
Independent neofunctionalization of Dxo1 in Saccharomyces and Candida led to 25S rRNA processing function
- 1Department of Microbiology and Molecular Genetics, University of Texas Health Science Center at Houston, Houston, Texas 77030, USA
- 2UT MD Anderson Cancer Center UTHealth Houston Graduate School of Biomedical Sciences, University of Texas Health Science Center at Houston, Houston, Texas 77030, USA
- Corresponding author: ambro.van.hoof{at}uth.tmc.edu
-
Handling editor: Eric Phizicky
Abstract
Eukaryotic genomes typically encode one member of the DXO/Dxo1/Rai1 family of enzymes, which can hydrolyze the 5′ ends of RNAs with a variety of structures that deviate from the canonical 7mGpppN. In contrast, the Saccharomyces genome encodes two family members and the second copy, Dxo1, is a distributive 5′ exoribonuclease that is required for the final maturation of the 5′ end of 25S rRNA from a 25S′ precursor. Here we show that this 25S rRNA maturation function is not conserved across kingdoms, but arose in the budding yeasts. Interestingly, the origin of 25S processing capacity coincides with the duplication of this gene, and this capacity is absent in the nonduplicated genes. Strikingly, two different clades of budding yeasts have undergone parallel evolution: Both duplicated their DXO/Dxo1/Rai1 gene, and in both cases, one copy gained the 25S processing function. This was accompanied by many parallel sequence changes, a remarkable case of reproducible neofunctionalization.
Keywords
INTRODUCTION
Noncoding RNAs are transcribed by one of the three RNA polymerases and then extensively processed by RNases. A prime example of this is the ribosomal RNA (rRNA) processing pathway, which is well-characterized in both Saccharomyces cerevisiae (yeast) and human and appears highly conserved across eukaryotes (for recent reviews, see Kalisiak et al. 2017; Dorner et al. 2023; Schneider and Bohnsack 2023). In yeast, at least 13 RNases are required to process the initial RNA polymerase I transcript into the mature 18S, 5.8S, and 25S rRNAs, and many of these steps and enzymes are conserved between yeast and human. The RNases involved are classified as 5′ to 3′ exoribonucleases, endoribonucleases, and 3′ to 5′ exoribonucleases. Endoribonucleases separate the initial transcript into five segments by cutting in the external and internal transcribed spacers (5′ ETS, 3′ETS, ITS1, and ITS2) (Schmitt and Clayton 1993; Kufel et al. 1999; Castle et al. 2010,2013; Wells et al. 2017; Lan et al. 2020). The processing of three of these segments into 18S, 5.8S, and 25/28S rRNA is carried out by additional RNases. The 18S rRNA is generated by additional endonucleases that cut at the mature 5′ and 3′ ends (Utp24/FCF1 and Nob1/NOB1, respectively, in yeast/human [Fatica et al. 2004; Bleichert et al. 2006]). In contrast, the 5.8S ends are generated by exonucleases. The 3′ end of 5.8S rRNA is generated by sequential action of several 3′ exonucleases: Rrp44, Rrp6, Rex1, Rex2, Rex3, and Ngl2 in yeast and RRP44 (aka DIS3), RRP6 (aka EXOSC10), ISG20L2, ERI1, and probably others in human (Mitchell et al. 1996; Briggs et al. 1998; van Hoof et al. 2000; Faber et al. 2002; Ansel et al. 2008; Coute et al. 2008; An et al. 2024). These 3′ exonucleases sequentially trim the 3′ end, with some overlapping function between them. In contrast to several RNases contributing to 3′ end formation, a single 5′ exonuclease (Rat1/XRN2) is responsible for the formation of the 5′ end of 5.8S rRNA in both yeast and human. While all of these steps are conserved, the ends of the largest rRNA, 25S in yeast and 28S in human, are generated by species-specific mechanisms. In yeast, the 3′ETS is cleaved by the endoribonuclease Rnt1 (Kufel et al. 1999), which is absent from the human genome, followed by trimming of the 25S 3′ end by Rex1 and Rex4 (Kempers-Veenstra et al. 1986; An et al. 2024). In contrast, it is not clear how the human 28S 3′ end is produced. The endonuclease that initially cleaves the human 3′ ETS remains to be identified. In fruit flies, this cleavage is followed by trimming by the exonuclease REXO5 (Gerstberger et al. 2017). Given that fruit flies and yeast both require a Rex/REXO family member for 25/28S 3′ end maturation, it seems likely that one or more human REXO family members perform this function. Whether the 5′ end is produced by a conserved mechanism is not clear: Initially, it was thought that the 5′ end was generated by Rat1/XRN2 in both yeast and human (Kalisiak et al. 2017; Dorner et al. 2023; Schneider and Bohnsack 2023). However, we recently identified Dxo1 as a previously uncharacterized distributive 5′ exoribonuclease that is required for the final 5′ end maturation of yeast 25S rRNA from a 25S′ intermediate (Hurtig and van Hoof 2022). A similar 28S′ intermediate has been described in mammals, but how its 5′ end is processed to the final 28S is unclear (Mullineux and Lafontaine 2012). These complex rRNA processing steps appear to be necessitated to allow coordination with the assembly of ribosomal subunits (e.g., Vanden Broeck and Klinge 2023).
Dxo1 is a member of the DXO/Dxo1/Rai1 family of enzymes that have a variety of decapping and 5′ exonuclease functions (Chang et al. 2012). Family members cleave the 5′ ends of a variety of RNAs and produce 5′ monophosphorylated products. The substrates can include triphosphate ends, unmethylated GpppN caps, NAD and FAD caps, and 5′ hydroxylated RNAs. A typical eukaryotic genome encodes one family member, and this is the case for human (the DXO or DOM3Z gene), the model plant Arabidopsis (DXO1), and the fission yeast Schizosaccharomyces pombe (rai1+ aka din1+). However, some genomes encode more than one DXO/Dxo1/Rai1 family member. The budding yeast and fruit fly genome each encode two family members, Rai1 and Dxo1 in budding yeast (Chang et al. 2012) and Rai1 and Cutoff in fruit flies (Chen et al. 2016). Even more extreme, the Caenorhabditis elegans genome encodes ten family members. How these duplicated genes differ from each other is poorly characterized. Yeast Dxo1 is a distributive 5′ exoribonuclease that is localized in the cytoplasm and required for the final maturation of 25S rRNA (Hurtig and van Hoof 2022). In contrast, yeast Rai1 is nuclear and has a more pronounced decapping activity that is thought to limit the accumulation of mRNAs with aberrant caps (Wang et al. 2015). In addition, the namesake function of Rai1 is as a noncatalytic Rat1 interacting protein that stabilizes the Rat1 protein in yeast (Xue et al. 2000). In fruit flies, both Cutoff and Rai1 interact with Rat1, but only Rai1 is thought to have enzymatic activity while Cutoff has a nonenzymatic function in piRNA expression (Chen et al. 2016; Pritykin et al. 2017). The protein sequence and structure changes that cause this diversification of function of the DXO/Dxo1/Rai1 family are incompletely understood.
In a previous publication, we used a transcriptome-wide approach to identify products of Dxo1 by sequencing 5′ monophosphorylated RNA ends (Hurtig and van Hoof 2022). We detected that Dxo1 can remove up to a few nucleotides from decapped or cleaved mRNAs when Xrn1 is absent, thus acting as a distributive 5′ exoribonuclease. However, the major function we identified is processing the 25S 5′ end. In a dxo1Δ strain, essentially no fully mature 25S rRNA was present, but instead, its precursor 25S′ persisted. The majority of 25S′ has a 2 or 7 nt extension on its 5′ end.
As mentioned above, in yeast, Dxo1 and Rai1 are duplicated genes of the DXO/Dxo1/Rai1 family. The related yeast Kluyveromyces lactis shares this duplication, and biochemical characterization of its Dxo1 suggests that a few amino acid changes changed the activity from decapping to distributive 5′ exonuclease activity (Chang et al. 2012). This suggests several possible models. First, the ancestral single ortholog carried out the 25S′ processing function as well as Rai1's function. After duplication, these two tasks were divided between the two genes, which would be an example of subfunctionalization (Hughes 1994; Force et al. 1999). Alternatively, the ancestral single ortholog was unable to carry out 25S′ to 25S processing, and this function arose in the DXO1 gene after duplication, which would be an example of neofunctionalization (Ohno 1970). Perhaps simultaneously optimizing the catalytic activities for Rai1 and Dxo1 function in one protein is impossible, and gene duplication allowed for escape from this adaptive conflict.
In this manuscript, we set out to determine whether the role of Dxo1 in final 25S′ to 25S rRNA maturation was conserved across eukaryotes and thus ancestral to the Dxo1/Rai1 duplication. We show that the single homologs in humans, Arabidopsis and S. pombe are not required for this processing step and speculate that some other distributive 5′ exonuclease carries out this role. In contrast, species that are closely related to yeast and have two DXO/Dxo1/Rai1 family members have one family member that is functionally interchangeable with Dxo1 and one that is functionally interchangeable with Rai1. Overall, our data suggest that the duplication of this gene allowed Dxo1 to evolve a new function in 25S rRNA maturation.
RESULTS
DXO/Dxo1/Rai1 family members are not required for final 5′ end maturation of rRNA in humans, plants, or fission yeast
First, we examined whether the rRNA processing function of the DXO/Dxo1/Rai1 family is conserved across kingdoms, focusing on humans and Arabidopsis as representative of the animal and plant kingdoms with a characterized DXO/Dxo1/Rai1. Humans and Arabidopsis each have only one family member, HsDXO and AtDXO1, respectively. To investigate if HsDXO processes rRNA like S. cerevisiae Dxo1 (ScDxo1), we obtained DXO knockout HEK293T cells (generously provided by Mike Kiledjian, Rutgers University [Jiao et al. 2017]). These cells grew slightly slower, and western blot confirmed that they lacked HsDXO. To detect a possible 28S rRNA processing defect, we isolated RNA from dxo and control HEK293T and analyzed it using the same RNA and bioinformatic methods, we previously used to detect a 25S processing defect in S. cerevisiae dxo1Δ (Fig. 1A–C). Briefly, a linker is ligated onto the 5′ ends of RNAs and used to build and sequence a next-generation sequencing library. The first nucleotide of the read therefore precisely maps the 5′ end of the RNA and is plotted. The results showed that the mutant and control cells generated the identical 5′ end of 28S rRNA, indicating that HsDXO is not required for 28S rRNA processing (Fig. 1C). These data also map the 5′ ends of 18S and 5.8S rRNA, and these were also very similar in both samples (Fig. 1A,B). For comparison, we have included our previously published S. cerevisiae data in Figure 1J, which clearly shows the difference in 25S rRNA. For the HEK cell data, we did observe some differences across the transcriptome between the dxo and control cells, but further analysis will be required to gain a deeper understanding of the human degradome and novel targets or functions of HsDXO. These data indicate that the human DXO is not required for rRNA 5′ end maturation.

The requirement for DXO/Dxo1/Rai1 for 25S′ processing is not conserved across kingdoms. Shown is next-generation sequence data for RNAs with a 5′ monophosphate mapped to the rDNA repeat. This analysis defines the 5′ end of the 18S small subunit RNA (ssRNA) (A, D, and G), 5.8S RNA (B, E, and H) and 25S/28S large subunit rRNA (lsRNA) (C, F, and I), which are unaffected by a null allele of the sole DXO/Dxo1/Rai1 family member in H. sapiens (A–C), A. thaliana (D–F), and S. pombe (G–I). In contrast, our previous published data (J) show a specific defect in 25S rRNA processing in S. cerevisiae (Hurtig and van Hoof 2022). This panel is in gray scale instead of color to distinguish it as previously published.
Next, we analyzed the role of the single Arabidopsis DXO/Dxo1/Rai1 homolog in 25S 5′ end maturation. A recent study (Zakrzewska-Placzek et al. 2022) used northern blot analysis to show that the Atdxo1-2 mutant accumulated slightly increased levels of rRNA processing intermediates, but the exact processing step that AtDXO1 affects was not identified. Furthermore, the accumulation of these intermediates was reversed by expressing a catalytically inactive form of AtDXO1, and thus these phenotypes do not reflect a catalytic function of AtDXO1 in Arabidopsis rRNA maturation. To study whether AtDXO1 is required for 25S rRNA 5′ end maturation, we analyzed publicly available RNA 5′ end sequencing data from wild-type (col-0) and dxo1-2 mutant Arabidopsis (Yu et al. 2021). We mapped and analyzed these reads using the same methods we used for S. cerevisiae and human and successfully identified the 5′ end of 25S rRNA. However, the 5′ ends were identical between the control and Atdxo1-2 mutant (Fig. 1F). Furthermore, the 5′ ends of the 18S and 5.8S rRNA were also unaffected by the Atdxo1-2 mutation (Fig. 1D,E). This suggests that AtDXO1 is not required for 5′ end maturation of rRNA. Combined with our HEK cell DXO knockout data, this suggests that the S. cerevisiae Dxo1 function in 25S rRNA maturation arose in the fungal lineage.
To start to delineate when during fungal evolution Dxo1 acquired a 25S rRNA maturation function, we next examined S. pombe. Like humans and Arabidopsis, S. pombe has only one family member of the DXO/Dxo1/Rai1 family, rai1+ (also called din1+). We performed PARE on rai1Δ and rai1+ S. pombe strains (generously provided by Ke Zhang Reid, Wake Forest University [Tucker et al. 2016]). Surprisingly, rai1+ is not required for 25S rRNA 5′ end processing (Fig. 1I), because the sequencing reads revealed the same 5′ end in rai1Δ and rai1 + strains. Similarly, the 5.8S and 18S 5′ ends were unaffected by rai1+ (Fig. 1G,H). Thus, despite Saccharomyces and Schizosaccharomyces both being in the phylum ascomycete fungi, they differ in the requirement for a DXO/Dxo1/Rai1 enzyme for 25S rRNA 5′ end maturation. Overall, the comparison of Arabidopsis, human, Schizosaccharomyces, and Saccharomyces 5′ RNA end sequencing data suggests that the requirement of Dxo1 in 25S rRNA maturation arose in the Saccharomyces lineage after it diverged from the other three species.
The 25S/28S rRNA 5′ end is similar across kingdoms
Las1 cleaves 27S pre-rRNA at a site 95 nt upstream of the mature 25S rRNA. The majority of the 95 nt are removed by Rat1, but the last 2–7 are removed by Dxo1 (Fig. 2A). We previously proposed that Dxo1 is required for 25S rRNA maturation in S. cerevisiae because the 5′ end of 25S is base-paired to the 3′ end of 5.8S rRNA (Hurtig and van Hoof 2022). Specifically, such base-paired structures may be incompatible with the requirement of Xrn1/Rat1 family members to have at least three unpaired nucleotides at the 5′ end of a substrate (Fig. 2B; Jinek et al. 2011). The 3 nt single-stranded region mediates the stable association of Xrn1/Rat1 family members with RNA during multiple catalytic cycles, and thus is linked to the processive nature of these enzymes. A corollary is that if Xrn1/Rat1 enzymes generate the mature 5′ end, this mature 5′ end should have a single-strand overhang. We thus considered the possibility that while yeast Dxo1 trims 25S rRNA close to a secondary structure, this close trimming may not occur in other organisms, and the 25S rRNA may contain a longer single-stranded 5′ end. We therefore compared the structures of the Dxo1-dependent rRNA in S. cerevisiae to the DXO/Dxo1/Rai1-independent ones in human, Arabidopsis, and S. pombe (Fig. 2C). Each of the 25S/28S rRNAs contains the conserved sequence 5′GACCYCARR3′ near the 5′ end that base-pairs to 5′YYUGRGyGUC3′ near the 3′ end of the 5.8S rRNA, with the lowercase y bulged out. In the 25S rRNA, this conserved helix with 5.8S is preceded by one single-stranded nucleotide in S. cerevisiae, but 3 or 8 in a dxo1Δ strain. Human, Arabidopsis and S. pombe have either one or two single-stranded nucleotides preceding the conserved 25S/5.8S helix. Importantly, there is no obvious correlation between the length of the single-stranded region at the 5′ end and the requirement for DXO/Dxo1/Rai1 for final processing. Thus, the unique requirement of Dxo1 for 25S 5′ end maturation is more likely due to changes in the rRNA processing machinery than due to changes in the structure of the very 5′ end of the 25/28S rRNA.
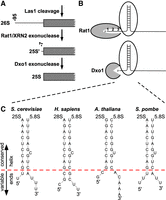
A short single-stranded 5′ end of large rRNA is a conserved feature across kingdoms. (A) 5′ end processing of 25/28S rRNA requires cleavage by Las1, at −95 in S. cerevisiae, followed by exonucleolytic digestion by Rat1 and then by Dxo1. (B) Rat1 family enzymes are processive, at least in part, because they require three single-stranded nucleotides of the 5′ end that mediate continuous binding of the substrate between cycles of catalysis. Although the active site for mouse DXO can accommodate three to four single-stranded nucleotides, whether this is true for Dxo1 is unclear. However, Dxo1 is a distributive enzyme that does not remain associated with the substrate between cycles of catalysis and thus does not require continuous association with an unstructured 5′ end. (C). The 5′ end of the large (25S or 28S) rRNA as defined by the analysis in Figure 1 is base-paired with the 3′ end of 5.8S rRNA. Although there is some variation, the large rRNA has a 1 to 2 nt single-stranded 5′ overhang. There is no obvious reason why the precise 25S 5′ end maturation requires Dxo1 in S. cerevisiae, but not the orthologs in H. sapiens, A. thaliana, or S. pombe.
Dxo1 rRNA processing function arose by gene duplication and neofunctionalization
The difference between S. cerevisiae and S. pombe led us to wonder whether the rRNA processing function of Dxo1 is related to the duplication of DXO1 and RAI1 in the Saccharomyces lineage. The Saccharomyces genome contains hundreds of duplicated gene pairs that arose at once as part of a whole-genome duplication (Wolfe and Shields 1997; Kellis et al. 2004). However, the order of flanking genes indicates that DXO1 and RAI1 did not arise as part of this whole-genome duplication. Instead, this duplication occurred earlier, before the divergence of K. lactis and S. cerevisiae, but after their divergence from Wickerhamomyces and Cyberlindnera (Fig. 3A). We imagined three different scenarios: First, the duplication of DXO1/RAI1 led to a novel function of rRNA processing in the duplicated DXO1 (neofunctionalization). This scenario predicts that (i) K. lactis Dxo1 has the same function as S. cerevisiae Dxo1, (ii) K. lactis Rai1 and S. cerevisiae Rai1 also have the same function, and (iii) the single RAI1 genes from Wickerhamomyces and Cyberlindnera have the same function as S. cerevisiae Rai1. In the second scenario, 25S rRNA processing function is an example of subfunctionalization. This scenario would make the same predictions for the K. lactis proteins, but would instead predict that the single gene from Wickerhamomyces and Cyberlindnera has both the S. cerevisiae Rai1 and S. cerevisiae Dxo1 function. In the third scenario, the evolution of the 25S rRNA function of Dxo1 is unrelated to the duplication and arose later. This scenario predicts that neither K. lactis Dxo1 nor K. lactis Rai1 share the 25S rRNA processing function of S. cerevisiae Dxo1.
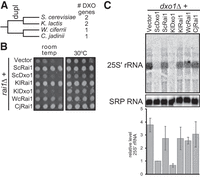
The 25S′ to 25S processing function of Dxo1 arose after duplication of Dxo1 and Rai1. (A) Dxo1 and Rai1 are duplicated in Saccharomyces and Kluyveromyces, but not in Wickerhamomyces or Cyberlindnera, suggesting that duplication occurred in the common ancestor of Saccharomyces and Kluyveromyces after divergence form the other two species. (B) Rai1 function is more ancient than the duplication. Shown is a growth assay of a rai1Δ strain that expresses Rai1 or Dxo1 homologs (from plasmids pRS416, pAv1820, pAv1804, pAv1814, pAv1819, pAv1821, and pAv1815 from top to bottom). The nonduplicated genes from Wickerhamomyces and Cyberlindnera complement indicating that the Rai1 function already existed before duplication. Shown is a representative growth assay from biological duplicates. (C) Dxo1 function arose after the duplication. The dxo1Δ strain was transformed with pRS416, pAv1804, pAv1820, pAv1819, pAv1814, pAv1821, and pAv1815 (from left to right). Shown is a representative northern blot of the 25S′ rRNA and the RNA subunit of the signal recognition particle (SRP; loading control) and the quantitation of two biological replicates.
To distinguish between these possibilities, we used complementation tests in S. cerevisiae. The rai1Δ mutation in S. cerevisiae causes a slow growth phenotype at room temperature (Fig. 3B). This slow growth phenotype can be suppressed by overexpressing the interacting partner Rat1 (Xue et al. 2000) and likely reflects that Rai1 stabilizes Rat1, rather than an enzymatic activity of Rai1. To test which homologs share the S. cerevisiae Rai1 function, we expressed them in a rai1Δ strain and tested for complementation of growth at room temperature (Fig. 3B). As expected, expressing S. cerevisiae Dxo1 does not restore growth to rai1Δ, while expressing S. cerevisiae Rai1 does. Similarly, K. lactis Rai1 complemented rai1Δ, while K. lactis Dxo1 did not. This shows that Rai1 from K. lactis has a similar function to that of S. cerevisiae. This also suggests that, like Dxo1 of S. cerevisiae, K. lactis Dxo1 has diverged in function from Rai1. Next, we looked at the homologs from species that diverged before duplication, Wickerhamomyces ciferrii, and Cyberlindnera jadinii. These species only have one family member of the DXO/Dxo1/Rai1 family and expressing them complements the rai1Δ growth defect, although in the case of W. ciferrii, only partially. This shows that the single family member from W. ciferrii and C. jadinii shares function(s) with Rai1 from S. cerevisiae. We therefore refer to them as WcRai1 and CjRai1. It also shows that the W. ciferrii, C. jadinii, and K. lactis Rai1 each were expressed and functional in yeast. Overall, these data indicate that the Rai1s from these four species are functionally equivalent.
We next tested whether these same homologs can complement the 25S processing function of S. cerevisiae Dxo1. To determine this, we used a dxo1Δ strain and transformed the same plasmids from the rai1Δ complementation assay. We then extracted RNA and performed a northern blot, probing for the 25S′ rRNA (Fig. 3C). As expected, S. cerevisiae Rai1 does not complement the Dxo1 function of 25S′ rRNA processing to 25S, resulting in accumulation of this 25S′ intermediate. Similarly, K. lactis Rai1 does not complement the Dxo1 deletion and results in the accumulation of the 25S′ intermediate. In contrast, K. lactis Dxo1 restores the 25S′ levels to a similar level as S. cerevisiae Dxo1. This demonstrates that the K. lactis Dxo1 can process 25S′, likely because this function arose before the divergence of K. lactis from S. cerevisiae. The single DXO/Dxo1/Rai1 member from W. ciferrii and C. jadinii, WcRai1 and CjRai1, did not complement the 25S rRNA processing defect of dxo1Δ.
The simplest explanation of all these observations is that Dxo1 is an example of neofunctionalization following gene duplication: Rai1 was first duplicated in the common ancestor of K. lactis and S. cerevisiae, and then the functions of the duplicated genes diverged. One copy maintained the Rai1 function, and the other copy evolved a new function in 25S rRNA maturation.
A similar neofunctionalization of Dxo1 occurred in the Candida clade
We have previously noted that Candida albicans also has two DXO/Dxo1/Rai1 family members, but Candida auris does not (Hurtig and van Hoof 2022). Therefore, the two C. albicans genes most likely reflect an independent duplication event from that of the Saccharomyces/Kluyveromyces clade (see also below). To determine which functions the proteins from Candida can carry out, we expressed them in S. cerevisiae as above (Fig. 4). Strikingly, the slow growth of the S. cerevisiae rai1Δ mutant could be complemented by expressing the C. auris gene (CauRai1) or one of the C. albicans genes (CalRai1; Fig. 4B), indicating they share a similar function. The other gene from C. albicans, CalDxo1, did not complement the rai1Δ growth defect. To determine the 25S′ rRNA processing function of these homologs, we performed a northern blot on S. cerevisiae dxo1Δ mutants complemented with the same plasmids as in the rai1Δ complementation assay. The CauRai1 and CalRai1 proteins did not complement the 25S′ rRNA accumulation. In contrast, CalDxo1, which failed to complement rai1Δ, was capable of processing the rRNA to the mature 25S rRNA and complementing dxo1Δ. These results suggest that the C. auris single protein retains the ancestral Rai1 function and does not process 25S′ rRNA. However, after the duplication of the protein in C. albicans, CalDxo1 developed the same capability in rRNA processing as Saccharomyces Dxo1. The fact that two separate gene duplication events resulted in a protein with the same new function of rRNA processing is rather remarkable and illustrates the reproducibility of evolution.
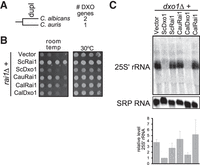
Dxo1/Rai1 is independently duplicated in C. albicans and Dxo1 also acquired 25S′ to 25S processing capacity. (A) Dxo1 and Rai1 are duplicated in C. albicans, but not in C. auris, suggesting that duplication occurred in an ancestor of C. albicans after divergence form C. auris. Note that although the two species are placed in the Candida genus, C. auris is distantly related to true Candida species and more closely related to Clavispora lusitaniae (formerly Candida lusitaniae, Fig. 5). (B) The C. albicans RAI1 gene and the C. auris nonduplicated gene can complement rai1Δ growth phenotype and thus carryout Rai1 function. Shown is a growth assay of a rai1Δ strain that expressed Rai1 or Dxo1 homologs (from plasmids pRS416, pAv1820, pAv1804, pAv1817, pAv1818, and pAv1816 from top to bottom). (C) Dxo1 function arose in C. albicans Dxo1 after the duplication. The dxo1Δ strain was transformed with pRS416, pAv1804, pAv1820, pAv1817, pAv1816, and pAv1818 (left to right). Shown is a representative northern blot of the 25S′ rRNA and the RNA subunit of the signal recognition particle (SRP; loading control) and the quantitation of two biological replicates.
Two separate duplication events were followed by similar sequence changes
To begin to understand the sequence changes that underlie the functional differences between Rai1 and Dxo1, we analyzed the sequences of 44 homologs from the Saccharomycetales (Fig. 5A). This included 34 sequences from 17 species with duplicated genes and 10 sequences from species with a single gene. We generally included one sequence per genus, except for the genus Candida, which includes a number of important pathogens, and the very diverse genus Lachancea. The species with duplicated genes are from two clades (Li et al. 2021), indicated as clades 1 (orange) and 2 (purple) in Figure 5. Saccharomyces and eight other species form clade 1 (orange), while C. albicans and seven other species form clade 2 (purple). The distribution of duplicated Rai1/Dxo1 is consistent with two independent duplications. The alternative, that one duplication occurred in a shared ancestor, would require four independent losses of Dxo1 (in a Wickerhamomyces/Cyberlindnera ancestor, a Clavispora/C. auris/Metschnikowia ancestor, a Yamadazyma ancestor, and a Komagataella/Ogataea ancestor).

The independent duplication of Dxo1/Rai1 in two different clades was followed by parallel sequence changes. (A) 44 Dxo1/Rai1 sequences from 17 budding yeast species with duplicated genes and ten species with a single nonduplicated gene were identified, revealing that the species with duplicated genes fall into two clades (highlighted in orange and purple), indicating two independent gene duplications. The tree is derived from Li et al. (2021). Branch lengths in black are proportional to divergence time estimated in timetree.org. For the branches in gray, no divergence time estimate is available. WGD indicates a well-characterized whole-genome duplication that occurred after and independent of the RAI1/DXO1 duplication. (B) The 44 sequences were aligned using Clustal. The alignment was then split into five sets of sequences: nonduplicated (top center), Dxo1s from clades 1 and 2 (right), and Rai1s from clade 1 and 2 (left). Sequence logos were generated for each of the five subsets. Shown is a sequence logo of “Motif I” of the active site showing that a conserved R/HH motif arose in the Dxo1 of both clades, and that a conserved D became more conserved in the Rai1 of both clades. (C) The R/HH motif of Dxo1 is located in close proximity to the 5′ monophosphorylated RNA. Shown are superimposed structures of K. lactis Dxo1 (dark gray; PDB# 4GPS) and mouse DXO that is bound to an uncleavable substrate analog (5′ monophosphate oligo U with phosphorothioate at the scissile bond; light gray; PDB#4J7M).
We aligned the 44 sequences with Clustal, which revealed several trends. First, the Rai1 proteins from species with duplicated genes were much more conserved than their Dxo1 paralogs (Supplemental Figs. S1 and S2). Specifically, there are only nine residues that are absolutely conserved among the 17 Dxo1 paralogs, while 56 residues are absolutely conserved among the 17 Rai1 paralogs. The nine residues that are 100% conserved in Dxo1 paralogs are a subset of the 56 residues that are 100% conserved in Rai1 paralogs and are the catalytic residues defined by Wang et al. (2015). Thus, while Dxo1 paralogs share a 25S′ processing function, they do not share any absolutely conserved residues that differ from Rai1. The residues that are conserved in Rai1 but absent from Dxo1 include many that are involved in protein–protein interactions with Rat1 (Supplemental Figs. S1 and S2; Xiang et al. 2009). Second, the 17 Rai1 paralogs largely resembled the 10 proteins from species with one homolog, consistent with our functional data that these nonduplicated proteins are functionally interchangeable with Rai1. Third, and most interestingly, the two independent duplications seemed to have been followed by similar sequence changes in both paralogs. In addition to the general trends above, four residues stand out when comparing Dxo1 paralogs and Rai1 paralogs to nonduplicated genes, suggesting that these are especially critical for the functional divergence of Dxo1 and Rai1. Two of the four residues become more conserved in Rai1 and less conserved in Dxo1 after duplication, and the other two residues are conserved only in Dxo1 after duplication.
The two residues that became more conserved in Rai1 after duplication are a D and a Q (D100 and Q134 in S. cerevisiae; Supplemental Figs. S1 and S2). The D residue is just N-terminal to “motif I” of the catalytic site and is the first residue included in Figure 5B. The D is 5 amino acids N-terminal of an absolutely conserved R that is part of motif I. This D residue is moderately conserved in nonduplicated sequences (6 of 10 are D), but 100% conserved in the Rai1 paralogs. The corresponding residue in the Dxo1 paralogs is never a D. Instead, this residue is always a K in clade 2, and mostly a T in clade 1 (Fig. 5B). The Q residue is always Q (6 of 10) or H (4 of 10) in the nonduplicated sequences, always Q in the Rai1 paralogs, but can be either L, M, I, R, K, or N (but never Q or H) in the 17 Dxo1 paralogs (Supplemental Fig. S1). This suggests that these D and Q residues may be important for Rai1 function. Their contribution to Rai1 function is unclear because they are both distant from the active site and unlikely to directly affect catalytic activity as well as distant from the Rat1 interface (Xiang et al. 2009).
The two residues that are conserved only in Dxo1 after duplication are in motif I of the catalytic site (Fig. 5B). In the species without duplication, the conserved R of motif I is followed by a very highly conserved G and a branched amino acid (I, V, or L), and this pattern is conserved in the Rai1 paralogs. In contrast, in the Dxo1 paralogs this GI/V/L motif is replaced by HH in clade 1 and RH in clade 2 (hereafter “the R/HH” motif). To begin to understand the molecular function of the R/HH motif, we compared the X-ray structure of K. lactis Dxo1 (PDB 4GPS) to that of mouse DXO bound to a 5′ monophosphate substrate analog (PDB 4J7M). Strikingly, the first residue of the Dxo1-specific R/HH motif (His163 in KlDxo1, His191 in ScDxo1) is in very close proximity to the 5′ monophosphate (Fig. 5C). The monophosphate RNA substrate differs from all other substrates by carrying two negative charges on the α phosphate. In other substrates, this phosphate is in a pyrophosphate bond and only carries one negative charge, or in the case of 5′ hydroxylated RNA is absent. We speculate that the additional positive charge introduced by the R/HH motif adds positive charge to accommodate the increased negative charge of the 5′ monophosphate of its substrate. Interestingly, biochemical analysis of the K. lactis Dxo1 enzyme has previously highlighted the first histidine of the R/HH motif as important for substrate specificity (Chang et al. 2012). Changing this His to the Rai1-like Gly in KlDxo1 caused a large increase in exonuclease activity on a 5′ triphosphate RNA in vitro but had no effects on decapping a 7mGPPPN RNA or on exonuclease activity on a 5′ monophosphate RNA. We analyzed point mutants of Rai1 and Dxo1, to test whether changing the GI of Rai1 to the HH of Dxo1 was sufficient to convert Rai1 to Dxo1 or vice versa (Supplemental Fig. S3) and observed no effect. Thus, while the previous enzymatic assays and our sequence analysis implicated the R/HH motif, there must be other changes in the proteins that contribute to their functional divergence, which likely include the second His of the R/HH motif and residues lost from Dxo1 (see Discussion).
DISCUSSION
Here we describe the evolutionary origin of the 25S rRNA function of Dxo1. We recently showed that the main function of Dxo1 is to generate the mature 25S rRNA 5′ end from a 25S′ precursor that is extended by 2 or 7 nt (Hurtig and van Hoof 2022). This step in rRNA processing has long been known, but the required enzyme was uncertain. In this paper, we show that this is not an ancient function of Dxo1. Unlike the S. cerevisiae dxo1Δ defect in rRNA processing, the human, A. thaliana, and S. pombe knockouts of their sole DXO/Dxo1/Rai1 gene have no effect on the 5′ end processing of 25/28S rRNA or the other rRNAs. This indicates that the 25S′ processing function of Dxo1 arose in an ancestor of S. cerevisiae after divergence from the S. pombe lineage (an estimated 543 MYA). We considered that this may reflect either a change in the protein function or in the RNA substrate. Because there are no obvious differences in the mature 25S rRNA 5′ end that would affect its processing, the most likely explanation is that the 25S′ processing function arose by a change in Dxo1. Consistent with this, the observation that Xrn1 binds 3 single-stranded nt of its substrate originated from structural and biochemical analysis of the Drosophila Xrn1 and yeast Rat1 (Jinek et al. 2011; Gasse et al. 2015; Fromm et al. 2017) and thus is not restricted to Saccharomyces Xrn1. We thus suspect that the final maturation of the large rRNA in other eukaryotes also requires a distributive 5′ exo, but that 5′ exo is not a DXO/Dxo1/Rai1 family member.
We further narrowed down the origin of this new function by testing whether homologs can complement a S. cerevisiae dxo1Δ defect. We show that the K. lactis Dxo1 homolog, but not homologs from W. ciferrii and C. jadinii, can complement the dxo1Δ defect. This suggests that Dxo1 acquired its 25S rRNA processing, or neofunctionalized, at least 114 MYA, the estimated divergence time of K. lactis from S. cerevisiae. This new function of Dxo1 closely coincides with the duplication of Dxo1 and its paralog Rai1 between 145 and 235 MYA, the estimated divergence times for H. osmophila and C. albicans, respectively. We were initially surprised by these findings, because similar approaches with other genes have found that that subfunctionalization appears to be a more pervasive fate of duplicated yeast genes (van Hoof 2005; Conant and Wolfe 2008; Hurtig et al. 2020; Kuzmin et al. 2022).
Although most eukaryotic genomes include a single DXO/Dxo1/Rai1 gene, the gene is duplicated in D. melanogaster (into Rai and Cutoff genes), C. elegans (into ∼ 10 genes), and in C. albicans. We further investigated the C. albicans duplication and show that one copy can functionally replace S. cerevisiae Dxo1, while the other copy can replace Rai1. We therefore refer to these Candida genes as Dxo1 and Rai1, even though they are not one-to-one orthologs of the yeast genes. The most parsimonious explanation is that an independent duplication occurred in the C. albicans lineage and that this duplication was followed by similar functional changes in the Dxo1 copy. Sequence analysis is consistent with this interpretation. The sequence of the Rai1 copy in both lineages largely resembles those of homologs that diverged before duplication, and we were unable to identify any new conserved amino acids that are unique to the duplicated Rai1s. In contrast, the Dxo1 copies underwent more extensive sequence changes: Many conserved residues were lost, which include many Rat1 interacting residues. The growth defect of rai1Δ we assayed likely reflects the role of Rai1 in stabilizing the essential protein Rat1, and Rai1 enzymatic activity may be unrelated to growth. This conclusion is based on the fact that overexpression of Rat1 or adding a nuclear localization signal onto Xrn1 can suppress the growth phenotype of rai1Δ (Xue et al. 2000). Thus, the inability to complement rai1Δ likely results from a loss of Rat1 interaction. It is therefore not surprising that changing two residues in Dxo1 did not result in rai1Δ complementation (Supplemental Fig. S3). The proteins are also localized differently with Rai1 being nuclear and Dxo1 being cytoplasmic. Rai1 does not have a recognizable nuclear localization signal, but it may be imported as a Rat1/Rai1 complex. Indeed, Rai1-GFP becomes cytoplasmic in a rat1-1 strain (Sydorskyy et al. 2003). Thus, the loss of the Rat1 interacting amino acids may have resulted in Dxo1 being cytoplasmic, where it could access the 25S′ RNA. In addition to lost residues, Dxo1 paralogs gained an R/HH motif near the catalytic site. We speculate that the added positive charges of the R/HH motif may accommodate the additional negative charge on the 5′ monophosphate end of the RNA. We suggest that this particular amino acid change was in part responsible for the gain of the 5′ exoribonuclease activity required for 25S′ processing. Overall, our results and previously published enzymatic data (Chang et al. 2012) suggest that the gain of extra positive charges near the active site optimized exonuclease activity, but that simultaneously other changes occurred that altered protein interactions and subcellular localization, which also contributed to functional divergence.
Our suggestion that similar duplication and neofunctionalization into a 5′ exoribonuclease occurred twice raises the question of whether it occurred in other clades as well. As mentioned above, the D. melanogaster genome contains two DXO/Rai1/Dxo1 family members, Rai and cutoff, but the cutoff gene appears to have lost catalytic activity and taken on a new function in piRNA processing (Pane et al. 2011; Mohn et al. 2014). The C. elegans genome encodes 10 DXO/Rai1/Dxo1 family members, but none of them are strong candidates for a Dxo1-like protein. One of the family members (M01G12.14) does have an HT in the positions corresponding to the R/HH motif, but in preliminary analysis, M01G12.14 failed to complement dxo1Δ. Although such a negative result is inconclusive, M01G12.14 may not have evolved similarly to Dxo1. Overall, we did not find evidence for a Dxo1-like function outside the Saccharomycetales, but it would not be surprising to find similar neofunctionalization elsewhere in the future. An alternative possibility is that perhaps the Saccharomycetales evolved some unknown feature that predisposed them to parallel duplications of Rai1/Dxo1. For example, perhaps the Saccharomycetales lost an unknown 5′ exonuclease, providing selective pressure for the evolution of a Rai1-like gene into a Dxo1-like exoribonuclease to replace the lost gene. To resolve these possibilities, it may be critical to determine the enzyme that matures the 5′ end of human 28S rRNA.
MATERIALS AND METHODS
Human cells, culture, and RNA extraction
HEK293T, including DXO KO, cells have been described previously (Jiao et al. 2017) and were obtained from Mike Kiledjian (Rutgers University) and maintained in DMEM + 10% FBS at 37°C, 5% CO2. RNA was extracted using RNeasy Kit (Qiagen).
S. pombe strains, culture, and RNA extraction
S. pombe strains have been described previously and were obtained from Ke Zhang Reid at Wake Forest (Tucker et al. 2016). S. pombe were grown in YES media. RNA was extracted using the hot phenol RNA extraction protocol, as previously described (He et al. 2008). Two biological replicates were analyzed with identical results.
S. cerevisiae strains, culture, and RNA extraction
S. cerevisiae strains and methods were as described in our previous paper on Dxo1 characterization (Hurtig and van Hoof 2022). RNA was extracted using hot phenol, as previously described (He et al. 2008). RNA gel electrophoresis, blotting and probing were all performed as previously described (Hurtig and van Hoof 2022). Imaging was performed on a Typhoon scanner (Cytiva). Signal from the 25S′ probe (5′ TTTGAGGTCAAACTTTAA 3′) was normalized to that from the SCR1 probe (5′ GTCTAGCCGCGAGGAAGG 3′). Rolling ball background subtraction was performed in ImageQuant.
RNA 5′ end determination by next-generation sequencing
5′ monophosphate containing RNA from human and S. pombe was sequenced (LC Sciences) as described previously (Hurtig and van Hoof 2022). This method uses T4 RNA ligase to ligate an adaptor to RNAs with a 5′ monophosphate to build a sequencing library. This library is then sequenced, and the first nucleotide after the adaptor corresponds to the first nucleotide of the RNA. Similar sequencing reads for Arabidopsis were downloaded from SRA (accession numbers SRR10742163 to SRR10742170). The four biological replicates were analyzed with identical results. Two of those replicates were treated with the plant hormone ABA, and two replicates were untreated.
Reads from all three species (Human, A. thaliana, and S. pombe) were analyzed as described previously (Hurtig and van Hoof 2022). Transcriptome-wide mapping of repetitive sequences such as rDNA can be challenging, and to rule out spurious findings arising from these challenges, we additionally mapped the reads to a reference “genome” file that only contained a single rDNA repeat. Both the genome-wide and rDNA mapping approaches gave comparable findings. Read maps were visualized with IGV. The degradome sequencing reads are available from the SRA under accession number PRJNA1103799.
Plasmids
To express DXO/Dxo1/Rai1 sequences from other yeasts (or M09G12.14), we first cloned S. cerevisiae DXO1 into pRS316 (Sikorski and Hieter 1989), resulting in pAv1804. The coding sequences from other budding yeast species were codon optimized and ordered from Genscript fused to the last 45 bp of the DXO1 promoter (starting with a ClaI site that naturally occurs in the DXO1 promoter) and the DXO1 3′UTR, followed by a SacI site. The gene fragment was then cloned into a ClaI/SacI digested pAv1804. The final result was a series of plasmids that contained the 274 bp DXO1 promoter, the heterologous Dxo1 or Rai1 coding region, and the 293 bp DXO1 3′UTR, with the coding region precisely fused to the DXO1 flanks at the start and stop codons. The point mutant plasmids were generated using Gibson assembly. All plasmids (Supplemental Table S1) were confirmed by sequencing. Yeast transformations were performed as previously described (Gietz and Schiestl 2007), and transformants were grown on SC-URA media (Sunrise Science).
Growth assays
Growth assays were conducted as previously described (Hurtig and van Hoof 2022) on SC-URA. Plates were grown for 2 days at either 30°C or room temperature. The rai1Δ strain shows a subtle growth defect at 30°C that becomes more easily detectable at room temperature.
Protein sequence analysis
The 44 proteins sequences were downloaded from GenBank and aligned using ClustalW (Larkin et al. 2007). The resulting FASTA file (Supplemental Fig. S1) was split into five FASTA files containing either nonduplicated proteins, or Dxo1-like or Rai1-like proteins from clade 1 or clade 2. The five resulting FASTA files were used to generate the sequence logos in Supplemental Figure S2 (Crooks et al. 2004). The K. lactis Dxo1 and mouse DXO structures were overlaid using the UCSF Chimera MatchMaker function (Pettersen et al. 2004).
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
We thank Mike Kiledjian (Rutgers University) and Ke Zhang Reid (Wake Forest University) for generously sharing HEK lines and S. pombe strains. This work has been supported by the National Institute of General Medical Sciences: R35GM141710.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080210.124.
- Received July 30, 2024.
- Accepted September 16, 2024.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








