
tRNAVal allows four-way decoding with unmodified uridine at the wobble position in Lactobacillus casei
- Riko Sugita1,
- Vincent Guérineau2,
- David Touboul3,
- Satoko Yoshizawa4,
- Kazuyuki Takai1 and
- Chie Tomikawa1
- 1Department of Materials Science and Biotechnology, Graduate School of Science and Engineering, Ehime University, 3 Bunkyo-cho, Matsuyama, Ehime 790-8577, Japan
- 2Institut de Chimie des Substances Naturelles, CNRS UPR 2301, Université Paris-Saclay, 91198 Gif-sur-Yvette Cedex, France
- 3Laboratoire de Chimie Moléculaire, CNRS UMR 9168, Ecole Polytechnique, IP-Paris, Route de Saclay, 91120 Palaiseau, France
- 4Université Paris-Saclay, ENS Paris-Saclay, CNRS UMR8113, Laboratory of Biology and Applied Pharmacology (LBPA), 91190 Gif-sur-Yvette, France
- Corresponding author: tomikawa.chie.mm{at}ehime-u.ac.jp
-
Handling editor: Eric Westhof
Abstract
Modifications at the wobble position (position 34) of tRNA facilitate interactions that enable or stabilize non-Watson–Crick base pairs. In bacterial tRNA, 5-hydroxyuridine (ho5U) derivatives xo5U [x: methyl (mo5U), carboxymethyl (cmo5U), and methoxycarbonylmethyl (mcmo5U)] present at the wobble positions of tRNAs are responsible for the recognition of NYN codon families. These modifications of U34 allow base-pairing not only with A and G but also with U, and in some cases, C. mo5U was originally found in Gram-positive bacteria, and cmo5U and mcmo5U were found in Gram-negative bacteria. tRNAs of Mycoplasma species, mitochondria, and chloroplasts adopt four-way decoding in which unmodified U34 recognizes codons ending in A, G, C, and U. Lactobacillus casei, Gram-positive bacteria, and lactic acid bacteria lack the modification enzyme genes for xo5U biosynthesis. Nevertheless, L. casei has only one type of tRNAVal with the anticodon UAC [tRNAVal(UAC)]. However, the genome of L. casei encodes an undetermined tRNA (tRNAUnd) gene, and the sequence corresponding to the anticodon region is GAC. Here, we confirm that U34 in L. casei tRNAVal is unmodified and that there is no tRNAUnd expression in the cells. In addition, in vitro transcribed tRNAUnd was not aminoacylated by L. casei valyl-tRNA synthetase, suggesting that tRNAUnd is not able to accept valine, even if expressed in cells. Correspondingly, native tRNAVal(UAC) with unmodified U34 bound to all four valine codons in the ribosome A site. This suggests that L. casei tRNAVal decodes all valine codons by four-way decoding, similarly to tRNAs from Mycoplasma species, mitochondria, and chloroplasts.
Keywords
INTRODUCTION
tRNA plays a crucial role in protein synthesis, and tRNA modifications facilitate the decoding of codons in mRNA on ribosomes. In particular, modified nucleosides in the anticodon loop modulate decoding during protein synthesis. Modifications at the wobble position (position 34, the first letter of the anticodon) directly facilitate interactions that allow or stabilize non-Watson–Crick base pairs. Modifications at positions 32, 37, and 38, which are part of the anticodon loop or adjacent to the anticodon also act to allow or stabilize non-Watson–Crick base pairs, but do so indirectly (Björk and Hagervall 2014; Rozov et al. 2015; Grosjean and Westhof 2016).
There are modified nucleosides that expand the codon decoding specificity. The 5-hydroxyuridine (ho5U) derivatives xo5U [x: methyl (mo5U), carboxymethyl (cmo5U), and methoxycarbonylmethyl (mcmo5U)] at the wobble position form base pairs with U, A, G, and in some cases with C (Fig. 1A; Näsvall et al. 2007). 5-methoxyuridine (mo5U) was found in Gram-positive bacteria (Murao et al. 1976; Ryu et al. 2018), and 5-carboxymethoxyuridine (cmo5U) and 5-methoxycarbonylmethoxyuridine (mcmo5U) were found in Gram-negative bacteria (Kuchino et al. 1979; Goodenbour and Pan 2006; Björk and Hagervall 2014). 5-carboxymethoxyuridine (cmo5U) is one of the most abundant modified nucleosides in Escherichia coli (Dong et al. 1996; Sakai et al. 2016) and serves to expand the decoding capacity for the NYN (Y, pyrimidine base) codons in four-codon families, i.e., codons of valine, serine, proline, threonine, alanine, and leucine. Furthermore, the xo5U modification at the wobble position in tRNA promotes base-pairing with G and pyrimidine at the third position of codons (Takai et al. 1996; Näsvall et al. 2007).
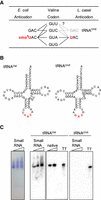
tRNAs for valine codon decoding in L. casei. (A) Codon–anticodon pairing in valine codon. In E. coli, tRNAVal(UAC) bearing cmo5U at the wobble position decodes all four valine codons, and tRNAVal(GAC) decodes the GUU and GUC codons. In L. casei, tRNAVal(UAC) with unmodified or modified U at the wobble position may decode the four valine codons. If tRNAUnd is present in the cells, the tRNA may decode the GUC and GUU codons. (B) Clover-leaf structures of tRNAVal and tRNAUnd of L. casei. Red letters are anticodon regions. (C) Detection of tRNAVal and tRNAUnd from L. casei BL23 by northern hybridization. The small RNA fraction of L. casei was separated by 10% PAGE (7 M urea), and the gel was stained with toluidine blue (left panel). The small RNA fraction (lanes: small RNA), native isolated tRNAVal (lane: native, only in tRNAVal), and in vitro transcribed tRNAVal or tRNAUnd (lanes: T7) were separated by 10% PAGE (7 M urea), and the RNAs were transferred to a membrane. Each tRNA was detected using a 5′-32P-labeled DNA probe with a sequence complementary to tRNAVal or tRNAUnd. Sequences of the DNA probes for tRNA detection are described in Materials and Methods.
The biosynthesis of the xo5U modification has been well-studied in E. coli. The first step of xo5U synthesis is catalysis by TrhP or TrhO to generate 5-hydroxyuridine (ho5U). Prephenate is required for TrhP-dependent ho5U synthesis. TrhO, is a rhodanese family protein that catalyzes ho5U synthesis in an oxygen-dependent manner (Sakai et al. 2019). ho5U34 can be carboxymethylated by CmoB to form cmo5U34. Four tRNA species, namely tRNAAla(UGC), tRNASer(UGA), tRNAPro(UGG), and tRNAThr(UGU), can be further methylated by CmoM to form mcmo5U34 (Murao et al. 1970). In tRNASer(UGA), the ribose of mcmo5U34 is partially methylated to generate 5-methoxycarbonylmethoxy-2'-O-methyluridine (mcmo5Um) (Sakai et al. 2016). In Gram-positive bacteria, ho5U is methylated by TrmR to produce mo5U34 (Murao et al. 1976; Ryu et al. 2018).
Mycoplasma species, mitochondria, and chloroplasts, all of which contain a limited number of genes, adopt four-way decoding in which unmodified U34 recognizes codons ending in A, G, C, and U (Osawa et al. 1992; Rogalski et al. 2008; Alkatib et al. 2012). In particular, in human mitochondria, all four-codon families are decoded by a single tRNA with unmodified uridine at the wobble position (Barrell et al. 1980; Andachi et al. 1987; Inagaki et al. 1995; Suzuki et al. 2020; Lei and Burton 2022).
Lactobacillus casei, a Gram-positive bacterium, does not contain a trmR ortholog in its genome. The genes trhO and trhP, which catalyze ho5U34 synthesis, are not encoded and ortholog genes of the enzymes known to be involved in xo5U synthesis are also absent from the L. casei genome. In addition, only one type of tRNAVal with the anticodon UAC has been identified in L. casei, and it is encoded by three genes that are completely identical in sequence. Intriguingly, there is an undetermined tRNA (tRNAUnd) gene with GAC sequence at the position corresponding to the anticodon region (Fig. 1B). In this study, we investigated whether the tRNAUnd is involved in valine codon decoding and whether the anticodon of tRNAVal undergoes posttranscriptional modification (Fig. 1A). We also investigated how L. casei deciphers the fourfold degenerate codon box for valine (GUN).
RESULTS
Detection of tRNAVal and tRNAUnd in L. casei BL23
L. casei BL23 contains three genes coding for tRNAVal, the anticodon of which is UAC, and the sequences of the tRNA genes are completely identical (Fig. 1B) (GtRNAdb, http://gtrnadb.ucsc.edu). In the database, there is an tRNAUnd gene, and the sequence corresponding to the anticodon region is GAC (Fig. 1B). The expression of tRNAVal and tRNAUnd genes was investigated. Purified small RNAs and isolated tRNAVal from L. casei were analyzed by northern hybridization using 10% PAGE (7 M urea). Hybridization bands derived from tRNAVal were detected in the RNA samples, and the migration patterns were consistent with in vitro transcribed tRNAVal (Fig. 1C). In contrast, attempts to isolate tRNAUnd from the small RNA fraction of L. casei were unsuccessful. In agreement with this result, no hybridization band derived from tRNAUnd was detected in the small RNA fraction (Fig. 1C).
Measurement of aminoacylation of tRNAVal and tRNAUnd transcripts by L. casei ValRS
In our experiment, the expression of tRNAUnd could not be detected. In the event that a small amount of tRNAUnd is expressed in cells, we examined whether tRNAUnd accepts valine. In vitro transcribed tRNAVal was charged with [14C]-valine using L. casei ValRS (Fig. 2A,B). In contrast, tRNAUnd was not charged with valine, and the level of radioactivity incorporated into RNA remained as low as the background level (Fig. 2B), indicating that tRNAUnd does not accept valine. The aminoacylation determinants of tRNAVal by E. coli ValRS are G20, A35, C36, G45, A73, and two base pairs, U4-A69 and U29-A41 (Tardif and Horowitz 2002). The tRNAVal contains all these determinants except U4-A69, and the tRNAUnd contains these determinants except U4-A69 and U29-A41 (Supplemental Fig. S1A). We prepared mutant tRNAUnd transcripts with A4-U69 or C29・C41 substituted with U4-A69 or U29-A41, respectively (Supplemental Fig. S1A,B) and measured the aminoacylation activity using L. casei ValRS. Neither of the mutant tRNAs was aminoacylated (Supplemental Fig. S1C,D). Therefore, there may be other reasons why tRNAUnd is not aminoacylated. Comparing the amino acid sequences of L. casei ValRS and E. coli ValRS, the E. coli ValRS has 16 and 37 amino acid residues inserted into the connective polypeptide 1 (CP1) domain and α-helix bundle domain, respectively (Supplemental Fig. S2A). Crystal structure of Thermus thermophils ValRS complexed with tRNAVal shows that the CP1 domain is in contact with the 3′ CCA end of tRNA, and the α-helix bundle domain contains an anticodon-recognition region (Supplemental Fig. S2B; Fukai et al. 2003). The tRNA determinants may differ between E. coli ValRS, in which 16 or 37 amino acids are inserted into these regions, and L. casei and T. thermophilus ValRSs, in which they are not.
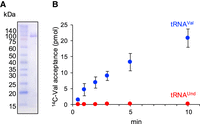
Aminoacylation activity of L. casei ValRS. (A) Purified L. casei ValRS was analyzed by 12% SDS-PAGE, and the gel was stained with Coomassie brilliant blue. The mass of L. casei ValRS is 96 kDa. (B) Aminoacylation activity of L. casei ValRS was measured for transcripts of tRNAVal (blue circles) and tRNAUnd (red circles). Error bars are SEM for three independent experiments.
Nucleoside/nucleotide analysis of tRNAVal by Kuchino's post-label method and mass spectrometry
All known modification enzyme genes for codon expansion are absent from the L. casei genome. Therefore, to clarify whether the anticodon of tRNAVal contains modified nucleosides or remains unmodified, tRNAVal from L. casei was analyzed after isolation using a solid-phase DNA probe method (Supplemental Fig. S3; Tomikawa et al. 2010) and further purification by 10% PAGE (7 M urea) (Fig. 3A). The purified tRNAVal was analyzed by Kuchino's post-label RNA sequencing method (Kuchino et al. 1979). 5′-end 32P-labeled tRNAVal fragments from limited cleavage were separated by 15% PAGE (7 M urea) (Fig. 3B) and extracted from the gel. The resulting fragments were digested with nuclease P1, and the 32P-labeled mononucleotides were analyzed by 2D TLC in two types of solvent systems. U34, A35, and C36 in tRNAVal were identified as unmodified nucleotides. In addition, we found a N6-methyladenosine-5′-monophosphate (pm6A) modification at position 37 (Fig. 3C).
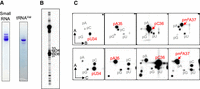
Nucleotide analysis of anticodon region in L. casei tRNAVal by Kuchino's post-labeling method. (A) L. casei tRNAVal was purified from the small RNA fraction. The purified small RNA and tRNAVal were analyzed by 10% PAGE (7 M urea), and the gels were stained with toluidine blue. (B) The purified tRNAVal was partially cleaved by formamide, and then the 5′-end of each fragment was labeled with γ-32P-ATP using T4 polynucleotide kinase. The RNA fragments were separated by 15% PAGE (7 M urea). Numbers correspond to the nucleotide positions in tRNAVal. (C) RNA fragments cleaved by formamide were purified from gel and digested with nuclease P1, and then their 5′ mononucleotides were analyzed by 2D TLC. The TLC analysis was performed in two types of solvent system, namely solvents A and B (upper four panels) or A and C (lower four panels). Solvent A was isobutyric acid:aqueous ammonia:water (66:1:33, v/v/v). Solvent B was isopropanol:HCl:water (70:15:15, v/v/v). Solvent C was 50 mM sodium phosphate, pH 6.8:ammonium sulphate:n-propanol (100:60:2, v/w/v). Nucleotides at positions 34–37 are shown. U34 was unmodified U, and spots of pm6A were detected at position 37.
The isolated tRNAVal was further analyzed for posttranscriptional modifications by MALDI-TOF mass spectrometry after RNase A or RNase T1 treatment. The analysis of the tRNAVal fragments encompassed the entirety of the tRNA sequence. Methylation was detected in the C31–G39 fragment containing the anticodon UAC in the RNase T1 treated sample (m/z = 2892.8) (Fig. 4A; Supplemental Fig. S4 and Table S1). The A37-C40 fragment obtained by RNase A treatment also contained methylation (m/z = 1341.5) (Fig. 4B; Supplemental Table S2). Given that a m6A modification was detected at position 37 using Kuchino's post-label method by 2D TLC (Fig. 3C), the results suggest that the anticodon nucleosides are unmodified. Other canonical modified nucleosides were suggested in the mass spectrometry analysis, namely 4-thiouridine at position 8 (s4U8), 7-methylguanosine at position 46 (m7G46), dihydrouridine (D) at position 16 or 17, and 5-methyluridine at position 54 (m5U54) (Fig. 4A,B; Supplemental Fig. S4 and Tables S1 and S2). Nucleoside analysis of sample treated with nuclease P1 and bacterial alkaline phosphatase (BAP) by Liquid chromatography-high resolution mass spectrometry (LC-HRMS) showed the presence of a pseudouridine (Ψ) (Fig. 4C). The Ψ is likely to be at position 55 because the Ψ55 synthase (truB) gene is present in L. casei. In bacterial tRNA, Ψ has been identified at positions 13, 38, 39, 40, 55, and 65 (Kammen et al. 1988; Nurse et al. 1995; Del Campo et al. 2001; Lecointe et al. 2002; Sprinzl and Vassilenko 2005). The tRNAVal of L. casei lacks the U13, U38, U39, and U40, while it contains the U65. The formation of Ψ65 is attributed to the enzyme TruC in E. coli (Del Campo et al. 2001). However, the L. casei genome does not encode a TruC ortholog. It is therefore reasonable to conclude that the pseudouridine detected in the analysis is derived from the Ψ at position 55. In agreement with MALDI-TOF analysis, we detected ions corresponding to s4U, D, m6A, m7G, and m5U. Genes for the enzymes responsible for these modifications at positions 8, 16–17, 37, 46, and 54, respectively, are also found in L. casei. m5U and m7G were confirmed from HPLC analysis (Supplemental Fig. S5). All the modified nucleosides detected are shown in Figure 4D.
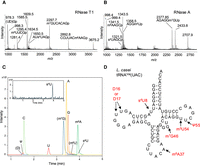
Mass spectrometry analysis of the native L. casei tRNAVal. (A) The MALDI-TOF mass spectrum of the native L. casei tRNAVal digested by RNase T1 shows s4U8 containing fragment AUs4UAGp, D containing fragment (UD)Gp, m6A37 containing fragment CCUUACm6AAGp, m7G46 containing fragment m7GUCACAGp, and m5U54 containing fragment m5UUCGp. (B) The MALDI-TOF mass spectrum of the native L. casei tRNAVal digested by RNase A shows s4U8, m6A37, m7G46, and m5U54. Supplemental Tables S1 and S2 give theoretical and empirical masses of singly protonated ions derived from the RNase T1 and RNase A fragments of the L. casei tRNAVal, respectively. (C) LC-HRMS analysis of nuclease P1 digested L. casei tRNAVal nucleosides. Overlay of chromatograms obtained for A, C, G, U, Ψ, (D), m6A, and s4U detected at m/z of 268.1040, 244.0928, 284.0989, 245.0768 (U and Ψ), 247.0925, 282.1197, and 261.0540, respectively, at the corresponding retention time expected from the authentic nucleoside samples. Ions at m/z of 259 and 298 are annotated as m5U and m7G, respectively. LC-HRMS data for s4U is provided in the inset. (D) Modified nucleosides of the native L. casei tRNAVal detected by mass spectrometry analyses. Detected modified nucleosides and their positions are shown in red letters.
Codon binding properties of native tRNAVal in L. casei ribosomes
Analysis of binding of the ribosome A site by isolated and aminoacylated 14C-Val-tRNAVal was performed using five different mRNAs, containing GUU, GUC, GUA GUG, and CCA located at the A site. The CCA codon corresponds to proline and serves as a negative control (Fig. 5). More than 80% of the aminoacylated tRNAVal bound to GUA and approximately half to GUG. tRNA binding was also observed for GUU, and the activity was not significantly different from that for GUG. Binding activity was also observed for GUC, although binding was very weak (about 20% binding). The results suggest that tRNAVal harboring the UAC anticodon can translate not only GUA and GUG, but also GUU and GUC.
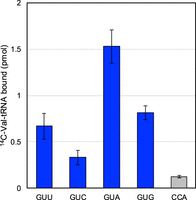
Ribosome A-site binding activity of native L. casei tRNAVal. GUU, GUC, GUA, GUG, and CCA (proline codon) are codons at the A site of the ribosomes. Aminoacylated native L. casei tRNAVal loaded with 14C-Val using ValRS bound not only to mRNA with GUA or GUG at the A site, but also to mRNA with GUU or GUC. Error bars are SEM for three independent experiments.
DISCUSSION
To date, four-way decoding has only been reported in organisms/organelles with reduced genomes, such as some parasitic bacteria (e.g., Mycoplasma species), mitochondria, and chloroplasts (Andachi et al. 1987; Rogalski et al. 2008; Alkatib et al. 2012). In the current work, we report that L. casei, a lactic acid bacterium, also uses four-way decoding in codon reading.
Eukaryotes have tRNA genes numbering between 170 and 570 and require between 41 and 55 tRNA isoacceptors to decode codons (Goodenbour and Pan 2006). In mammalian mitochondria, translation occurs with 22 different tRNAs (Suzuki et al. 2020). Bacteria typically use between 28 and 46 types of tRNA, encoded by genes numbering between 28 and 120 (Ayan et al. 2020; Chan et al. 2021). For example, in the well-studied E. coli, there are 87 tRNA genes, classified into 41 tRNA types (Chan et al. 2021). In Mycoplasma pneumoniae, a member of the Mollicutes class, 62 codons are decoded with only 32 isoacceptor tRNAs using four-way decoding (Simoneau et al. 1993).
The genome of L. casei BL23 encodes a total of 60 tRNA genes, which can be classified into 43 tRNA types. L. casei has more tRNA isoacceptors than E. coli, but there is only one type of tRNAVal, which harbors the anticodon UAC. E. coli has two tRNAVal isoacceptors with the anticodons GAC and UAC. The genome of L. casei also encodes an tRNAUnd with an anticodon sequence of GAC. However, in our experiments, expression of the tRNAUnd was not detected, at least not in L. casei BL23 cultured in rich medium. Moreover, in vitro transcribed tRNAUnd showed no valine acceptance activity using L. casei ValRS. Even if tRNAUnd is expressed in the cell, it is unlikely that this molecule is used in decoding the valine codon GUY unless tRNAUnd contains modifications that positively function for L. casei ValRS. Our results suggest that tRNAVal(UAC) is the only tRNA for decoding the four valine codons despite the absence of known codon expansion modification enzyme genes in the genome. We therefore inferred that there might be incorporation of a novel modification in the anticodon or its neighbor which allows tRNAVal(UAC) to decipher all four valine codons. However, Kuchino's post-labeling method and mass spectrometry analyses showed that the wobble position was an unmodified U. Purified tRNAVal(UAC) with unmodified U34 showed binding activity for all four valine codons at the ribosome A site, and L. casei was shown to use four-way decoding. The results indicate that U at the wobble position is capable of decoding all valine codons but also distinguishes between them. It is known that U-C and U-U wobble base pairs are stereochemically acceptable if adjustment of the backbone chain of the polynucleotide can be accommodated (Grosjean et al. 1978). Bacillus subtilis, which is a Gram-positive bacterium such as L. casei, also has only one type of tRNAPro with the anticodon UGG, which contains mo5U34 (Yamada et al. 2005). Thus, this finding is the first to be reported of four-way decoding in unmodified U34 for bacteria other than Mollicutes in wild-type cells. In four-way decoding in Mycoplasma capricolum, the translation efficiency by tRNAAla(UGC) with unmodified U34 is in the order of GCC, GCG > GCU > GCA (Andachi et al. 1989; Ōsawa 1995). However, M. capricolum rarely uses GCC codon, and the GCG codon usage is much lower than that of GCU and GCA. In general, C and G ending codons in the other four codon family boxes are absent or very rarely used in M. capricolum (Osawa et al. 1992). In L. casei, the binding activity of tRNAVal(UAC) toward the GUA codon is higher than those for the other valine codons, but the usage of the GUA codon is lower than the other three codons (Supplemental Fig. S6). Here, we show that there is no correlation between codon usage frequency and A-site binding activity. Not only in L. casei, but also in some Lactobacillus species, the GUA codon usage is used less frequently than the other valine codons. This infrequent use of the GUA codon may be a strategy used by certain Lactobacillus species, including L. casei, to avoid misreading by tRNAAla(UGC).
Recently, an analysis was performed to understand why tRNA in Mycoplasma species can decode by four-way decoding compared to E. coli tRNA (Kompatscher et al. 2024). Mycoplasma mycoides has only 28 different types of tRNA encoded by 30 tRNA genes (Chan et al. 2021). In M. mycoides, the four glycine codons can be translated on tRNAGly(UCC) which has unmodified uridine at the wobble position. In decoding the four glycine codons, modifications of the tRNAGly(UCC) are not always necessary, but modifications that are not at the anticodon or around the anticodon, such as s4U8 and Ψ55, can increase the activity of the codon translation (Kompatscher et al. 2024). In addition, m6A37 was shown to have no particular effect on four-way decoding. In contrast, in E. coli tRNAGly(UCC), translation activity increased when m6A37 was incorporated (Kompatscher et al. 2024). m6A37 has been proposed to stabilize the codon–anticodon interaction (Grosjean and Westhof 2016) and contributes to the preorganization of the anticodon loop (Agris 2008). In contrast, for the base pair C in the third position of the codon and G34 of the tRNA (C3-G34), m6A37 is not required because the base pair is strong enough, and in Mycoplasma luteus it is always unmodified A37 (Kano et al. 1991). In the study by Kompatscher et al., m6A37 had no significant effect on translation in M. mycoides. However, in our A-site binding experiment, tRNAVal(UAC) containing m6A37 was more efficient at binding to the GUA valine codon than to the other valine codons. Therefore, m6A37 in L. casei tRNAVal(UAC) might positively affect the A3-U34 base pair interaction as observed for E. coli. L. casei tRNAVal(UAC) also contains s4U8 and Ψ55. The presence of these modifications (s4U8 and Ψ55) in L. casei may also facilitate translation.
By comparing the tRNA genes corresponding to NYN in the four-codon box family in species of the genus Lactobacillus for which the sequence of complete tRNA genes has been reported (Table 1), we found the following: all the Lactobacillus sp. have only a single tRNAVal which harbors the anticodon UAC, and they do not have the trmR gene. The exceptions, not listed in Table 1, are L. paragasseri and L. rogosae, which do seem to have the trmR gene and L. paragasseri retains only the tRNAVal(UAC) gene. Although the ortholog for trhP that synthesizes ho5U34 is encoded in the genomes of L. delbrueckii and L. brevis, there are no orthologs for trmR, cmoB, and cmoM. Therefore, for these Lactobacillus species, the four valine codons are likely to be decoded using tRNAVal(UAC) with unmodified U34 (Fig. 6). For proline codons, since these Lactobacillus species do not have tRNAPro with G at the wobble position, which would allow for decoding of CCU and CCC codons (Fig. 6), it is suggested that tRNAPro(UGG) with unmodified U34 decodes the codons in a manner similar to the valine codon. For tRNAAla (Table 1), there are species with a single type of tRNAAla(UGC), two types of tRNAAla(UGC, CGC), or three types of tRNAAla(UGC, CGC, and GGC) (Fig. 6). The GCU and GCC alanine codons are considered to be decoded by tRNAAla(UGC) and/or tRNAAla(GGC). Exceptionally and interestingly, some strains of L. plantarum and L. brevis have only one type of tRNAAla, which harbors the anticodon CGC. In the codon–anticodon rule, unmodified C34 base pairs only with G (Crick 1966). In mitochondria, it has been suggested that unmodified C34 of tRNAMet(CAU) can wobble-base pair with A and G as the first letters in the isoleucine codon (HsuChen et al. 1983), but it is very unlikely to form a C-C base pair. tRNAAla(CGC) containing an unknown modification might be able to translate four alanine codons, or tRNAAla(CGC) with unmodified C34 might translate four codons. In contrast, the tRNAThr of Lactobacillus species has GGU, UGU, and CGU anticodon types, except Lactobacillus hokkaidonensis LOOC260 (Fig. 6). Lactobacillus species appear to have a unique translation system, but further research is needed.
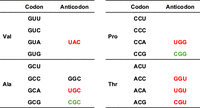
The tRNAs for decoding NYN in the four codon family boxes in the genus Lactobacillus. From the known tRNA genes in the genus Lactobacillus, tRNAs corresponding to Val, Pro, Ala, and Thr codons are shown. The tRNA anticodons present in all Lactobacillus species are shown in red letters, and in some cases the green highlighted anticodon tRNAs are present in addition to the red anticodon tRNAs. In a very small number of species, a tRNAAla harboring the anticodon GGC is present in addition to UGC and CGC anticodon tRNAAla.
tRNA genes corresponding to NYN in the four-codon box family among species of the genus Lactobacillus for which the sequence of complete tRNA genes has been reported
MATERIALS AND METHODS
Purification of small RNA and tRNAVal from L. casei BL23
The culture source of L. casei BL23 was a kind gift from Luis Bermudez–Humaran (French National Institute for Agricultural Research). L. casei was grown on Man–Rogas–Sharpe (MRS; Difco Lactobacilli MRS broth) agar (containing 1.5% agar, w/v) or cultured statically in MRS broth at 37°C. The cells were harvested by centrifugation at OD590 of 1.0. L. casei small RNA was extracted using ISOGEN II (Nippon Gene) according to the manufacturer's manual with slight modifications. Frozen L. casei cells (1 g) were ground in a chilled mortar for 10–20 min. The ISOGEN II (4 mL) was added to the mortar, and cells were further ground for 10–20 min. The cell suspension was transferred to a tube, and then treated according to the ISOGEN II manual. The isolated small RNA was analyzed by 10% PAGE (7 M urea). tRNAVal(UAC) was isolated from the extracted small RNA using a solid-phase DNA probe method described in our previous report (Tomikawa et al. 2010). The sequences of 3′-biotinylated DNA probes are described in Supplemental Table S3, and the position of hybridization on the tRNAs is shown in Supplemental Figure S3. The isolated tRNA was analyzed by 10% PAGE (7 M urea). Attempts to purify tRNAUnd were performed similarly.
Analysis of L. casei tRNAVal and tRNAUnd by northern hybridization
Purified small RNA, tRNAVal and in vitro transcribed tRNAs were separated by 10% PAGE (7 M urea) and transferred to a membrane (Hybond-N+, GE Healthcare) by electro-blotting. The tRNA transcripts were prepared by run-off transcription using T7 RNA polymerase (Hori 2010) and synthetic DNA oligonucleotides shown in Supplemental Table S3. Northern hybridization was performed with hybridization buffer (GE Healthcare) and 5′-32P-labeled DNA probe at 50°C. The DNA probe sequences are as follows: tRNAVal, 5′-AGG CAG ACG CTC TCC CAA CTG-3′; tRNAUnd (CAC), 5′-GTC TTC CGG GTT CCC ACT AGC C-3′. The hybridized bands were monitored using a Typhoon FLA7000 (GE Healthcare).
Cloning and expression of L. casei ValRS
The gene for L. casei ValRS, valS, was amplified by PCR from L. casei genomic DNA using KOD One DNA polymerase (Toyobo). A hexahistidine-tag (6xHis) was inserted into the N terminus of the obtained PCR fragment (Supplemental Table S3). The fragment and a pET22b PCR fragment were assembled using NEBuilder HiFi assembly (New England BioLabs). The original 6xHis sequence of pET22b (Novagen) was removed in the amplified PCR fragment. The resulting expression plasmid was named pET22b-His-LcValRS. E. coli Rosetta 2(DE3) was transformed with the pET22b-His-LcValRS plasmid, and the expression of ValRS was induced by adding 1 mM isopropyl β-D-thiogalactopyranoside (IPTG) at 37°C for 4 h. Harvested cells were stored at −80°C.
Purification of L. casei ValRS
The cells expressing L. casei ValRS (0.8 g) were resuspended in 4 mL buffer A (50 mM Tris-HCl, pH 7.5, 500 mM NaCl, and 6 mM β-mercaptoethanol) and then disrupted with an ultrasonic processor (Sonics and Materials). Supernatant from the resulting cell lysate prepared by centrifugation was applied to a Ni-Sepharose high-performance column (GE Healthcare) equilibrated in buffer A and separated using a 0–500 mM imidazole linear gradient. The fractions containing ValRS were pooled and dialyzed against buffer B (50 mM Tris-HCl, pH 7.5, 50 mM NaCl, and 6 mM β-mercaptoethanol). The dialyzed sample was applied to a Q-Sepharose Fast Flow column (GE Healthcare) equilibrated with buffer B and then separated using a 50–500 mM NaCl linear gradient. The fractions containing ValRS were collected and concentrated using a Vivaspin15 (Sartorius) centrifugal filter device (MWCO = 10,000). The purified ValRS was stored at −30°C in buffer B containing 50% (v/v) glycerol. The concentration of purified ValRS was determined using a Bradford assay. The purity of the protein was monitored by SDS-PAGE.
Assay of aminoacylation of tRNAVal and tRNAUnd transcripts by L. casei ValRS
tRNAVal and tRNAUnd transcripts were aminoacylated with [14C(U)]-Val (9.62 GBq/mmol) from Moravek Biochemicals using purified L. casei ValRS. Reaction mixtures containing 100 mM Tris-HCl, pH 7.5, 5 mM MgCl2, 20 mM KCl, 2 mM ATP, 3.3 µM tRNA transcripts, and 7.7 µM [14C(U)]-Val were preincubated for 5 min at 37°C. The reaction was started by adding 0.1 µM ValRS to the reaction mixture (total volume 50 µL). Forty-five microliters of the reaction mixture was spotted onto a 3MM filter (Whatman), and the filter was immediately soaked in ice-cold 5% trichloroacetic acid (TCA) at appropriate time intervals. The filters were washed three times with ice-cold 5% TCA and twice with 100% ethanol and dried. Radioactivity was quantified on a liquid scintillation counter (AccuFLEX LSC-8000, Hitachi).
Nucleoside analysis of native L. casei tRNAVal
To determine whether the U at position 34 in tRNAVal is modified or unmodified, the anticodon nucleotides of isolated tRNAVal were analyzed using Kuchino's post-labeling method (Kuchino et al. 1979) with slight modifications as in our previous report (Uesugi et al. 2022). Briefly, limited cleavage of the isolated and gel-purified tRNAVal (0.03 A260 units) was performed with formamide (98.5%) at 90°C for 40 sec. The 5′ end of the RNA samples recovered by ethanol precipitation were phosphorylated using T4 polynucleotide kinase (New England BioLabs) and [γ-32P]-ATP (PerkinElmer). The 5′-32P-labeled RNA samples were analyzed by 15% PAGE (7 M urea, 40 cm gel) and then anticodon nucleotides were identified by 2D-TLC (Keith 1995). 5′-32P-labeled nucleotides were monitored using a Typhoon FLA7000 (GE Healthcare).
Mass spectrometry analyses of native L. casei tRNAVal
After purification using the solid-phase DNA probe method, the native tRNAVal was analyzed by mass spectrometry as described previously (Tomikawa et al. 2018). Forty picomoles of native tRNAVal was digested with 50 units of RNase T1 (Thermo Scientific) in 25 mM ammonium acetate, pH 5.3 or with 50 ng of RNase A in 20 mM ammonium acetate, pH 7.5 for 1 h at 37°C. The residual cyclic 2′,3′-phosphates formed were removed by the addition of HCl to a final concentration of 0.1 M and incubation at room temperature for 5 min. A 1 µL aliquot of the digest was mixed with 9 µL 3-Hydroxypicolinic acid (40 mg/mL in water:acetonitrile 50:50), and 1 µL of this mixture was spotted on the MALDI plate and air-dried (“dried droplet” method). MALDI-TOF MS analyses were performed on a MALDI-TOF/TOF ultrafleXtreme (Bruker Daltonics). Acquisitions were performed in reflectron positive ion mode.
For nucleoside composition analysis, 4 µg of purified L. casei tRNAVal was digested with 0.6 units of Nuclease P1 (Boehringer) and 0.1 units of bacterial alkaline phosphatase BAP C75 (Takara) in 50 mM ammonium acetate, pH 7. The reaction mixture was incubated for 45 min at 37°C. The LC-HRMS experiments were performed with an LC 1260 Prime (Agilent Technologies) coupled to a high-resolution tandem mass spectrometer QTOF 6546 (Agilent Technologies). The column (HSS T3, 150 mm × 2.1 mm × 2.7 µm, Waters) was chosen for an efficient separation without the use of any buffer such as ammonium acetate in the mobile phase. The elution gradient started from 100% water + 0.1% formic acid to 50% acetonitrile in 8 min, then 100% acetonitrile during 2 min, and finally back to the initial conditions. The injected volume was fixed at 2 µL. For electrospray (ESI) analysis, mass spectra were recorded in positive ion mode with the following parameters: gas temperature 325°C, drying gas flow rate 10 L.min−1, nebulizer pressure 30 psi, sheath gas temperature 400°C, sheath gas flow rate 10 L.min−1, capillary voltage 3500 V, nozzle voltage 500 V, fragmentor voltage 110 V, skimmer voltage 45 V, octopole 1 RF voltage 750 V. For ESI, internal calibration was achieved with two calibrants, purine and hexakis (1 h,1 h, 3 h-tetrafluoropropoxy) phosphazene (m/z 121.0509 and m/z 922.0098), providing a high mass accuracy better than 3 ppm.
Ribosome A-site binding assay
L. casei ribosomes were prepared according to the literature (Belinite et al. 2021) with slight modifications. L. casei cells were harvested at mid-log phase (OD590 0.6), washed in buffer C (20 mM Hepes-KOH, pH 7.5, 21 mM magnesium acetate, 100 mM NH4Cl, 1 mM dithiothreitol, and 1 mM EDTA), and centrifuged at 7000g for 10 min. Twice the cell weight of alumina was added to the washed cells, which were then ground in a mortar precooled at −80°C, and resuspended in buffer C. The lysate was centrifuged at 30,000g for 90 min to remove debris. The supernatant was loaded onto a 37.6% sucrose cushion prepared in buffer D (10 mM Hepes-KOH, pH 7.5, 25 mM magnesium acetate, 500 mM KCl, 1 mM dithiothreitol, 0.5 mM EDTA, and 37.6% sucrose) and centrifuged for 15 h at 158,400g in a P70AT rotor (Hitachi) to obtain a crude ribosomal pellet. This pellet was resuspended in buffer E (10 mM Hepes-KOH, pH 7.5, 10.5 mM magnesium acetate, 100 mM KCl, 1 mM dithiothreitol, and 0.5 mM EDTA). The resuspended sample was loaded onto 10%–40% (w/v) sucrose density gradients in buffer E and separated by ultracentrifugation in a P28S swing rotor (Hitachi) for 15 h at 38,600g. The 70S ribosome fractions were collected. The ribosomes were pelleted in a P70AT rotor for 15 h at 213,000g. The resultant pellet was washed twice with 1 mL of buffer F (10 mM Hepes-KOH, pH 7.5, 10 mM magnesium acetate, 30 mM NH4Cl, and 1 mM dithiothreitol) and resuspended in buffer F. The aliquoted 70S samples were snap-frozen in liquid nitrogen and stored at −80°C.
The A-site binding assay was performed accordingly to the literature (Pape et al. 1998; Yoshizawa et al. 1999; Kirino et al. 2004; Köhrer et al. 2014). The mRNAs containing GUU, GUC, GUA, GUG, or CCA codons in the A site were prepared by in vitro transcription using T7 RNA polymerase (Supplemental Table S3). The mRNA transcript sequences (A-site codon underlined) were 5′-G GGU UAA CUU AAG AAA GGA GGU UAU ACU GAC NNN UAA CUG CAG AAA AAA-3′, in which NNN indicates GUU, GUC, GUA, GUG, or CCA. Aminoacylated tRNA was prepared as follows: isolated tRNAVal was aminoacylated with [14C(U)]-Val (9.62 GBq/mmol) from Moravek Biochemicals using purified L. casei ValRS as described above.
Frozen ribosomes were thawed and incubated at 42°C for 10 min before use. The P site was first filled with tRNAAsp. The 10 µL reaction mixture contained 10 pmol L. casei ribosomes, 1 µg mRNA, and 23 pmol tRNAAsp in buffer G (50 mM Tris-HCl, pH 7.5, 6.5 mM MgCl2, 60 mM KCl, 1 mM DTT, and 0.5 mM spermine). The mixture was incubated at 37°C for 20 min. Then, 8.3 pmol of tRNA (1.85 pmol aminoacylated) was added and incubated at 37°C. After 20 min, the reaction was stopped with 0.5 mL of ice-cold buffer G. The solution was spotted under vacuum on a nitrocellulose membrane (Sartorius Stedim Biotech, 2.5 cm diameter, 0.45 µm pore size) and washed twice with 1 mL ice-cold buffer G. Ribosome-bound tRNA was quantified on a liquid scintillation counter (AccuFLEX LSC-8000, Hitachi).
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
We thank Dr. Hiroyuki Hori for the helpful discussion. Experiments using radioisotopes were carried out with the support of the Division of Applied Protein Research Support, the Advanced Research Support Center (ADRES) at Ehime University. This work was financially supported by Grants-in-Aid for Scientific Research (19K06491 to C.T.) from the Japan Society for the Promotion of Science, Mishima Kaiun Memorial Foundation (to C.T.), Institute for Fermentation (G-2024-2-043 to C.T.), and the Centre National de la Recherche Scientifique (CNRS) (to S.Y.).
Author contributions: C.T. designed research. R.S., V.G., D.T., S.Y., and C.T. performed experiments. C.T., R.S., V.G., D.T., S.Y., and K.T. analyzed data. C.T., S.Y., R.S., V.G., D.T., and K.T. wrote the paper.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080155.124.
- Received June 21, 2024.
- Accepted September 3, 2024.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵Köhrer C, Mandal D, Gaston KW, Grosjean H, Limbach PA, Rajbhandary UL. 2014. Life without tRNAIle-lysidine synthetase: translation of the isoleucine codon AUA in Bacillus subtilis lacking the canonical
 . Nucleic Acids Res 42: 1904–1915. doi:10.1093/nar/gkt1009
. Nucleic Acids Res 42: 1904–1915. doi:10.1093/nar/gkt1009 - ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵








