of 5′ UTRs binned based on their enrichment ranking in the 166 nM library. Position of the motif is indicated by the red rectangle. Benjamini–Hochberg adjusted P-values shown (top) for each nucleotide position. See also Supplemental Figure S3.
Binding of yeast eIF4G to its preferred sequence motif, oligo uridine, is favored when the motif is in an unstructured context (Zinshteyn et al. 2017). We therefore hypothesized that there is a structural difference between RNAs with AMAYAA that readily bind eIF3 and those that do not. To determine the RNA secondary structures of the 5′ UTRs, we performed chemical probing with dimethyl sulfate and analyzed DMS reactivity by sequencing (DMS-MaP-seq) (Zubradt et al. 2017). Globally, eIF3 enrichment was positively correlated with folding energy of the RNA (R = 0.42, P = 2.08 × 10−279, Fig. 3C), which is consistent with preferred binding of eIF3 to unstructured 5′-UTR sequences. To investigate the impact of RNA folding with nucleotide resolution, sequences containing at least one AMAYAA motif were ranked according to their enrichment score in the 166 nM library, aligned by the motif, and folded in silico using RNAfold (Fig. 3D). DMS reactivity was used to constrain 5′-UTR folding to determine the pairing probability for regions of interest. AMAYAA motifs are generally less likely to be paired than flanking sequences, likely due to the lack of G residues. Preferentially bound 5′ UTRs were significantly less folded immediately upstream and downstream from the motif (Fig. 3E). Together, these data are consistent with a preference for yeast eIF3 to bind AMAYAA motifs within unstructured regions of 5′ UTRs. Overall, AMAYAA motifs may contribute to much of the observed binding specificity of purified eIF3, being present in 50.6% of binders (763 in 1508 binders).
eIF3 binders are preferentially translated in growing yeast and maintain higher translation upon glucose starvation
Ribosome footprint profiling and RNA sequencing have identified several 5′-UTR properties that correlate with translation activity genome-wide (Weinberg et al. 2016), but most of the observed variance in ribosome density still cannot be explained by current models. Thus, we asked whether direct eIF3 binding has an impact on translation in cells. Ribosome density, the average number of ribosome-protected footprints normalized to total mRNA levels, is a measure of translation activity that is thought to be predominantly affected by mRNA-specific differences in translation initiation (Shah et al. 2013). We therefore compared ribosome density in exponentially growing yeast to eIF3 binding as approximated by enrichment in the 166 nM library. We restricted this analysis to 2469 genes with a dominant 5′-UTR isoform (Materials and Methods) because ribosome footprints observed within CDS regions cannot be assigned to a specific 5′-UTR isoform and many alternative 5′-UTR isoforms show differential binding to eIF3. In fact, 913 tested genes have isoforms with significantly different (Padj < 0.05) eIF3 enrichment (Supplemental Fig. S4A; Supplemental Table 2).
The mRNAs with 5′ UTRs that bound eIF3 in vitro (Fig. 1E) showed higher ribosome density genome-wide (Fig. 4A). Globally, eIF3 enrichment showed a slight but significant positive correlation with ribosome density (R = 0.18, P = 1.65×10−18) (Fig. 4B; Supplemental Fig. S4B). In addition, mRNAs containing a greater number of AMAYAA motifs display greater ribosome density per mRNA: one motif in the 5′ UTR increased median ribosome density by 20% and two motifs increased it by 49% (Fig. 4C). To directly test the impact of AMAYAA motifs on protein synthesis, we selected three yeast genes with natural 5′-UTR isoform pairs that satisfied the following criteria: (1) one isoform contained an AMAYAA motif not present in the other; (2) the isoform containing the additional AMAYAA motif exhibited significantly higher eIF3 enrichment in vitro (Padj < 0.05, Supplemental Table 2); and (3) the 5′-UTR isoforms were similar in length (±25 nt) (Table 1). We generated nano-luciferase (nLuc) reporters for these UTR pairs expressed from a common beta-estradiol inducible promoter. For the long isoform of YBR014C, which contains two AMAYAA motifs, we additionally generated 5′-UTR variants deleting individual motifs. Loss of a single AMAYAA motif caused a significant decrease in luciferase activity (Fig. 4D). Likewise, for YBR095C, the AMAYAA-containing 5′-UTR isoform that preferentially bound eIF3 in vitro drove significantly higher protein synthesis in cells (Supplemental Fig. S4C). Interestingly, the 5′-UTR isoforms from YDL003W, which also differed significantly in binding to eIF3 in vitro (eIF3 enrichment = 1.2 and 4.48 for shorter and longer variant, respectively, Supplemental Table 2), directed similar levels of protein synthesis in cells (Supplemental Fig. S4C), suggesting that the effect of eIF3 binding to an AMAYAA motif on downstream protein synthesis may be context-dependent.
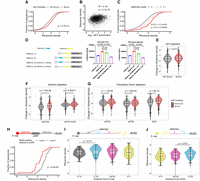
eIF3-binding and AMAYAA motifs enhance ribosome recruitment and start codon recognition in vivo. (A) mRNAs with 5′ UTRs that bound eIF3 in vitro show higher ribosome densities in growing yeast. Cumulative distributions of ribosome densities for bound (red) or not bound (gray) mRNAs. “Bound” includes the union of 55, 166, and 500 nM libraries. Ribosome density equals the average number of ribosome-protected mRNA fragments per mRNA for genes expressing a single dominant 5′-UTR isoform (see Materials and Methods). See Table 2 for ribosome profiling data sources. (B) eIF3 binding in vitro correlates with ribosome density in growing yeast (Pearson). (C) 5′-UTR AMAYAA motifs increase ribosome density in a dose-dependent manner. (D) 5′-UTR AMAYAA motifs promote higher translation of luciferase reporter mRNAs in unstarved conditions (left) and during glucose starvation (right). (E–G) eIF3-binding mRNAs (red) maintain higher ribosome densities than nonbinding (gray) under conditions of limiting eIF3 (E), following acute starvation for glucose or amino acids (F), and upon inactivation of various initiation factors (G). (H) mRNAs with a 5′-UTR AMAYAA motif 5′ to a uORF (red) show reduced ribosome density compared to genes with the motif 3′ (gray). mRNAs were grouped based on the position of the first AMAYAA motif relative to the last uORF. (I,J) Ribosome density varies by AMAYAA motif location for mRNAs without uORFs. Higher ribosome densities for mRNAs with motifs located 11–25 nt from the 5′ end of the mRNA (I) or 0–25 nt upstream of the start codon (J). Bonferroni corrected Mann–Whitney P-values shown for selected comparisons. (A–J) All values of ribosome densities and changes in ribosome densities have been log2-transformed. See also Supplemental Figure S4.
UTR sequences used in luciferase assays
Data sets
Next, we asked whether mRNAs with 5′ UTRs that preferentially bound eIF3 in vitro are able to maintain translation under conditions where eIF3 is limiting. We performed ribosome profiling in tif32-td prt1-td cells in which eIF3 levels were substantially depleted, and bulk translation was reduced by ∼80% (Supplemental Fig. S4E; Jivotovskaya et al. 2006; Stanciu et al. 2022). Because read counts from ribosome profiling experiments are normalized to total reads from each library, TE changes within each sample reflect the translational status of individual mRNAs relative to the distribution of TE values observed for all mRNAs within that sample, which can be highly skewed by large changes to highly expressed genes. Thus, these relative TE changes cannot be compared in absolute terms between samples, as evidenced by the observation of positive TE changes in eIF3-depleted cells.
The mRNAs that we observed to bind eIF3 in our RBNS experiments display greater ribosome occupancy upon eIF3 complex depletion (Stanciu et al. 2022). This counterintuitive observation likely originates from the fact that, unlike a true deletion mutant, eIF3 concentrations are quickly depleted but not eliminated in tif32-td prt1-td cells growing under restrictive conditions (Fig. 4E). Under these eIF3-deplected conditions, mRNAs containing AMAYAA motifs likely outcompete other mRNAs.
Interestingly, the apparent translational advantage of eIF3-binding mRNAs persisted in cells expressing a mutant form of eIF3i, tif34DDKK, which disrupts the eIF3i:eIF3b binding interface resulting in the loss of both eIF3i and eIF3g from the eIF3 complex (Supplemental Figs. S4F and S4G; Herrmannová et al. 2012). This result suggests that AMAYAA-containing mRNAs might bind eIF3 via interactions with the eIF3a/b/c subunits remaining in the complex or can productively interact with eIF3i or eIF3g outside the context of the full eIF3 complex. Either of these potential scenarios raise the possibility that regulation of individual yeast eIF3 subunits could mediate mRNA-specific translational control, consistent with a previous report tying individual subunits of human eIF3 to differential translation outcomes (Lamper et al. 2020).
We then examined translation activity of eIF3-binding mRNAs under various stress conditions where alternate mechanisms of ribosome recruitment may contribute to selective translation of some genes. We found that mRNAs that are capable of direct eIF3 binding are significantly more resistant to downregulation of translation during acute glucose starvation (P = 3.7×10−8) (Fig. 4F, left; Zid and O'Shea 2014) and amino acid withdrawal (Fig. 4F, right; Santos et al. 2019). To test the impact of AMAYAA motifs during nutrient deprivation, we repeated the luciferase reporter experiments following acute glucose starvation. To ensure that we measured luciferase protein produced de novo during starvation, these reporters contained an IsoE degron fused to the C-terminus of nLuc. IsoE fusion leads to rapid degradation of tagged proteins (Peter et al. 2021). For two out of the three isoform pairs we tested, we observed significantly higher translation during glucose starvation for the isoform whose 5′ UTR contains an additional AMAYAA motif (Fig. 4D; Supplemental Fig. S4C, right panel). Deletion of individual AMAYAA motifs from YBR014C was sufficient to reduce translation in starved cells (Fig. 4D, right panel). However, for YDL003W the longer AMAYAA-containing isoform was not better translated in glucose-starved cells (Supplemental Fig. S4C), again suggesting that the effects of the AMAYAA motif on eIF3 binding and downstream translational outcomes might be context-dependent or be modulated by other unknown factors.
We speculate that stresses that downregulate early steps in initiation (e.g., cap-binding activity) allow selective, eIF3-dependent translation of specific mRNAs. Consistent with this possibility, eIF3-binding mRNAs maintained higher translation in cells genetically depleted of eIF4G (P = 8.94×10−6) as well as in cold-sensitive mutants of factors that collaborate with eIF4G to recruit ribosomes, including Ded1 (ded1-cs, P = 8.9×10−13) and eIF4B (tif3-cs, P = 8.8×10−7) (Fig. 4G; Sen et al. 2015, 2016; Zinshteyn et al. 2017). In each of these cases, AMAYAA-containing mRNAs likely outcompete AMAYAA-lacking mRNAs for eIF3 and the translational machinery, thus enabling them to escape the global repression of translation experienced by other mRNAs.
eIF3-binding motifs can promote or repress translation depending on their location
Our analysis of eIF3 binding in vitro and ribosome profiling in vivo is consistent with widespread translational enhancement via eIF3 binding to 5′ UTRs. Focusing on the AMAYAA binding motif, we expanded our analysis to include all genes with a single dominant 5′ UTR and sufficient reads to quantify ribosome density in exponentially growing yeast. This allowed us to include an additional 536 genes, many of which contained multiple upstream open reading frames (uORFs). Intriguingly, while the AMAYAA motif was associated with enhanced translation if the longer 5′ UTR lacked uORFs, in mRNAs with multiple uORFs AMAYAA motifs were associated with translational repression of the main ORF (Supplemental Fig. S4H).
We hypothesized that upstream eIF3-binding motifs enhance translation of uORFs, which leads to fewer ribosomes initiating translation of the main ORF. Additionally, as ribosomes sense the 5′-UTR sequence by scanning, we hypothesized that the order of the motif and the uORF on the 5′ UTR will matter. Therefore, we examined ribosome density on 78 mRNAs that contain one uORF and one AMAYAA motif within the 5′ UTR. Genes were separated into two categories based on the relative positions of uORF and motif: motif 5′ of the uORF or motif 3′. Notably, there was a twofold difference in ribosome density between mRNAs with the motif upstream versus downstream from uORFs, with median ribosome densities of 0.24 and 0.55, respectively (P = 2.7×10−8) (Fig. 4H). Therefore, it is likely that binding of eIF3 to AMAYAA motifs promotes recognition of downstream AUG codons, which can be in uORFs or the main ORF.
eIF3 binding to AMAYAA motifs has the potential to enhance translation initiation by multiple mechanisms, which include initial recruitment of 43S PIC as well as recognition of initiation codons (AUGi) during scanning. The most likely mechanism depends on the position of the AMAYAA motif relative to the cap and AUGi. We therefore compared ribosome density among groups of genes with AMAYAA motifs at varying distances from the annotated transcriptional start site (TSS) and from the AUGi, excluding genes with uAUGs. Ribosome density was highest for mRNAs where the motif is within 11–25 nt from the TSS (Fig. 4I), or within 25 nt upstream of the AUGi (Fig. 4J). In contrast to the apparent stimulatory effect of AMAYAA located 11–25 nt downstream from the 5′ end, we observed lower ribosome density for mRNAs with motifs located within 10 nt of the 5′ end. This positional difference is consistent with the potential for occlusion of cap-proximal motifs by the eIF4F complex, preventing eIF3 from recognizing them. Together, our results indicate that high-affinity binding interactions between yeast eIF3 and specific 5′-UTR sequences increase translation initiation at downstream start codons, and may do so by promoting distinct steps depending on the site of eIF3 binding (Fig. 5).
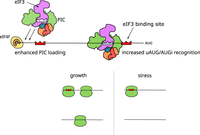
Model of location-dependent translational enhancement by direct binding of eIF3 to 5′-UTR motifs. eIF3 binding to AMAYAA motifs may enhance translation initiation by multiple mechanisms depending on the location of the motif. (Top left) AMAYAA motif binding to eIF3 increases initial recruitment of 43S PIC. (Top right) AMAYAA motif upstream of initiation codons (AUGi) increases their recognition during scanning. The presence of AMAYAA motifs increases ribosome loading during growth (left) and allows some mRNAs to maintain ribosome loading during starvation stress (right).
DISCUSSION
5′ UTRs are the site of action during translation initiation and are an important mRNA feature for controlling protein production posttranscriptionally in eukaryotic cells. We hypothesize that translation-enhancing elements within cellular 5′ UTRs include sequences that bind preferentially to multiple eIFs, as recently demonstrated for yeast eIF4G1 (Zinshteyn et al. 2017). Here we performed an unbiased testing of purified eIF3 binding to a library of thousands of yeast 5′ UTRs to uncover hundreds of mRNAs that preferentially bind eIF3. We show that eIF3-binding mRNAs have higher translation activity in growing cells and are less sensitive to translational inhibition in response to a variety of genetic and environmental perturbations that result in widespread effects on cellular translation. Together, our findings support a broad role for high-affinity interactions between cellular 5′ UTRs and core initiation factors for translational control of gene expression.
eIF3 promotes translation initiation by multiple mechanisms which include selective ribosome recruitment to viral mRNAs that bind eIF3 with high affinity (Filbin et al. 2009; Valášek et al. 2017). Our results suggest a similar role for eIF3 interactions in cellular mRNA selectivity—preferential translation of certain mRNAs under conditions that limit global initiation activity or favor noncanonical pathways of ribosome recruitment. In support, yeast eIF3-binding mRNAs maintained higher ribosome densities (average number of ribosomes per mRNA) when eIF3 protein was limited by depletion in the eIF3a/b degron strain, when cap-dependent translation was inhibited by depletion of eIF4G, and when Ded1 or eIF4B were inactivated by cold-sensitive mutations (Fig. 4E,G). Globally, eIF3 binding to 5′ UTRs in vitro was modestly but significantly correlated with ribosome density in rapidly growing cells (Fig. 4B; Supplemental Fig. S4B). This result was surprising and suggests that binding to eIF4F is not the limiting factor for initiation on all mRNAs despite the fact that eIF4F is the least abundant initiation factor during exponential growth in rich media (von der Haar and McCarthy 2002). Previous work established a requirement for eIF3 for RPL41A mRNA association with PICs in vivo (Jivotovskaya et al. 2006). Our findings suggest that quantitative differences in eIF3 binding partially explain differences in PIC recruitment to different mRNAs in cells.
Analysis of eIF3-binding RNAs supports the existence of at least two modes of high-affinity binding to cellular 5′ UTRs, one that is sequence-dependent and another that remains to be determined. Binding to eIF3 in vitro (Fig. 3D) and ribosome density in vivo (Fig. 4C) increased with increasing numbers of AMAYAA motifs within the 5′ UTR unless the motif was located upstream of a uAUG (Fig. 4H). This context-dependent effect of the eIF3-binding motif on translation of the main ORF is consistent with a simple model whereby binding to eIF3 promotes initiation on the closest downstream AUG. The observed optimal spacing with respect to the 5′ end of the mRNA is consistent with enhanced 43S recruitment immediately downstream from the cap-proximal region bound by eIF4F (Fig. 4I). In addition, translational enhancement by AMAYAA motifs close to and upstream of AUGi suggests favorable interactions with 43S-bound eIF3 during start codon recognition (Fig. 4J). This spacing is consistent with crosslinks observed between conserved subunits of mammalian eIF3, a and b, and the mRNA at positions −17 to −14 relative to the start codon (Pisarev et al. 2008).
Our results raise the question of which parts of the multiprotein eIF3 complex are responsible for high-affinity binding to specific cellular 5′ UTRs. It is likely that distinct eIF3 surfaces contribute to binding in different cellular 5′ UTRs as shown for two classes of eIF3-binding viral 5′ UTRs (Neupane et al. 2020). Multiple conserved subunits of human eIF3, including a, b, d, and g, crosslink directly to cellular mRNA in cultured human cells (Lee et al. 2015), highlighting the potential for distinct subunit binding preferences to contribute to mRNA selection. In addition, direct binding of mRNA caps to mammalian eIF3d supports translation during glucose deprivation (Lamper et al. 2020). The cap-binding eIF3d subunit is not conserved in budding yeast. It will be interesting to see whether binding to AMAYAA motifs is conserved in human eIF3. Candidates to mediate recognition of the AMAYAA sequence include the HLH motifs in eIF3a and eIF3c and the RRM in eIF3b. eIF3g, which also contains an RRM, appears to be dispensable for the preferential translation of many eIF3-binding mRNAs in vivo based on the observation that eIF3-binding mRNAs maintain high ribosome density in tif34DDKK mutants (Supplemental Fig. S4G) in which eIF3g and eIF3i are destabilized from the core complex of eIF3a/b/c (Herrmannová et al. 2012). It is possible that eIF5 or eIF1 contribute to sequence-specific binding. These proteins associate with eIF3 in vivo and may be present in purified eIF3 at substoichiometric amounts. Our results also suggest the potential for mRNA-specific translational regulation by posttranslational modifications to specific domains within eIF3, such as succinylation of the RRM in eIF3b or phosphorylation of the RRM in eIF3g (Albuquerque et al. 2008; Weinert et al. 2013).
The comprehensive approach used here to identify cellular 5′ UTRs that bind to wild-type yeast eIF3 can be used to tease out the contributions of individual domains, and even specific amino acids of interest. It is increasingly clear that core initiation factors engage cellular 5′ UTRs in highly specific interactions that contribute to mRNA-specific rates of initiation—in growing cells and global retuning of translation during stress. Our results suggest a broad potential for direct eIF3 binding to maintain the translation of specific cellular mRNAs during a variety of stress responses. Such mRNA-specific sensitivities are likely to be important for the pathological effects of dysregulated initiation factors in cancer and other human diseases.
MATERIALS AND METHODS
Yeast strains and growth
Genotypes are listed in Table 3. Strain LPY87 was grown for eIF3 purification as previously described (Phan et al. 2001). Briefly, culture was grown overnight in synthetic complete (SC) media without leucine and uracil at 30°C. The starter culture was used to inoculate 18 L of YPD media and grown for 14–18 h to OD600 4–5. Wild type (PRT1) and eIF3 temperature-sensitive (prt1-1) strains used to prepare extracts for in vitro translation, and complementation assays were grown at 23°C in YPD. The eIF3a/b degron strain YAJ34 (Jivotovskaya et al. 2006) was grown at 25°C in SCRaff + Cu2+ before being shifted to prewarmed SCRaff/Gal + BCS at 36°C for 90 min to deplete eIF3a/b. The temperature-sensitive tif34-DDKK strain (Herrmannová et al. 2012) was grown at 30°C in SC media before being shifted to prewarmed SC media at 37°C for 30 min.
Yeast strains
eIF3 purification
LPY87 cells expressing His-tagged eIF3 were harvested (85–95 g of wet cell pellet), washed with lysis buffer (20 mM HEPES.KOH, 350 mM KCl, 5 mM MgCl2, 10% glycerol, 20 mM imidazole, 10 mM β-mercaptoethanol, pH = 7.4), resuspended in 30 mL of lysis buffer, frozen as droplets in liquid N2 and stored in −80°C until lysis on a Retsch CryoMill, using 2 × 5 min shaking at 15 Hz with 1 min intermittent cooling at 5 Hz. Cell powder was thawed in 400 mL of lysis buffer in the presence of cOmplete Protease Inhibitors (Sigma-Aldrich), 1 µg/mL pepstatin A, 1 µg/mL aprotinin, 1 µg/mL leupeptin, and 100 µL of Turbo DNase. Lysate was clarified at 12,500 × g for 40 min and applied to 5 mL of freshly regenerated Ni2+ Sepharose. Bound protein was washed with lysis buffer until there was no detectable protein in the flowthrough and eluted using lysis buffer supplemented with 350 mM imidazole. Fractions containing eIF3 were concentrated with 10 kDa MWCO centrifugation columns to ∼ 2 mL and resolved in two batches on HiLoad 16/60 Superdex 200 preequilibrated in low salt buffer 20 mM HEPES.KOH, 100 mM KCl, 10% glycerol, 0.1 mM EDTA, 2 mM DTT, pH = 7.4. Fractions containing all five eIF3 subunits were combined and loaded on a phosphocellulose column prepared from cellulose phosphate (Sigma-Aldrich) sequentially activated in 1M HCl, 1 M NaOH, and preequilibrated in low salt buffer. The column was then washed with 100 mL of low salt buffer and the protein was sequentially eluted with 200/350/1000 mM KCl in 20 mM HEPES.KOH, 10% glycerol, 0.1 mM EDTA, 2 mM DTT, pH = 7.4. eIF3-containing fractions were pooled, dialyzed against 2 L of storage buffer (20 mM HEPES.KOH, 100 mM KOAc, 10% glycerol, 2 mM DTT, pH = 7.4), concentrated to 0.5–1 mL, and stored in 15 µL aliquots in −80°C. Final eIF3 concentration was determined using Bradford assay with BSA as the protein standard.
eIF3 complementation assay
Translationally active extracts were prepared from wild type (PRT1) and heat-sensitive eIF3 mutant (prt1-1) strains grown at 23°C using published protocols (Zinshteyn et al. 2017). Three replicate extracts prepared from independent cultures were either pretreated at 39°C for 10 min or kept on ice. Translation of capped nanoluciferase reporter mRNA was performed with supplemental 100 nM eIF3 or buffer control in three technical replicates as described previously (Rojas-Duran and Gilbert 2012). The reaction was stopped after 30 min by 100-fold dilution into PBS, and the amount of nanoluciferase was measured using Nano-Glo (Promega) on a Centro XS3 Luminometer (Berthold).
5′-UTR pool design and synthesis
5′-UTR boundaries and abundances were calculated from sequencing of wild-type yeast (Pelechano et al. 2014). When a 5′ UTR started within 10 nt of its nearest neighbor, the sequences were merged. Inclusion in the pool also required the following: 5′ UTRs must be expressed within 25% of the mode abundance for a given 5′ UTR, and 5′ UTRs must make up at least 5% of the total abundance for that ORF, unless the mode was <5% of the total, in which case we used the mode. Upstream AUGs within 761 (6.3% of all) 5′-UTR sequences were mutated to AGT, such that the first AUG encountered by a scanning PIC moving 5′ to 3′ would be the annotated AUGi. For technical reasons, the synthetic pool used for eIF3 RBNS was limited to 5′ UTRs ≤ 122 nt. Each sequence consisted of a randomized 10 nt unique identifier barcode and an adaptor sequence used for priming reverse transcription and Illumina sequencing. RNA was in vitro transcribed from the PCR-amplified pool using in house prepared T7 RNA polymerase and gel-purified.
RBNS library preparation
Binding reactions (50 µL) were assembled at room temperature with 60 ng/µL of pool RNA (∼1 µM) and various concentrations of eIF3 (0–1500 nM, two replicates each) in 20 mM HEPES. KOH, 100 mM KOAc, 2 mM Mg(OAc)2, 10% glycerol, 2 mM DTT, RNasin (80 U, Promega), and 100 ng/µL of yeast tRNA (∼4 µM, Sigma-Aldrich). Binding reactions were incubated for 30 min (Mitchell et al. 2010), after which the reaction was passed through layered nitrocellulose (top) and nylon (bottom) filters preequilibrated in binding buffer using a vacuum manifold. Filters were washed 2 × 200 µL with ice-cold binding buffer and dried. eIF3-bound RNA was extracted from nitrocellulose by incubation with proteinase K (Sigma-Aldrich), concentrated by Zymo column, and reverse transcribed with AMV reverse transcriptase (RT primer: OWG921, Table 5). Gel-purified cDNA was ligated with a barcoded (10N) 5′-adapter as previously described (Niederer et al. 2022).
RNA sequences used for validating effects of AMAYAA motif in filter-binding assays
Oligonucleotides used in library construction
Ribosome profiling of eIF3a/b degron and eIF3i-DDKK
Each strain and its matching wild type (Table 3) were grown under restrictive conditions (see “Yeast strains and growth” section) for a duration resulting in an ∼80% decrease in bulk translation as judged by analytical polysome profiling and quantification of polysome:monosome ratios, as previously described (Jivotovskaya et al. 2006). Cycloheximide was added to a final concentration of 100 µg/mL 2 min prior to harvesting by filtration through a Kontes filtration apparatus and flash freezing in liquid nitrogen. We performed all subsequent steps as described previously (Sen et al. 2016).
Data analysis
RBNS raw sequencing reads were trimmed with cutadapt and aligned to the pool sequences. PCR duplicates in each library were removed using 10N barcodes in the 5′-adapter, and reads were normalized in each library according to their sequencing depth. Artificial sequences designed for other studies were excluded from the data processing and analysis (Niederer et al. 2022). For quantification, we required >1 rpm in each of the input replicates and >2 reads in each of the sample libraries. The enrichment score for each 5′ UTR in a given library was then calculated as the ratio of normalized reads in the library to input. Data were processed and visualized in Python and R using custom scripts.
For comparisons between translation activity in vivo and eIF3 binding in vitro, in vivo 5′ UTRs were defined based on sequencing reads (Pelechano et al. 2014). For every ORF, 5′ UTRs within 10 nt of one another were merged. Only 5′ UTRs that contributed at least 5% to the total mRNA pool for a given gene were considered. Dominant 5′ UTRs were defined as the most abundant 5′ UTR for a given gene, which must account for at least 40% of all mRNAs for that gene and be at least twice as abundant as the second most common 5′ UTR. The presence of an upstream AUG was the selection criteria for assigning uORFs to a given 5′ UTR. For the motif positional comparisons, only 5′ UTRs with sizes between 40 and 150 nt were selected.
Filter binding
Individual 5′ UTRs were amplified from the pool using target-specific primers and in vitro transcribed. Gel-purified RNA was dephosphorylated (FastAP, Thermo Fisher) and 5′-labeled with 32P-γ-ATP (PerkinElmer) using T4 polynucleotide kinase (NEB). Binding reactions (5 µL) contained ∼10 nM labeled RNA and 0/16/32/63/125/250/500/1000 nM eIF3 in buffer as described for RBNS. After 30 min, reactions were passed through layered nitrocellulose (top) and nylon (bottom) filters preequilibrated in binding buffer. Filters were washed with 80 µL of ice-cold binding buffer, disassembled, and dried. Captured RNA was visualized by phosphorimaging and quantified using ImageJ (Schneider et al. 2012). At each concentration, eIF3-bound RNA was quantified as the ratio of nitrocellulose-bound RNA to the sum of RNA captured on both filters. Data were fit using nonlinear regression with Hill's equation in Origin.
Yeast luciferase assays
To measure 5′-UTR-dependent translation activity in vivo, 5′-UTR sequences (Table 1) generated as gene fragments (Twist Bioscience) were cloned into a β-estradiol inducible expression vector pKP3915 (Addgene) in front of nanoluciferase (nLuc). YWG506 yeast strain was transformed with 5′-UTR-nLuc plasmids and grown to log phase in synthetic media containing 2% glucose and lacking uracil (SD-Ura +2% glucose). Cells were pelleted and resuspended either in SD-Ura with glucose (SD-Ura +2% glucose) or without glucose media (SC-Ura +0% glucose). mRNA expression was induced by the addition of β-estradiol (Sigma E1024-25G) to a final concentration of 50 nM for 20 min at 30°C. Whole-cell lysates were prepared by vortexing with glass beads in 1 × PBS with protease inhibitors (2 mM phenylmethanesulfonyl fluoride), 1 × protease inhibitor cocktail (Roche). Lysate was diluted 1:10 with 1× passive lysis buffer (Promega), and luciferase activity was monitored by using the NanoGlo luciferase assay reagent (Promega) on a Promega GloMax Luminometer. Luciferase values were normalized to total protein measured by BCA assay.
DMS-MaPseq library preparation
DNA pool was amplified with PCR (forward primer oRN07; reverse primer oRN08). RNA was produced by T7 in vitro transcription using the MEGAshortscript kit (Thermo Fisher Scientific AM1354), treated with TURBO DNase (Invitrogen), and gel-purified. For DMS probing, RNA (2 μg in 6 μL water) was denatured at 95°C for 2 min. RNA was refolded by adding 88.8 μL 300 mM sodium cacodylate and 1 μL RNasin Plus (Promega N2611) and 10 min incubation at 30°C, followed by addition of 1.2 μL 500 mM MgCl2 and 20 min incubation at 30°C. DMS (3 μL, Sigma-Aldrich D186309) was added to the folded RNA solution and allowed for 2 min incubation at 30°C, then quenched by adding 42.8 μL β-mercaptoethanol (BME). For denatured RNA control, RNA (2 μg in 6 μL water) was mixed with 1 μL RNasin Plus, 39.2 μL water, 50 μL formamide (Invitrogen 15515-026) and 0.8 μL 0.5 M EDTA and incubated at 95°C for 2 min. The solution was incubated with 3 μL DMS and quenched with 42.8 μL BME. RNAs were purified via ethanol precipitation and reverse-transcribed using TGIRT-III (InGex) and primer oRN08. The resulting cDNA was gel purified and a 5′ adaptor (oWG920) was ligated. Libraries were prepared as described above and sequenced via NovaSeq.
DMS-MaPseq analysis
Paired-end sequencing reads were first merged using BBMerge and PCR duplicates were removed with Dedupe. BBDuk was used to remove adaptors and trim the 10N UMI sequences. Reads were then aligned with STAR using the following parameters: ‐‐ alignSJoverhangMin 999, ‐‐ alignIntronMax 1, ‐‐ alignIntronMin 999. Mismatches to the reference sequence were labeled with MD tags using the SAMtools calmd function. The coverage and mutation rate for each position were calculated and averaged within replicates using a house-developed script. The sequences with a coverage of < 400 reads were removed from further analysis. For each sequence, the mutation rates on A/C positions were normalized to a 0-1 scale, while U/G positions were not considered. The mutation rates were used to predict base-pairing probability using RNAfold from the ViennaRNA package.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
We thank Alan Hinnebusch and Jon Lorsch for the eIF3 production strain and eIF3 purification protocols. We thank members of the Gilbert laboratory for discussion and comments on the manuscript. This work was supported by National Institutes of Health (NIH) R01GM132358 to W.G. and R15GM140372 to C.E.A. R.O.N. was supported by the American Cancer Society (Postdoctoral Fellow) and the NIH (K99GM135533).
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080310.124.
- Received October 10, 2024.
- Accepted November 4, 2024.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵Phan L, Schoenfeld LW, Valášek L, Nielsen KH, Hinnebusch AG. 2001. A subcomplex of three eIF3 subunits binds eIF1 and eIF5 and stimulates ribosome binding of mRNA and
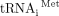 . EMBO J 20: 2954–2965. doi:10.1093/emboj/20.11.2954
. EMBO J 20: 2954–2965. doi:10.1093/emboj/20.11.2954 - ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
MEET THE FIRST AUTHOR

Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Jiří Koubek is the first author of this paper, “Cellular translational enhancer elements that recruit eukaryotic initiation factor 3.” Jiří obtained his PhD in 2014 from the Taiwan International Graduate Program (TIGP) in Academia Sinica, Taiwan, under the supervision of Joseph Jen-Tse Huang, studying cotranslational folding using fluorescence anisotropy. Jiří then joined Wendy Gilbert's lab, where he worked on the subject of this paper. Currently, Jiří is in the Bukau lab in ZMBH, Heidelberg University, applying ribosome profiling and cryoEM to reveal the mechanisms of cotranslational assembly.
What are the major results described in your paper and how do they impact this branch of the field?
In my opinion, the major results described in the paper is that eIF3 has a binding motif that is fairly frequent in the UTR. It is probably no surprise that eIF3 has a binding specificity; after all, it is wrapped around the ribosomal small subunit during translation initiation. Nevertheless, when we looked into the impact of the motif on translation, we could show that the presence of the motif leads to a higher translational output, suggesting that eIF3 might play a role in translation initiation rates. However, things are not as simple as they seem—the motif and upstream ORF leads to lower ribosome density on the main ORF. While we provide a plausible explanation, I think a proper follow-up study is worthwhile to do.
What led you to study RNA or this aspect of RNA science?
This came about very naturally. When I started the lab rotation in Joseph Huang's lab, I supported a project on folding on the ribosome using fluorescence spectroscopy. I wanted to develop the idea, trying to map an entire pathway so working with ribosomes was a prerequisite. In addition, I had to have a system which would incorporate fluorophores in a site-specific manner, so that's how I had to get my hands dirty with tRNA.
If you were able to give one piece of advice to your younger self, what would that be?
Oh, there are so many. There have been multiple lessons that I have learnt really hard in the last 10 years which would have been very useful to know 10 years ago. But probably the most critical (since I can pick only one) would be—don't rush with your PhD, there is no reason to graduate too early. Academia is a very competitive environment, and the moment one graduates is when the real academic clock starts, which impacts everything—from postdoc fellowships to tenure track positions openings. However, as I know myself all too well, I would have probably ignored this advice.
Are there specific individuals or groups who have influenced your philosophy or approach to science?
There were multiple people along my career path that had a profound impact on my approach to science, each in a different time period. During my stay in Academia Sinica, I have to mention three people—my advisor Dr. Joseph Jen-Tse Huang, who allowed me to follow my curiosity and do something completely unrelated within his group; Dr. Tien-Hsien Chang, who has deeply imprinted in me his approach to teaching and proposal writing; and Dr. Sunney I. Chan, who has always shared his wisdom, which resonates with me even now.
Moreover, I have also been deeply impacted by people who supervised me (and are still supervising me) during my postdoc. I have grown a lot as a scientist in Dr. Wendy Gilbert's laboratory which included switching fields and exploring new methodologies (results of my work are now published here). Some of the lessons that I have learnt were hard, but that I take as an important part of the learning process. My current advisor Dr. Bernd Bukau and co-advisor Dr. Guenter Kramer have helped me develop the skills that I was lacking so far, and thanks to our close collaborator, Dr. Stefan Pfeffer, I have had a chance to expand my horizons to cryoEM.
Finally, I cannot neglect the influence of my wife Tracy, who is also a scientist. She has been with me the entire time, and through our daily discussion I can get a completely different perspective. I do not necessarily need to agree with everything, but she provides me a daily peer review of my science.
What are your subsequent near- or long-term career plans?
My long-term goal has always been to stay in academia and lead my own research group. Everybody who goes through this path knows that there are serious obstacles along the way due to the high competition. So I'll be developing my own research project and applying for positions. Whether I will succeed depends on my nearest plan—getting the study from my current lab published.








