
2′-O-methylation (Nm) in RNA: progress, challenges, and future directions
- 1Division of Medical Oncology, Department of Medicine, Duke University, Durham, North Carolina 27710, USA
- 2UNC Lineberger Comprehensive Cancer Center, University of North Carolina at Chapel Hill, Chapel Hill, North Carolina 27599, USA
- 3Division of Hematology and Oncology, University of North Carolina at Chapel Hill, Chapel Hill, North Carolina 27514, USA
- 4University of North Carolina RNA Discovery Center, UNC Lineberger Comprehensive Cancer Center, University of North Carolina at Chapel Hill, Chapel Hill, North Carolina 27599, USA
- 5Division of Cardiology, Department of Medicine, Duke University, Durham, North Carolina 27710, USA
- Corresponding author: cholley{at}duke.edu
Abstract
RNA 2′-O-methylation (Nm) is highly abundant in noncoding RNAs including ribosomal RNA (rRNA), transfer RNA (tRNA), and small nuclear RNA (snRNA), and occurs in the 5′ cap of virtually all messenger RNAs (mRNAs) in higher eukaryotes. More recently, Nm has also been reported to occur at internal sites in mRNA. High-throughput methods have been developed for the transcriptome-wide detection of Nm. However, these methods have mostly been applied to abundant RNAs such as rRNA, and the validity of the internal mRNA Nm sites detected with these approaches remains controversial. Nonetheless, Nm in both coding and noncoding RNAs has been demonstrated to impact cellular processes, including translation and splicing. In addition, Nm modifications at the 5′ cap and possibly at internal sites in mRNA serve to prevent the binding of nucleic acid sensors, thus preventing the activation of the innate immune response by self-mRNAs. Finally, Nm has been implicated in a variety of diseases including cancer, cardiovascular diseases, and neurologic syndromes. In this review, we discuss current challenges in determining the distribution, regulation, function, and disease relevance of Nm, as well as potential future directions for the field.
Keywords
INTRODUCTION
Methylation of the 2′-hydroxyl group of the ribose sugar of a nucleotide, or 2′-O-methylation (Nm), is an abundant posttranscriptional modification found in diverse classes of noncoding and coding RNAs (Fig. 1; Höfler and Carlomagno 2020). Nm is highly abundant in noncoding RNAs, especially ribosomal RNA (rRNA), transfer RNA (tRNA), and small nuclear RNA (snRNA) (Maden 1990; Krogh et al. 2017; Marchand et al. 2017). In addition, Nm is an essential feature of the 5′ cap of eukaryotic messenger RNA (mRNA) and a characteristic feature of the 3′ end of Piwi-interacting RNA (piRNA) in animals and microRNA (miRNA) in plants (Yu et al. 2005; Saito et al. 2007; Galloway and Cowling 2019). More recently, several reports have described internal Nm modifications in mRNA, with evidence of physiologic relevance despite seemingly low stoichiometry of modification (Dai et al. 2017; Bartoli et al. 2018; Elliott and Holley 2021; Leighton et al. 2022; Tang et al. 2023). However, the accurate mapping of internal Nm in mRNA remains a challenge, and the true abundance, distribution, and physiologic relevance of internal Nm in mRNA remain controversial (Motorin and Marchand 2018). The stoichiometry of Nm in noncoding RNAs varies in different physiologic and pathologic states, and the modulation of Nm levels has functional consequences in diverse processes including translation, splicing, and innate immunity (Hyde and Diamond 2015; Bohnsack and Sloan 2018; Monaco et al. 2018). Currently, our understanding of the regulation of Nm is limited by methodological challenges in measuring and manipulating Nm levels at specific sites. Dysregulation of Nm has been noted in a variety of human diseases, but much of this data is correlative, and the contribution of Nm to disease pathogenesis remains unclear (Dimitrova et al. 2019).

Nm in cellular RNAs. (A) Nm refers to 2′-O-methylation of the ribose moiety of a ribonucleotide N. (B) Nm occurs in noncoding RNAs including rRNA, tRNA, and snRNA, as well as at the 3′ end of miRNAs and piRNAs. (C) Nm also occurs in the mRNA 5′ cap and more recently has been reported to occur at internal sites in mRNA. Created with BioRender.com.
In this review, we aim to provide an overview of Nm modifications in RNA, focusing on what we view as key issues that are either controversial or largely unresolved: (i) Mapping and measurement—where is Nm located in RNA, and how much? (ii) Regulation—when and where does Nm change, and what regulates Nm levels? (iii) Functional implications—how does Nm alter RNA function and fate? and (iv) Disease relevance—how is Nm important to health and disease?
MAPPING AND MEASUREMENT OF Nm
Although there are no reported antibodies that selectively bind to Nm, the 2′-O-methylation of ribonucleotides confers several chemical properties that can be exploited for the site-specific mapping of Nm in RNA, including resistance to alkaline hydrolysis, periodate oxidation, and enzymatic cleavage, as well as steric hindrance during reverse transcription (RT) (Table 1; Motorin and Marchand 2018). One commonly used method for the high-throughput mapping of Nm is RiboMeth-seq, which detects Nm sites as gaps in 5′- and 3′-end coverage indicating resistance to alkaline hydrolysis (Birkedal et al. 2015; Krogh et al. 2016; Marchand et al. 2016). In contrast, RibOxi-seq and Nm-seq detect Nm sites as 3′-end peaks after the treatment of fragmented RNA with multiple cycles of periodate oxidation (Dai et al. 2017; Zhu et al. 2017). NJU-seq takes advantage of the resistance of Nm to cleavage by Mycoplasma genitalium RNase R (MgR) (Tang et al. 2024). Methods exploiting the steric properties of Nm include 2OMe-seq and MeTH-seq, which detect the termination of RT at Nm sites under restrictive conditions with either low deoxynucleotide triphosphate (dNTP) or low magnesium concentrations (Incarnato et al. 2017; Bartoli et al. 2018), and Nm-Mut-seq, which uses an engineered reverse transcriptase to identify Nm sites as base mutations in the RT step, but is unable to detect Um (Chen et al. 2023). Emerging reports suggest that nanopore sequencing can map Nm at ±2 nt resolution based on Nm-induced changes in the current signal (Stephenson et al. 2022). Despite these many approaches that accurately detect Nm sites on highly abundant rRNA and should theoretically enable site-specific detection of Nm on mRNA, there is little to no agreement between the methods as to specific mRNA sites that are subject to Nm modification.
High-throughput methods for site-specific mapping of Nm sites in RNA
This inability to conclusively map Nm sites on mRNA likely relates in part to the fact that mRNA Nm sites are incompletely methylated. Of the high-throughput methods for Nm mapping, only RiboMeth-seq has been shown to quantitatively measure absolute Nm fraction, yet it is impractical for application to mRNA due to the high depth of sequencing required (Marchand et al. 2016; Motorin and Marchand 2018). For Nm sites in rRNA, the stoichiometry of Nm determined by RiboMeth-seq correlates well with the Nm stoichiometry measured by liquid chromatography (LC) with tandem mass spectrometry (MS) (Marchand et al. 2016; Motorin and Marchand 2018; Taoka et al. 2018). 2OMe-seq, Nm-Mut-seq, and NJU-seq can capture relative but not absolute Nm fraction with the use of spiked-in in vitro transcribed RNA controls (Incarnato et al. 2017; Chen et al. 2023; Tang et al. 2024). Low-throughput methods can also measure Nm fraction: RT-based methods such as RTL-P (RT at low dNTP concentrations followed by PCR) provide a semi-quantitative measure of Nm fraction (Dong et al. 2012b; Elliott and Holley 2021), while Nm-VAQ (Nm validation and absolute quantification), which uses site-specific chimeric RNA/DNA probes to detect resistance to RNase H cleavage, is reported to allow absolute quantification of Nm stoichiometry (Tang et al. 2024). The combination of LC and tandem MS (LC–MS/MS) can also be used to quantitatively measure Nm stoichiometry (Motorin and Marchand 2018). Digestion of RNA into single nucleosides or cap dinucleotides allows for the measurement of Nm content by LC–MS/MS (Elliott 2004; Dai et al. 2017; Wang et al. 2019), while analysis of intact or partially digested RNA by LC–MS/MS has enabled the site-specific mapping and quantification of Nm in rRNA and tRNA (Taoka et al. 2016; Wein et al. 2020). LC–MS/MS has the benefit of measuring multiple different types of RNA modifications simultaneously but remains limited by challenges in data analysis and accurate sequence assignments, particularly for complex samples.
High-throughput sequencing-based approaches have successfully been used to map Nm in abundant RNAs such as rRNA, tRNA, and snRNA (Krogh et al. 2017; Marchand et al. 2017; Motorin and Marchand 2018). The detection of Nm in less abundant RNAs is more difficult, in part because many of these methods require large amounts of input RNA (Motorin and Marchand 2018). Nm sites at the 3′ end of miRNAs and piRNAs can be detected by Nm-seq or using a low-throughput method based on the resistance of 3′ Nm to polyadenylation (Dai et al. 2017; Wang et al. 2018). Nm also occurs as part of the 5′ cap of mRNA in higher eukaryotes, with Nm at the first nucleotide (cap-1) occurring at a high stoichiometry across almost all mRNAs, and Nm at the second nucleotide (cap-2) occurring at a lower rate (Galloway and Cowling 2019; Wang et al. 2019; Despic and Jaffrey 2023). Transcriptome-wide 5′ cap 2′-O-methylation can be measured by CLAM-Cap-seq (CircLigase-assisted mapping of caps by sequencing), in which reverse-transcribed complementary DNA (cDNA) is ligated to the 5′ end of the template mRNA, and Nm at the first or second nucleotide is detected by resistance to RNase T2 (Despic and Jaffrey 2023).
In addition to Nm in the 5′ cap, Nm has also been found to occur at internal mRNA sites. When highly purified mRNA is assessed by LC–MS/MS, the Nm/N ratio in mRNA is higher than expected from Nm sites in the 5′ cap alone, supporting the presence of internal Nm sites (Elliott et al. 2019). A few internal Nm sites in mRNA have been identified and validated using low-throughput RT-based methods (Elliott et al. 2019; Leighton et al. 2022), but transcriptome-wide mapping of internal mRNA Nm sites has proved challenging. Although Nm-seq, NJU-seq, Nm-Mut-seq, and MeTH-seq have been used to map Nm in mRNA, only a small subset of the predicted Nm sites have been validated using low-throughput methods, and few of these sites have been validated biochemically (Dai et al. 2017; Bartoli et al. 2018; Chen et al. 2023; Tang et al. 2024). Notably, in the initially published Nm-seq method, a mispriming artifact led to many false-positive Nm sites; although this was corrected in a corrigendum, it further underscores the importance of validation (Dai et al. 2018; Grozhik and Jaffrey 2018). The validity of predicted Nm sites would also be supported if the same Nm sites could be detected using complementary methods, but the overlap between the mRNA Nm sites detected by Nm-seq, Nm-Mut-seq, and NJU-seq has not been examined. Other supportive evidence could include dependence of Nm sites on the cellular machinery that installs Nm (i.e., Nm writers) and overlap of Nm sites with the binding sites of Nm writers or their small nucleolar RNA (snoRNA) guides (Sharma et al. 2016; Gumienny et al. 2017; Dunn-Davies et al. 2021). There is growing evidence that at least some mapped Nm sites are placed by fibrillarin (FBL) (Elliott et al. 2019; Chen et al. 2023; Tang et al. 2024). However, it remains unclear which Nm writers are responsible for most internal Nm in mRNA, and different studies have used different Nm writers as supportive evidence, complicating the interpretation of these results (Dai et al. 2017; Bartoli et al. 2018). Moreover, the binding sites of snoRNAs often lack evidence for Nm modification, suggesting that these binding sites may correspond to noncanonical functions of snoRNAs that are unrelated to Nm, and thus decreasing the utility of this method to provide support for Nm sites in the absence of direct validation (Gumienny et al. 2017; Dunn-Davies et al. 2021). Therefore, although internal Nm sites have been found in mRNA and could potentially have important biological implications, their overall abundance and distribution remain controversial.
REGULATION OF Nm
In general, enzymes that deposit RNA modifications are termed “writers,” while enzymes that remove these modifications are called “erasers.” In eukaryotic rRNA (but not mitochondrial rRNA), Nm modifications are installed cotranscriptionally by box C/D small nucleolar ribonucleoproteins (snoRNPs), which consist of the methyltransferase FBL, the RNA-binding protein 15.5K, a heterodimer of nucleolar protein 56 (NOP56) and NOP58, and a box C/D snoRNA guide that directs the site of 2′-O-methylation based on sequence complementarity (Fig. 2; Monaco et al. 2018). Nm in snRNA is also deposited by box C/D snoRNPs or small Cajal body-specific ribonucleoproteins (scaRNPs), which have the same protein composition as box C/D snoRNPs but use a small Cajal body-specific RNA (scaRNA) guide (Bohnsack and Sloan 2018). However, the writers for some Nm sites in snRNA have not yet been identified (Bohnsack and Sloan 2018). In contrast to most Nm in rRNA and snRNA, most Nm in tRNA is introduced posttranscriptionally by stand-alone enzymes, which in yeast are termed transfer RNA methyltransferase 3 (Trm3), Trm7, Trm13, and Trm44 (Towns and Begley 2012). However, a recent publication has shown that some tRNAs are also subject to snoRNA-guided Nm modification (Zhang et al. 2023). Stand-alone enzymes are also responsible for Nm in the 5′ cap of mRNA and in mitochondrial rRNA (Sirum-Connolly and Mason 1993; Pintard et al. 2002; Byszewska et al. 2015). Box C/D snoRNPs appear to be responsible for guiding at least some internal Nm in mRNA. For example, FBL and two box C/D snoRNAs (U32A and U51) are required for internal Nm in peroxidasin (Pxdn) mRNA, while knockdown of the box C/D snoRNA Snord64 was found to reduce internal Nm in ring finger nuclease 146 (Rnf146) mRNA (Elliott et al. 2019; Leighton et al. 2022). Stand-alone enzymes have also been proposed to function as internal Nm writers in mRNA. In yeast, hundreds of internal Nm sites detected in mRNA by MeTH-seq were found to be dependent on Spb1, the 2′-O-methyltransferase that installs methylguanosine (Gm) at position 2933 in 25S rRNA (Bartoli et al. 2018). Interestingly, the human homolog of Spb1, FtsJ 2′-O-methyltransferase 3 (FTSJ3), is recruited by TAR RNA-binding protein (TRBP) to install internal Nm in human immunodeficiency virus 1 (HIV-1) RNA (Ringeard et al. 2019). Thus, both box C/D snoRNPs and stand-alone enzymes may function as writers of internal Nm in mRNA, but the relative contributions of these and other possible writers remain unclear. Unlike reversible RNA base modifications, Nm does not have known erasers, and given the inert chemistry of 2′-O-methyl, Nm is most likely an irreversible modification.
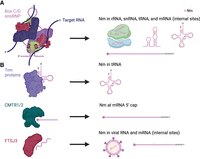
Nm writers. (A) Box C/D snoRNPs deposit Nm on rRNA, snRNA, at some sites in tRNA, and at internal sites in mRNA. (B) Stand-alone enzymes deposit Nm on tRNA, at the 5′ cap of mRNA, and in viral RNA. Stand-alone enzymes have also been proposed to deposit Nm at some internal mRNA sites. Created with BioRender.com.
Despite the lack of known Nm erasers, Nm levels are dynamically regulated. Analysis of Nm at different stages of development by RiboMeth-seq demonstrated that Nm levels in rRNA tend to be higher in adult tissues compared to developing tissues in both zebrafish and mice (Ramachandran et al. 2020; Hebras et al. 2020a). Nm levels also vary by subcellular localization, as in the case of the neuronal small noncoding RNA BC1, which contains Nm in the cell body but is not methylated in the synapse (Lacoux et al. 2012). Additionally, Nm patterns are altered under conditions of stress. Using an RT-based approach, the Nm fraction was found to change in 7 out of 14 tested Nm sites in rRNA upon exposure of human cell lines to hypoxic stress (Metge et al. 2021). Using Nm-seq, mouse macrophages infected with vesicular stomatitis virus were found to have increased Nm in transcripts related to viral infection and increased levels of 2′-O-methyladenosine (Am) in mRNA overall (Li et al. 2022). Nm patterns have also been shown to vary in diseases such as cancer (Dimitrova et al. 2019). Of note, when considering any comparison between samples, it is important to distinguish biologically significant variability from the technical variability associated with the measurement method being used, which is particularly relevant for Nm given the limitations of current Nm mapping methods (Grozhik and Jaffrey 2018). Moreover, biological changes in Nm levels should be validated using low-throughput or ideally biochemical approaches, particularly in less abundant RNAs such as mRNA, in which the accuracy of high-throughput Nm mapping remains unclear.
The plasticity of Nm in different physiologic and pathologic states suggests the existence of regulatory mechanisms that modulate Nm levels. In particular, the variability of Nm in rRNA contributes to ribosome heterogeneity, and the modulation of Nm levels in rRNA can be used to regulate populations of differentially methylated ribosomes under different conditions (Monaco et al. 2018). Since Nm erasers do not seem to exist, Nm levels are likely determined primarily by RNA turnover and at the level of Nm writers or snoRNA guides. The manipulation of Nm writers is made challenging by the lethality of FBL knockout, the existence of hundreds of distinct snoRNA guides, and the absence of a one-to-one relationship between Nm sites and Nm writers/guides, in which multiple different writers/guides can methylate a single Nm site or a single writer/guide can methylate multiple different Nm sites (Newton et al. 2003; Bergeron et al. 2020). FBL knockdown has differential effects on the methylation of different Nm sites in rRNA: Nm sites at the functional core of the ribosome tend to be relatively insensitive to FBL knockdown, perhaps because these sites are more efficiently methylated or because ribosomes lacking these Nm modifications do not pass quality control, whereas peripheral Nm sites are more sensitive to FBL knockdown (Erales et al. 2017; Sharma et al. 2017; Jaafar et al. 2021). Regulation of FBL levels may play a role in tumorigenesis. Notably, the proto-oncogene MYC up-regulates FBL expression, whereas the tumor suppressor p53 directly represses transcription of the FBL gene, with downstream effects on translation and cell proliferation (Marcel et al. 2013; Jansson et al. 2021). Regulation of writers also occurs during development. Adult mouse tissues exhibit lower levels of the box C/D snoRNA Snord78, as well as decreased methylation of its target site Gm4593 in 28S rRNA (Hebras et al. 2020a). The down-regulation of Snord78 during development has been proposed to occur via differential alternative splicing of its host gene, growth arrest-specific 5 (GAS5), which leads to less efficient processing of Snord78 from introns in the adult mouse (Hebras et al. 2020a).
Regulation of Nm can also occur at the level of writer-associated factors. Independent of its functions in transcriptional repression, enhancer of zeste homolog 2 (EZH2) has been shown to bind FBL and NOP56 to facilitate box C/D snoRNP assembly, and EZH2 depletion leads to decreased methylation of a subset of rRNA Nm sites (Yi et al. 2021). The impact of EZH2 on Nm in rRNA may be one of the mechanisms by which EZH2 drives tumorigenesis in certain tumor types such as prostate cancer (Yi et al. 2021). Two other factors that modulate rRNA Nm levels through interactions with box C/D snoRNPs are nucleophosmin 1 (NPM1) and groucho-related amino-terminal enhancer of split (AES), both of which are implicated in the pathogenesis of acute myeloid leukemia (AML) (Zhou et al. 2017; Nachmani et al. 2019). However, in many cases it remains unclear why modulation of these factors leads to changes in Nm levels at only a subset of rRNA Nm sites. Other writer-associated factors have been found to impact Nm levels in snRNA. TAR DNA-binding protein 43 (TDP43), which is involved in the pathogenesis of amyotrophic lateral sclerosis, binds to UG-rich box C/D scaRNAs and promotes their localization to Cajal bodies, and depletion of TDP43 reduces Nm at the target sites of these scaRNAs in U1 and U2 snRNAs (Izumikawa et al. 2019). La-related protein 7 (LARP7) has been shown to facilitate the interaction between box C/D snoRNAs and U6 snRNA, and depletion of LARP7 leads to a reduction in Nm levels in U6 snRNA (Hasler et al. 2020; Wang et al. 2020). The loss of Nm in U6 snRNA due to LARP7 depletion has consequences for spermatogenesis in mice and may contribute to the phenotype of the developmental disorder Alazami syndrome in humans (Hasler et al. 2020; Wang et al. 2020).
FUNCTIONAL IMPLICATIONS OF Nm
Unlike base modifications, Nm occurs on the nucleotide ribose moiety and thus does not directly affect base-pairing (Fig. 3A). However, the 2′-O-methyl group stabilizes the C3′-endo ribose conformation found in the A-form RNA helix or in stacked nucleotides, and as a result, each Nm stabilizes RNA duplexes by ∼0.2 kcal/mol (Yildirim et al. 2014). Using TAR RNA as a model system, Nm was shown to preferentially stabilize and prolong the lifetime of conformations in which Nm is in a paired helical conformation, demonstrating that one function of Nm may be to modulate RNA secondary structure (Abou Assi et al. 2020). The conformational biases of Nm may explain how Nm in the tRNA anticodon loop influences codon recognition, and how Nm sites in rRNA stabilize distorted RNA conformations at functional interfaces of the ribosome (Kawai et al. 1992; Polikanov et al. 2015). However, for most Nm sites, the structural consequences of Nm have not been directly examined. Using single-molecule Förster resonance energy transfer, one study demonstrated that several Gm sites in U2 snRNA have an additive stabilizing effect on U2 stem I, and thereby alter the dynamic equilibrium of the U2–U6 complex (Karunatilaka and Rueda 2014). However, the energetic effect of Nm modification was smaller than expected given the observed impact of Nm on spliceosome assembly in vivo, raising the possibility that Nm on U2 snRNA in vivo might primarily function through other mechanisms such as by promoting protein interactions (Dönmez et al. 2004; Karunatilaka and Rueda 2014).
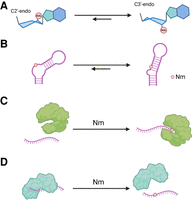
Effects of Nm on RNA structure and protein binding. Nm stabilizes the C3′-endo conformation of the ribose moiety of nucleotides (A) and stabilizes RNA conformations in which Nm is in an RNA duplex (B). Proteins may selectively bind RNA with Nm (C) or may have decreased affinity for RNA with Nm (D). Created with BioRender.com.
In addition to (or as a result of) its effects on RNA structure, Nm may also influence protein binding, which is a common mechanism by which other RNA modifications exert their cellular functions (Fig. 3B; Yang et al. 2018; Guo et al. 2021). RNA-binding proteins that preferentially bind to RNAs with a specific RNA modification are referred to as “readers” of that RNA modification. Piwi family proteins and Argonaute 2 (AGO2) could be considered to be readers of Nm, since they demonstrate a weak preference for binding piRNAs and miRNAs with 3′ Nm (Tian et al. 2011; Liang et al. 2020). Surprisingly, there have not been reports of proteins that bind to Nm with high specificity. However, Nm modification of RNA decreases the binding affinity of a number of proteins. For example, AGO1 binding to RNA is reduced in the presence of 3′ Nm, likely due to steric hindrance (Tian et al. 2011). In addition, an internal Nm site in the pri-miRNA pri-let7a has been proposed to decrease binding of the microprocessor complex subunit DiGeorge syndrome critical region 8 (DGCR8), leading to decreased let-7a maturation, although the impact of Nm on DGCR8 binding has not been directly demonstrated (Bian et al. 2023). Most notably, the immune receptors retinoic acid inducible gene I (RIG-I) and interferon-induced protein with tetratricopeptide repeats (IFIT1) have decreased binding affinity for mRNA with Nm at the 5′-most nucleotide position, as in the cap-1 form of the mRNA 5′ cap (Hyde and Diamond 2015; Devarkar et al. 2016). The addition of Nm at the second nucleotide (cap-2) further decreases RIG-I binding (Despic and Jaffrey 2023). In this way, Nm at the 5′ cap of mRNA functions to prevent the activation of the innate immune response by self-RNAs. Cap-1 Nm also inhibits the binding of decapping exoribonuclease (DXO), which selectively degrades RNAs lacking Nm at the first nucleotide as part of RNA quality control before nuclear export (Picard-Jean et al. 2018).
Although the molecular mechanisms are not always clear, Nm modifications have been shown to influence diverse cellular processes including translation, splicing, and innate immunity. FBL knockdown alters Nm levels at a subset of rRNA sites, producing ribosomes with lower efficiency of translation from several internal ribosome entry sites (IRES) but equally efficient cap-dependent translation (Erales et al. 2017). In another example, which suggests a role for rRNA Nm in regulating IRES-dependent translation, the vascular endothelial growth factor C (VEGF-C) IRES was shown to associate with ribosomes with a distinct Nm pattern under hypoxic conditions (Metge et al. 2021). Altering rRNA Nm levels can also impact other aspects of translation. In yeast, mutation of the box C/D snoRNP assembly factor Bcd1 (box C/D snoRNA protein 1) resulted in decreased Nm levels at a subset of rRNA sites, as well as decreased translational fidelity, changes in ribosome rotational status, and decreased ribosome binding to eukaryotic initiation factor 1 (Khoshnevis et al. 2022). In human cell lines, knockdown of the box C/D snoRNA Snord45C led to loss of 2′-O-methylation at C174 in 18S rRNA, which in turn led to decreased efficiency of translation of mRNA transcripts containing AU-rich codons (Jansson et al. 2021). Thus, Nm in rRNA contributes to ribosome heterogeneity, and alteration of Nm patterns can up- or down-regulate certain ribosome populations to modulate the translation of certain pools of mRNA transcripts (Monaco et al. 2018). In addition to Nm in rRNA, Nm in tRNA can also impact translation efficiency of specific codons. For example, knockout of the Trm7 homolog FTSJ1, which introduces Nm in the anticodon loop of tRNAPhe(GAA), results in decreased translation efficiency of UUU but not UUC codons (Li et al. 2020). Additionally, Nm in the mRNA 5′ cap increases translation efficiency in at least some systems (Kuge et al. 1998; Zamudio et al. 2009; Sikorski et al. 2020). In contrast, Nm in coding regions of mRNA tends to decrease translation efficiency, especially at the second nucleotide position of a codon, although the magnitude of this effect depends on the codon context (Hoernes et al. 2016, 2019). A crystal structure of the Escherichia coli ribosome provides a mechanism for this internal mRNA Nm-induced decrease in translation efficiency: Nm at the second nucleotide position of a codon disrupts the interaction of ribosomal-monitoring bases with the codon–anticodon helix, leading to the inappropriate rejection of cognate tRNAs (Choi et al. 2018). Although the impact of internal Nm on mRNA translation was initially demonstrated only in synthetically modified RNAs, the snoRNA-guided internal Nm modification of Pxdn mRNA was found to inhibit translation, with either knockout of the corresponding guide snoRNAs or point mutation of the Nm site in Pxdn mRNA leading to increased translation as measured by metabolic pulse-labeling (Elliott et al. 2019).
In addition to its multiple effects on translation, Nm has been implicated in RNA splicing and turnover. Nm sites in snRNAs are clustered in functionally important sequences that are involved in RNA–RNA or RNA–protein interactions, suggesting that Nm might fine-tune these interactions, although studies directly examining this putative function are limited (Bohnsack and Sloan 2018). Nm at positions 1, 2, 12, and 19 in U2 snRNA are individually required for the formation of the ATP-independent early spliceosomal E complex and for splicing in HeLa cells, possibly by playing a role in the interaction of U2 with U1 snRNA or other factors (Dönmez et al. 2004). Loss of LARP7, which facilitates the Nm modification of U6 snRNA, was found to induce significant changes in mRNA splicing (Wang et al. 2020). These splicing changes were rescued by wild-type LARP7, but not by a mutant form of LARP7 specifically defective in U6 snRNA loading, suggesting that modulation of Nm in U6 snRNA is a potential mechanism for regulating alternative splicing (Wang et al. 2020). However, another study found only a minor impact of LARP7 knockout on splicing under normal conditions, whereas more splicing changes were observed at high temperature, suggesting that LARP7 may be important for maintaining splicing robustness under stress (Hasler et al. 2020). Nm in mRNA could in principle impact splicing as well. Transcriptome-wide mapping of Nm suggests that Nm sites are enriched around mRNA splice sites (Dai et al. 2017; Tang et al. 2024). The artificial introduction of Nm at the branchpoint adenosine in pre-mRNA leads to the use of cryptic branchpoints and can prevent splicing in the absence of cryptic branchpoints (Ge et al. 2010). To date, no naturally occurring Nm sites in pre-mRNA have been shown to specifically modulate alternative splicing, although the box C/D snoRNA Snord115 regulates the alternative splicing of the serotonin receptor 2c (HTR2C) mRNA, and HTR2C mRNA contains an internal Nm site (Hebras et al. 2020b; Leighton et al. 2022). However, it remains unclear whether Snord115 guides this Nm modification and whether Nm modification leads to changes in HTR2C mRNA splicing. In terms of RNA stability, Nm at both the first and second nucleotide positions (in cap-1 and cap-2) confer resistance to decapping and degradation by DXO (Picard-Jean et al. 2018; Drazkowska et al. 2022). In addition, 3′ Nm increases the stability of miRNA, piRNA, and small interfering RNA (siRNA) by blocking 3′-to-5′ exoribonuclease activity, as well as 3′ uridylation in plant small RNAs, animal piRNAs, and fly AGO2-associated siRNAs (Ji and Chen 2012; Liang et al. 2020). Finally, internal Nm modification may increase the stability of Pxdn mRNA based on the decrease in Pxdn mRNA expression upon FBL knockdown, although the mechanism by which internal Nm might promote mRNA stability is unknown (Elliott et al. 2019). Direct protection from RNA cleavage seems unlikely, but recruitment of Nm-binding proteins could protect mRNA from degradation or sequester mRNA in a way that prolongs its half-life.
RNA modifications play a critical role in allowing the innate immune system to distinguish self from non-self RNAs. In addition to internal base modifications, which suppress the activation of the innate immune response through Toll-like receptors (TLRs), 5′ cap Nm suppresses the activation of the innate immune response through RIG-I and IFIT1 (Karikó et al. 2005; Hyde and Diamond 2015; Schuberth-Wagner et al. 2015; Despic and Jaffrey 2023). Many RNA viruses encode cap-1 2′-O-methyltransferases that place Nm at the 5′ end of viral RNA to impair recognition by RIG-I and IFIT1 (Hyde and Diamond 2015; Schuberth-Wagner et al. 2015). Other viruses, such as orthomyxoviruses, co-opt the host 5′ cap by using the 5′ end of host mRNA as a primer for viral transcripts, a process termed cap snatching (Hyde and Diamond 2015). Interestingly, interferon-β (IFN-β) stimulates expression of host cap-specific mRNA nucleoside-2′-O-methyltransferase 1 (CMTR1), which deposits Nm on the first nucleotide of host IFN-stimulated gene (ISG) transcripts, thereby marking ISG transcripts as self-RNAs and ensuring their translation as part of the antiviral response (Williams et al. 2020). In addition to 5′ cap Nm, internal Nm may play a role in preventing activation of the innate immune response. The methyltransferase FTSJ3 deposits internal Nm in HIV-1 RNA, and viral particles produced in cells treated with FTSJ3 siRNA trigger type 1 IFN expression (Ringeard et al. 2019). Interestingly, this type 1 IFN expression is independent of RIG-I and instead depends on the RNA sensor melanoma differentiation-associated protein 5 (Ringeard et al. 2019). The cap 2′-O-methyltransferases of flaviviruses have also been found to introduce internal Am sites in viral RNA, although the function of these Am sites in immune evasion is unclear (Dong et al. 2012a). In addition to Nm in mRNA, Gm18, which is found in the tRNAs of certain Gram-negative bacteria, has been shown to suppress the TLR7-mediated immune response in human peripheral blood mononuclear cells (PBMCs) (Gehrig et al. 2012; Jöckel et al. 2012). In macrophages, the NPM1 D180del mutation, which decreases Nm levels in rRNA, leads to reduced phagocytosis and lower levels of interleukin-β (IL-β) and IL-6 (Sportoletti et al. 2020). Of note, FBL expression decreases upon infection, and FBL knockdown increases resistance to viral and bacterial infection, although it is unclear whether these effects are related to 2′-O-methylation of rRNA or other targets, or are related to methylation-independent functions of FBL (Tiku et al. 2018; Li et al. 2022).
DISEASE RELEVANCE OF Nm
Nm is dysregulated in a variety of diseases (Table 2), but examples where alterations in Nm modification underlie or cause disease are limited (Dimitrova et al. 2019). Using RiboMeth-seq, Nm patterns in rRNA were shown to differ in breast cancer tissues compared to normal breast tissues, as well as among different breast cancer subtypes (Marcel et al. 2020). In diffuse large B-cell lymphoma (DLBCL), Nm levels in rRNA were more significantly decreased in activated B-cell-type DLBCL, which is the more aggressive subtype, than in germinal center B-cell-type DLBCL (Krogh et al. 2020). Interestingly, Nm levels in rRNA tend to be decreased in cancer, which could indicate a decoupling between ribosome biogenesis and the Nm writers, in which the rate of methylation cannot keep pace with the increased rate of ribosome biogenesis in cancer cells (Jaafar et al. 2021). However, Nm modification increases at a subset of rRNA sites in cancer, supporting the existence of site-specific regulation of Nm levels (Jaafar et al. 2021). Nm levels in other classes of RNA may also be dysregulated in cancer. For instance, Jurkat cells, which are a T-cell leukemia model, exhibit changes in the stoichiometry of Nm at certain sites in snRNA (Krogh et al. 2017). Another study found that the miRNA, miR-21-5p, had increased 3′ Nm levels in non-small cell lung cancer (NSCLC) compared to normal lung tissue (Liang et al. 2020). This differential methylation was proposed to lead to greater stability of miR-21-5p in NSCLC, leading to increased inhibition of translation of the tumor suppressor programmed cell death protein 4 (Liang et al. 2020). However, studies on the distribution of Nm in cancer have primarily examined Nm in rRNA, in part because the analysis of Nm in patient samples is limited by the high input RNA requirements of many currently available high-throughput Nm mapping methods.
Nm in disease
Supporting a role for Nm in cancer, FBL and snoRNAs are overexpressed across multiple tumor types (Su et al. 2014; Gong et al. 2017; Barros-Silva et al. 2021; Kärkkäinen et al. 2022). Moreover, knockdown of box C/D snoRNP components including FBL, NOP56, and NOP58 decreases the tumorigenicity of breast cancer cell lines (Su et al. 2014). Further supporting an essential role for box C/D snoRNPs in tumorigenesis, Nm writers/guides are regulated by multiple oncogenic signaling pathways. The proto-oncogene MYC, as well as the oncogenic fusions AML–ETO9a and MLL–AF9, induce box C/D snoRNA expression in AML, while MYC up-regulates and TP53 down-regulates FBL expression in solid tumors (Marcel et al. 2013; Zhou et al. 2017; Jansson et al. 2021). Nm writers have also been linked to prognosis in patients with cancer. Expression of FBL is associated with poor prognosis in multiple tumor types, including breast cancer and melanoma (Su et al. 2014; Jasinski-Bergner et al. 2021). Certain snoRNAs have been found to associate with survival in solid tumors, and box C/D snoRNA levels in AML correlate with the frequency of leukemic stem cells (Gong et al. 2017; Zhou et al. 2017). The up-regulation of Nm writers in cancer may simply reflect the need for more efficient methylation of rRNA to support increased ribosome biogenesis in the setting of rapid proliferation. However, Nm writers may also contribute to tumorigenesis by modulating translation. Both FBL and the box C/D snoRNA Snord42A promote the IRES-dependent translation of proteins including MYC, VEGF-A, and ribosomal proteins (Marcel et al. 2013; Zhou et al. 2017). Other Nm writers may promote tumorigenesis through targets other than rRNA. For example, the box C/D snoRNA Snord11B deposits Nm on pri-let-7a, leading to decreased expression of the mature miRNA let-7a-5p and thereby inhibiting apoptosis (Bian et al. 2023). Interestingly, in melanoma, the expression of box C/D snoRNP components negatively correlates with the expression of genes involved in antigen processing and presentation, raising the possibility that Nm writers may play a role in tumor immune invasion (Jasinski-Bergner et al. 2021).
Besides cancer, Nm has been implicated in other disease states, including cardiovascular and neurologic disorders. SnoRNAs seem to play an important role in cardiac development. Morpholino-mediated knockdown of scaRNA1 and Snord94 in zebrafish leads to changes in the splicing of cardiac regulatory genes and disrupts cardiac development (Patil et al. 2015). Moreover, the right ventricular myocardium of infants with the congenital heart disorder Tetralogy of Fallot demonstrates differential expression of snoRNAs that guide the Nm modification of U2 and U6 snRNAs, as well as changes in alternative splicing patterns (O'Brien et al. 2012; Patil et al. 2015; Nagasawa et al. 2018). SnoRNAs encoded by the ribosomal protein L13a (Rpl13a) locus, which guide Nm modifications in the 28S and 18S rRNAs, have been implicated in systemic glucose metabolism, the response to lipid-induced oxidative stress, and the development of atherosclerosis (Michel et al. 2011; Holley et al. 2015; Lee et al. 2016; Zhang et al. 2022; Sun et al. 2023). SnoRNAs may also play a role in the pathogenesis of certain neurodevelopmental disorders. Prader–Willi syndrome (PWS) is caused by paternal loss of imprinted genes in locus 15q11–q13, which includes many copies of Snord115 and Snord116. Snord115 has been proposed to contribute to PWS pathogenesis by altering adenosine-to-inosine editing and alternative splicing of HTR2C mRNA (Hebras et al. 2020b). Another study suggested that HTR2C mRNA contains internal Nm based on an RT-based assay, although the writer for this putative Nm was not identified (Leighton et al. 2022). However, the importance of Snord115 in the pathogenesis PWS has since been questioned, since Snord115-deficient mice do not exhibit the behavioral changes characteristic of PWS (Hebras et al. 2020b). Snord116 is likely critical in the pathogenesis of PWS given that microdeletion of the Snord116 cluster has been observed in a patient with PWS (Duker et al. 2010). Although multiple mRNA targets for Snord116 have been proposed based on sequence complementarity and evolutionary conservation, these putative targets have not been experimentally validated, and the mechanism by which Snord116 may contribute to PWS remains unclear (Baldini et al. 2022). Another neurodevelopmental disorder, non-syndrome X-linked intellectual disability (NSXLID), can be caused by mutations in FTSJ1, the 2′-O-methyltransferase that deposits Nm in the anticodon loop of certain tRNAs (Dimitrova et al. 2019). Certain patients with NSXLID have a missense mutation in FTSJ1 that specifically leads to loss of Nm34 in target tRNAs, suggesting a specific role for Nm34 in the pathogenesis of NSXLID (Guy et al. 2015). Mechanisms by which Nm34 has been proposed to lead to NSXLID include the modulation of translation, the activation of the general amino acid control response, and the accumulation of tRNA fragments (Dimitrova et al. 2019). In addition to their functions in neurodevelopment, box C/D snoRNAs have been suggested to play a role in fear extinction learning (Leighton et al. 2022). Extinction learning led to increased Snord64 levels in the synapse, and Snord64 knockdown enhanced the retention of fear-related memory (Leighton et al. 2022).
In summary, Nm and Nm writers have been implicated in multiple human diseases. However, our knowledge about the regulation of Nm in disease remains limited since most studies have only examined Nm in rRNA rather than in other RNA classes. Moreover, reliable data on the expression of snoRNAs in disease is largely lacking since conventional RNA sequencing approaches exclude small RNAs such as snoRNAs. Even for box C/D snoRNAs that are implicated in disease, their target RNAs may be unknown, they may methylate multiple targets, or it may be unclear whether the snoRNAs contribute to disease through their canonical function of depositing Nm modifications or through other Nm-unrelated mechanisms. Particularly for multigenic diseases, the impact of Nm or Nm writers on disease may only be evident in the appropriate genetic context. Finally, given the plasticity of Nm, processes such as development and cancer metastasis might be dependent on the dynamics of Nm and Nm writers, rather than their levels at a specific snapshot in time. These are some of the critical gaps in knowledge that need to be addressed.
CONCLUSION
Within the last decade, the field of Nm has seen considerable progress, including the development of high-throughput methods for mapping Nm, the discovery of internal Nm sites in mRNA, and insights into the impact of Nm on physiologic and pathologic processes. However, challenges remain in understanding the distribution, regulation, and function of Nm. While high-throughput Nm mapping methods have greatly enhanced our understanding of the variability of Nm in rRNA under different physiologic conditions and disease states, the reliability of Nm mapping for less abundant RNAs remains unclear. To establish the reproducibility and accuracy of Nm mapping more clearly in mRNA, orthogonal Nm mapping methods should be shown to consistently identify the same Nm sites, and putative Nm sites identified using these methods need to be validated. While the plasticity of Nm has been well documented, in most cases the mechanisms underlying this observed plasticity remain unknown. Moreover, while Nm has been shown to influence multiple cellular processes, the functions of most Nm sites are still unclear. The use of genetic approaches to modulate Nm writers or snoRNA guides may help clarify how Nm levels are regulated, as well as identify the function of specific Nm sites. The development of programmable box C/D snoRNAs that introduce Nm at a designated target site would be another useful tool to help elucidate the function of Nm in a site-specific manner (Deryusheva and Gall 2019). Finally, the application of these approaches to patient samples and disease models has the potential to provide valuable information about the function of Nm in human disease.
ACKNOWLEDGMENTS
K.I.Z. was supported by the Duke Hematology and Transfusion Medicine Training Program (T32 HL007057). C.V.P. was supported in part by the National Institutes of Health (NIH) R01CA215075, R01CA258451, and 1R41CA246848, the Lung Cancer Research Foundation, the Free to Breathe Metastasis Research Award, and a North Carolina Biotechnology Translation Research Grant (NCBC TRG). C.L.H. was supported by NIH R01 GM135383 and R01 HL146381.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079970.124.
-
Freely available online through the RNA Open Access option.
This article, published in RNA, is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








