
Duf89 abets lncRNA control of fission yeast phosphate homeostasis via its antagonism of precocious lncRNA transcription termination
- 1Molecular Biology Program, Sloan Kettering Institute, New York, New York 10065, USA
- 2Gerstner Sloan Kettering Graduate School of Biomedical Sciences, New York, New York 10065, USA
- 3Department of Microbiology and Immunology, Weill Cornell Medical College, New York, New York 10065, USA
- Corresponding authors: bschwer{at}med.cornell.edu, s-shuman{at}ski.mskcc.org
Abstract
Fission yeast phosphate homeostasis gene pho1 is actively repressed during growth in phosphate-rich medium by transcription in cis of a long noncoding (lnc) RNA from the 5′ flanking prt(nc-pho1) gene. Pho1 expression is: (i) derepressed by genetic maneuvers that favor precocious lncRNA 3′-processing and termination, in response to DSR and PAS signals in prt; and (ii) hyperrepressed in genetic backgrounds that dampen 3′-processing/termination efficiency. Governors of 3′-processing/termination include the RNA polymerase CTD code, the CPF (cleavage and polyadenylation factor) complex, termination factors Seb1 and Rhn1, and the inositol pyrophosphate signaling molecule 1,5-IP8. Here, we present genetic and biochemical evidence that fission yeast Duf89, a metal-dependent phosphatase/pyrophosphatase, is an antagonist of precocious 3′-processing/termination. We show that derepression of pho1 in duf89Δ cells correlates with squelching the production of full-length prt lncRNA and is erased or attenuated by: (i) DSR/PAS mutations in prt; (ii) loss-of-function mutations in components of the 3′-processing and termination machinery; (iii) elimination of the CTD Thr4-PO4 mark; (iv) interdicting CTD prolyl isomerization by Pin1; (v) inactivating the Asp1 kinase that synthesizes IP8; and (vi) loss of the putative IP8 sensor Spx1. The findings that duf89Δ is synthetically lethal with pho1-derepressive mutations CTD-S7A and aps1Δ—and that this lethality is rescued by CTD-T4A, CPF/Rhn1/Pin1 mutations, and spx1Δ—implicate Duf89 more broadly as a collaborator in cotranscriptional regulation of essential fission yeast genes. The duf89-D252A mutation, which abolishes Duf89 phosphohydrolase activity, phenocopied duf89+, signifying that duf89Δ phenotypes are a consequence of Duf89 protein absence, not absence of Duf89 catalysis.
Keywords
- Pol2 CTD code
- Schizosaccharomyces pombe
- cleavage and polyadenylation factor
- inositol pyrophosphate dynamics
INTRODUCTION
Fission yeast phosphate homeostasis entails a transcriptional response to phosphate availability affecting genes that comprise a PHO regulon. The PHO genes specify proteins involved in extracellular phosphate acquisition, these being a cell surface acid phosphatase Pho1, an inorganic phosphate transporter Pho84, and a glycerophosphate transporter Tgp1 (Carter-O'Connell et al. 2012). pho1, pho84, and tgp1 are actively repressed during growth in phosphate-rich medium by the transcription in cis of a long noncoding (lnc) RNA from the respective 5′ flanking genes: prt(nc-pho1), prt2, and nc-tgp1 (Shuman 2020). Transcription of the upstream PHO lncRNA interferes with the downstream PHO mRNA genes by displacing the activating transcription factor Pho7 from its binding site(s) in the mRNA promoters that overlap the lncRNA transcription units. Whereas shutting off the lncRNA promoters suffices to turn on expression of the PHO mRNAs, the sensing and signaling pathway(s) that result in cessation of PHO lncRNA synthesis during phosphate starvation are not known.
Nonetheless, there has been substantial progress via genetics in elucidating influences on the PHO regulon in phosphate-replete cells. In a foundational study, the Wykoff laboratory screened a fission yeast gene deletion collection for mutations that either reduced or increased Pho1 acid phosphatase expression under phosphate-rich conditions (Henry et al. 2011). Subsequent work showed that PHO lncRNA 3′-processing and termination is a key control point in PHO mRNA repression, that is, tandem lncRNA-mRNA transcriptional interference can be tuned by increasing or decreasing the frequency with which Pol2 terminates lncRNA transcription prior to encounter with the mRNA promoter (Shuman 2020). Events that enhance “precocious” termination of lncRNA transcription result in derepression of PHO mRNA expression in phosphate-replete cells and those that reduce the probability of lncRNA termination prior to the mRNA promoter result in hyperrepression of the flanking PHO mRNAs relative to their basal levels. Precocious lncRNA termination is favored by PHO derepressive Pol2 Rpb1-CTD phospho-site mutations S7A and S5A, by a Pol2 Rpb1-N494D mutation that slows elongation rate, by a gain-of-function mutation in the Pol2 termination factor Seb1, by deletion of Erh1, and by deletion of the 14-3-3 protein Rad24 (Schwer et al. 2014, 2015a,b, 2020, 2021; Chatterjee et al. 2016; Sanchez et al. 2018a,b; Yague-Sanz et al. 2020; Garg et al. 2022). lncRNA termination is antagonized by PHO hyperrepressive Pol2 CTD phospho-site mutant T4A, by loss-of-function mutations of the cleavage and polyadenylation factor (CPF) complex, CTD prolyl isomerase Pin1, and termination factor Rhn1 (Chatterjee et al. 2016; Sanchez et al. 2018a,b, 2020).
A distinctive feature of eukaryal cellular physiology is the participation of inositol pyrophosphates in the phosphate starvation response, albeit in ways that are unique to each model system studied (Lee et al. 2008; Azevedo and Saiardi 2017; Dong et al. 2019; Sanchez et al. 2019; Zhu et al. 2019). The inositol pyrophosphates IP7 and IP8 are generated from phytic acid (IP6) by the action of inositol polyphosphate kinases. There are two cellular forms of IP7 that differ according to whether the pyrophosphate moiety is at the 1 or 5 position of the inositol ring; IP8 is pyrophosphorylated at both ring positions (Figs. 1C, 5C). Inositol pyrophosphate dynamics are dictated by a balance between inositol polyphosphate kinases and several pyrophosphatase enzymes that remove the inositol pyrophosphate β-phosphate groups. The initial clues that inositol pyrophosphates might be involved in fission yeast phosphate homeostasis emerged from the Wykoff laboratory screen and follow-up studies in which a deletion of asp1 (which encodes a kinase that synthesizes inositol 1-pyrophosphates) was found to hyperrepress pho1 under phosphate-replete conditions and a deletion of aps1 (which encodes a Nudix-family pyrophosphatase) derepressed pho1 under phosphate-replete conditions (Henry et al. 2011; Estill et al. 2015; Sanchez et al. 2019). Asp1 is a bifunctional enzyme composed of an N-terminal kinase domain that converts 5-IP7 to 1,5-IP8 and a carboxy-terminal pyrophosphatase domain that converts 1,5-IP8 back to 5-IP7 (Fig. 1C; Pascual-Ortiz et al. 2018; Dollins et al. 2020; Benjamin et al. 2022). Asp1 can also phosphorylate IP6 to yield 1-IP7 and de-phosphorylate 1-IP7 back to IP6. The in vivo effect of an asp1Δ null allele or a kinase-dead asp1-D333A allele is to eliminate intracellular IP8 and 1-IP7 and to increase the level of 5-IP7; the in vivo effect of a pyrophosphatase-defective asp1-H397A allele is to increase the level of IP8 (Pascual-Ortiz et al. 2018; Dollins et al. 2020). Aps1 is a Nudix-family pyrophosphatase that converts 1,5-IP8 to 1-IP7 or, with lower flux, to 5-IP7 (Safrany et al. 1999; Kilari et al. 2013; Zong et al. 2021).
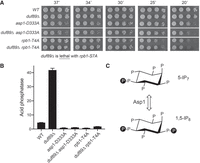
duf89Δ derepression of Pho1 expression depends on inositol 1-pyrophosphate synthesis and the Pol2 CTD Thr4 mark. (A) Growth of S. pombe strains with the indicated duf89, asp1, and rpb1 alleles. Cells were inoculated in YES broth and grown at 30°C. Exponentially growing cultures were adjusted to A600 of 0.1 and aliquots (3 µL) of serial fivefold dilutions were spotted on YES agar and then incubated at the temperatures specified. (B) The indicated fission yeast strains were grown to A600 of 0.5 to 0.8 in liquid culture in YES medium at 30°C. Cells were then harvested, washed with water, and assayed for Pho1 acid phosphatase activity by conversion of p-nitrophenylphosphate to p-nitrophenol. Activity is expressed as the ratio of A410 (p-nitrophenol production) to A600 (input cells). (C) Structures of 5-IP7 and 1,5-IP8 are shown. Asp1 kinase converts 5-IP7 to IP8 and the Asp1 pyrophosphatase reverses this process.
Metabolite control of fission yeast phosphate homeostasis by IP8 is exerted through the 3′-processing/termination machinery and the Pol2 CTD code (Sanchez et al. 2019). Increasing IP8 via an Asp1 pyrophosphatase active site mutation derepresses the PHO regulon and leads to precocious termination of prt lncRNA synthesis in a manner dependent on CPF subunits, termination factor Rhn1, and CTD Thr4. Also, pho1 derepression by CTD-S7A depends on IP8 synthesis by the Asp1 kinase. Simultaneous inactivation of the Asp1 and Aps1 pyrophosphatases is lethal, signifying that too much IP8 is toxic, but this lethality is suppressed by mutations of CPF subunits and CTD mutation T4A. IP8 toxicosis is also suppressed by deleting Spx1, a protein composed of an inositol pyrophosphate-binding SPX domain and a RING ubiquitin ligase domain (Schwer et al. 2022). Spx1 is proposed to act as a transducer of IP8 signaling to the 3′-processing/transcription termination machinery (Schwer et al. 2022). Failure to synthesize IP8 via Asp1 kinase mutation results in pho1 hyperrepression. Thus, IP8 status is an integral part of the termination-centric governance of transcriptional interference in the PHO regulon. Findings of synthetic lethality of asp1Δ with CPF subunit mutations argues that IP8 plays an important role in essential 3′-processing/termination events, albeit in a manner genetically redundant to CPF. These results establish a novel action for IP8 in cell physiology as an agonist of Pol2 transcription termination.
We hypothesize that there are additional influences on inositol pyrophosphate dynamics and/or transcriptional interference in fission yeast that await discovery and that the sensitivity of the PHO regulon to changes in inositol pyrophosphate metabolism might provide inroads to such discovery. Here, we follow up on the observation by Henry et al. (2011) that Pho1 expression is elevated by deletion of the SPCC1393.13 gene that encodes a member of the DUF89 (domain of unknown function 89) protein family. Whereas the significance of this finding was unclear at the time, recent studies highlight DUF89 proteins as a novel clade of phosphohydrolase enzymes found in all three domains of life (Huang et al. 2016).
Our group has shown that purified recombinant fission yeast SPCC1393.13 protein (renamed Duf89) is a cobalt/nickel-dependent phosphohydrolase with vigorous activity against p-nitrophenylphosphate (a generic phosphomonoester), which it hydrolyzes with a kcat of 9.7 s−1. Duf89 also hydrolyzes the phosphoanhydride bond of inorganic pyrophosphate with a kcat of 32 min−1 (Sanchez et al. 2022). We identified a paralogous cobalt/nickel-dependent phosphatase/pyrophosphatase enzyme in fission yeast, SPAC806.04c (renamed Duf8901), but found that its deletion does not influence Pho1 expression (Sanchez et al. 2022).
Here we conduct a genetic and transcriptomic interrogation of fission yeast Duf89. We show that: (i) Duf89 deletion results in derepression of pho1 in a manner that depends on Asp1 kinase, CPF/Rhn1/Pin1, the Pol2 CTD Thr4 mark, and Spx1; (ii) derepression of pho1 by duf89Δ ensues from squelching the production of interfering prt lncRNA; (iii) absence of Duf89 is synthetically lethal with a deletion of the Aps1 pyrophosphatase; (iv) duf89Δ and aps1Δ have highly concordant effects on the fission yeast transcriptome, including derepression of the three-gene PHO regulon; (v) duf89Δ aps1Δ synthetic lethality is suppressed by mutations in Asp1 kinase and CPF/Rhn1, and by loss of the CTD Thr4 mark; (vi) duf89Δ rescues the synthetic lethality of a CPF dis2Δ ssu72-C13S double mutant; and (vii) increased duf89 gene dosage hyperrepresses Pho1 expression. These results implicate Duf89 as an antagonist of precocious transcription termination.
RESULTS
duf89Δ derepresses Pho1 expression
A fission yeast duf89Δ strain grows as well as a wild-type control strain on YES agar at all temperatures, as assessed by colony size (Fig. 1A). The basal Pho1 cell-surface acid phosphatase activity of phosphate-replete wild-type cells was derepressed by ninefold in duf89Δ cells (Fig. 1B). The loss of Duf89 might derepress pho1 expression either by: (i) reducing the activity of the prt lncRNA promoter that drives transcription interference with the pho1 promoter; or (ii) increasing the intrinsic activity of the pho1 mRNA promoter independent of lncRNA synthesis. To address these issues, we used a plasmid-borne prt–pho1 reporter (Fig. 2A) that was introduced into fission yeast cells in which the chromosomal pho1 gene was deleted. This reporter responds faithfully to known homeostatic controls over the native pho1 locus (Chatterjee et al. 2016; Sanchez et al. 2019). We found that the prt–pho1 reporter was responsive to ablation of Duf89, whereby Pho1 levels under phosphate-replete conditions were derepressed by approximately fourfold in duf89Δ cells compared to the wild-type duf89+ control (Fig. 2D). We then tested a mutated version of the prt–pho1 reporter construct in which the prt promoter is inactivated by nucleotide changes in the HomolD and TATA box elements that drive prt lncRNA synthesis (Fig. 2B; Chatterjee et al. 2016). This mutant reporter provides a readout of the intrinsic activity of the pho1 promoter, without interference by transcription of the flanking prt lncRNA. The Pho1 activity of the mutant plasmid in wild-type cells is high (i.e., derepressed) and is not different from the Pho1 activity in duf89Δ cells (Fig. 2E), thereby indicating that the derepressive effect of duf89Δ on pho1 expression from the wild-type prt–pho1 locus is not caused by up-regulation of the pho1 promoter per se. The effect of duf89Δ on the prt promoter was assessed using a different plasmid reporter (Fig. 2C) in which the prt promoter directly drives expression of the pho1 ORF. Pho1 expression from this plasmid was the same in wild-type and duf89Δ cells (Fig. 2F), signifying that the derepression of native pho1 by duf89Δ is not caused by decreased activity of the prt promoter. These results raise the prospect that duf89Δ affects pho1 expression by enhancing precocious termination of prt lncRNA synthesis.
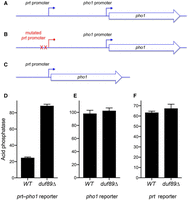
Does duf89Δ affect the prt lncRNA or pho1 mRNA promoters? (A) Schematic of the plasmid-borne prt-pho1 reporter in which pho1 expression is repressed by prt lncRNA transcription. (B) A reporter of pho1 promoter activity in which prt lncRNA transcription is abolished by mutations (indicated by X) in the HomolD and TATA box elements in the prt promoter (Chatterjee et al. 2016). (C) A reporter of prt promoter activity in which the prt promoter directly drives transcription of the pho1 gene. (D–F) The indicated reporter plasmids were transfected into duf89+ (WT) or duf89Δ strains in which the chromosomal pho1 locus was deleted. Transformants were selected and single colonies of individual transformants were pooled (≥20) and grown in plasmid-selective liquid medium to A600 of 0.5–0.8. Aliquots were harvested for acid phosphatase activity measurements. Each datum in the bar graph is the average of assays using cells from three independent cultures ±SEM.
Derepression of pho1 expression by duf89Δ ensues from squelching the production of interfering prt lncRNA
The prt lncRNA derived from the chromosomal prt–pho1 locus in logarithmically growing vegetative cells is rapidly degraded by the nuclear exosome under the direction of DSR (determinant of selective removal) elements in the prt RNA (Lee et al. 2013; Shah et al. 2014; Chatterjee et al. 2016). However, increasing the gene dosage of the prt–pho1 locus (depicted in Fig. 3A) by placing it on a multicopy reporter plasmid in pho1Δ cells has permitted the analysis of internally terminated prt transcripts by northern blotting and the identification of two internal prt poly(A) sites, PAS and PAS2, by 3′-RACE (Sanchez et al. 2018a). The prt locus gives rise to three classes of poly(A)+ RNA: (i) a ∼2.5 kb RNA corresponding to a prt–pho1 read-through transcript, this being the lncRNA that interferes with pho1 mRNA synthesis; (ii) a ∼0.4 kb species, prt PAS, that corresponds to prt RNA that was cleaved and polyadenylated at the +351 PAS site; and (iii) a ∼0.6 kb species, prt PAS2, that corresponds to prt RNA that was cleaved and polyadenylated at the +589 PAS2 site (Fig. 3A). These three classes of transcript are seen in a northern blot of RNAs isolated from three independent cultures of duf89-WT pho1Δ cells bearing the prt–pho1 reporter plasmid (Fig. 3B, top left panel, lanes WT). We find that production of the prt–pho1 read-through transcript is strongly squelched in reporter-bearing duf89Δ cells, whereas the internally terminated transcripts are relatively spared (Fig. 3B, top left panel, lanes duf89Δ). This result is consistent with the idea that absence of Duf89 enhances the propensity of Pol2 to terminate prt transcription prior to traversal of the pho1 gene. Northern blotting with an mRNA-specific probe revealed two transcripts derived from the pho1 reporter in duf89-WT cells: a ∼1.6-kb pho1 mRNA and a longer prt–pho1 lncRNA (Fig. 3B, middle left panel, lanes WT). The duf89Δ mutation curtailed production of the interfering lncRNA and increased the level of the pho1 mRNA (Fig. 3B, middle left panel, lanes duf89Δ). The effects of duf89Δ on the prt and pho1 transcripts echo those seen for the asp1-H397A and rad24Δ mutations that elicit pho1 derepression (Sanchez et al. 2019; Garg et al. 2022).
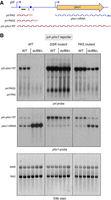
Derepression of pho1 expression by duf89Δ ensues from squelching the production of interfering prt lncRNA. (A) Schematic of the prt–pho1 locus in the reporter plasmid. Transcription start sites are indicated by bent blue arrows. Triangles denote internal poly(A) sites PAS and PAS2. DSR element clusters are indicated by small blue boxes. The gene-specific probe for prt (a 32P-labeled ssDNA complementary to the segment of the prt RNA from nucleotides +160 to +202) is denoted by a horizontal black bar. Three classes of poly(A)+ prt transcripts are depicted as red wavy lines below the prt–pho1 locus. (B) RNA was isolated from three independent cultures of duf89-WT pho1Δ or duf89Δ pho1Δ cells bearing the wild-type (WT) prt–pho1 reporter plasmid or mutant prt–pho1 plasmids in which the prt DSR clusters or PAS and PAS2 polyadenylation signals were altered by nucleobase substitutions. The RNAs were resolved by formaldehyde-agarose gel electrophoresis and stained with ethidium bromide to visualize 28S and 18S ribosomal RNAs (3485 and 1842 nucleotides, respectively) (bottom panel). The RNAs in the gel were transferred to a membrane and hybridized to the prt probe (top panel) and a pho1 mRNA probe (middle panel). Annealed probes were visualized by autoradiography. The various classes of prt and pho1 transcripts are indicated on the left.
Derepression of pho1 by duf89Δ depends on DSR elements in the prt lncRNA
The prt lncRNA contains two clusters of DSR elements (Fig. 3A, denoted by blue boxes), each composed of three DSR hexanucleotide motifs. The DSR clusters in the nascent lncRNA are binding sites for Mmi1 (Kilchert et al. 2015; Chatterjee et al. 2016). Previous studies underscored the role of the DSR clusters in prt-promoted pho1 repression, by surveying the effects of compound mutations introduced into each of the hexanucleotide motifs comprising the DSR sequences of the prt–pho1 reporter plasmid (Chatterjee et al. 2016; Sanchez et al. 2018a). Mutating the distal DSR cluster reduced Pho1 expression by fivefold; mutating the upstream DSR cluster reduced Pho1 expression by half (Chatterjee et al. 2016). Mutating both DSR clusters reduced Pho1 expression to 11% of the level achieved with the wild-type prt–pho1 reporter (Fig. 4). The hyperrepressive effect of the prt DSR mutations on Pho1 expression were shown previously to “win out” over the derepressive effects of CTD-S7A, asp1-H397A (increased IP8), erh1Δ, and rad24Δ, suggesting that DSRs are key to establish the precocious termination of prt lncRNA synthesis that underlies Pho1 derepression (Chatterjee et al. 2016; Sanchez et al. 2019; Schwer et al. 2020; Garg et al. 2022). Here, we found that the DSR mutations reduced Pho1 expression in duf89Δ cells to 4% of the wild-type DSR reporter control (Fig. 4). We conclude that duf89Δ derepression of pho1 is contingent on the DSR elements in the prt lncRNA.
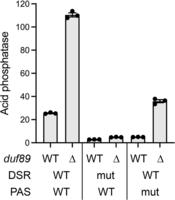
Contribution of DSR elements and poly(A) signals in the prt lncRNA to derepression of Pho1 acid phosphatase by duf89Δ. duf89-WT pho1Δ or duf89Δ pho1Δ cells bearing prt–pho1 reporter plasmids with wild-type or mutated (mut) DSR clusters and polyadenylation signals (PAS) as indicated were assayed for acid phosphatase activity.
A prt-probed northern blot of RNAs isolated from three independent cultures of duf89-WT pho1Δ and duf89Δ pho1Δ cells bearing the DSR mutant prt–pho1 reporter plasmid is shown in Figure 3B. Compared to the prt signal of wild-type cells wild-type reporter, we see that mutation of the DSRs stabilized the prt–pho1 read-through transcript and a short prt transcript running slightly ahead of prt PAS2 (observed previously in rad24Δ cells and presumed to reflect use of an alternative poly(A) site), thereby increasing their steady-state levels (Fig. 3B, top middle panel, lanes WT). The salient finding was that the ablation of the prt-pho1 read-through transcript by duf89Δ was prevented by DSR mutation, as was the duf89Δ-driven increase in the pho1 mRNA (Fig. 3B, top middle panel, lanes duf89Δ). The RNA analysis affirms that derepression of pho1 by duf89Δ requires the prt lncRNA DSRs. The level of the prt PAS transcript derived from the DSR mutant reporter plasmid was slightly higher, and the read-through transcript slightly lower, in duf89Δ versus duf89-WT cells (Fig. 3B, top middle panel), consistent with the idea that duf89Δ elicits precocious 3′-processing.
Effect of prt lncRNA PAS mutations on prt–pho1 reporter activity in duf89Δ cells
3′-processing of nascent prt lncRNA at PAS and PAS2 via the canonical poly(A) pathway will terminate prt transcription upstream of the Pho7 binding sites in the pho1 mRNA promoter and thus alleviate transcription interference. PAS and PAS2 utilization is a tunable influence on pho1 expression, insofar as: (i) simultaneous nucleobase substitution mutations of the prt PAS and PAS2 elements of the prt–pho1 reporter result in fivefold hyperrepression of Pho1 expression (Fig. 4); and (ii) dual PAS mutations attenuate the derepression of Pho1 expression elicited by CTD-S7A, seb1-G476S, and rad24Δ alleles (Sanchez et al. 2018a; Schwer et al. 2021; Garg et al. 2022). A prt-probed northern blot of RNAs from duf89-WT pho1Δ cells bearing the dual PAS mutant prt–pho1 reporter plasmid (Fig. 3B, top right panel) revealed an increase in the steady-state level of the prt–pho1 read-through transcript compared to the wild-type reporter control, albeit to a lesser extent than the increase seen in the DSR mutant reporter. The dual PAS mutations partially ameliorated the squelching of prt–pho1 read-through by duf89Δ, and partially restored pho1 mRNA expression (Fig. 3B, middle right panel) and Pho1 acid phosphatase activity (Fig. 4) vis-à-vis duf89Δ cells bearing the wild-type reporter. The persistence of short prt transcripts derived from the PAS mutant reporter underscores that there are additional poly(A) sites, yet unmapped, within the prt transcription unit that can come into play when the two known PAS elements are mutated.
duf89Δ derepression of Pho1 expression depends on IP8 synthesis by Asp1 kinase
Previous studies showed that derepression of Pho1 elicited by the Pol2 CTD-S7A mutation, the Seb1-G476S mutation, a deletion of Erh1, or a deletion of Rad24 depends on the activity of the kinase Asp1 (Sanchez et al. 2019; Schwer et al. 2020, 2021; Garg et al. 2022). To see if IP8 status affects Pho1 derepression by duf89Δ, we crossed duf89Δ to a kinase-dead asp1-D333A strain. As noted previously (Sanchez et al. 2019), asp1-D333A cells evinced a sixfold hyperrepression of acid phosphatase activity (Fig. 1B). We obtained viable duf89Δ asp1-D333A haploids after mating and sporulation. Growth of the double-mutant on YES agar was like that of asp1-D333A; both strains formed smaller colonies than the wild-type control at 20°C and 37°C (Fig. 1A). The instructive finding was that the derepression of Pho1 activity by duf89Δ was eliminated in the asp1-D333A background (Fig. 1B), signifying a requirement for inositol 1-pyrophosphate synthesis, presumably via IP8 action as an agonist of 3′-processing/termination during prt transcription (Sanchez et al. 2019).
duf89Δ derepression of Pho1 depends on the Pol2 CTD Thr4 mark
Changing all Pol2 CTD Thr4 residues to alanine leads to a sixfold hyperrepression of Pho1 expression in phosphate-replete rpb1-CTD-T4A cells (Fig. 1B). Previous epistasis analyses showed that the CTD-T4A allele erases the derepression of Pho1 elicited by an active site mutation of the Asp1 pyrophosphatase (asp1-H397A) or a null mutation of the Aps1 pyrophosphatase (aps1Δ) (Sanchez et al. 2019). Here, we constructed a duf89Δ CTD-T4A double mutant, which grew as well on YES agar as the CTD-T4A single mutant (Fig. 1A). The salient finding was that the CTD-T4A allele canceled the derepression of Pho1 by duf89Δ (Fig. 1B).
duf89Δ is synthetically lethal with rpb1-CTD-S7A
Changing all Pol2 CTD Ser7 residues to alanine derepresses pho1. CTD-S7A is synthetically lethal with the pyrophosphatase-dead asp1-H397A allele that elicits precocious prt termination (Sanchez et al. 2019). Here we found that CTD-S7A was lethal in combination with duf89Δ (Fig. 1A), as judged by a failure to recover viable duf89Δ rpb1-S7A haploids after mating the respective single mutants, sporulating the heterozygous diploids, and screening a large population of random spores for the antibiotic-resistance markers linked to the duf89Δ and CTD-S7A loci. We hypothesize that Duf89 and the CTD Ser7 mark play genetically redundant roles in evading precocious 3′-processing and transcription termination.
Duf89 genetic interactions with fission yeast inositol pyrophosphatases
The fission yeast proteome includes three pyrophosphatase enzymes that convert IP8 to IP7 (Fig. 5C). The C-terminal pyrophosphatase domain of Asp1, which belongs to the histidine acid phosphatase family, hydrolyzes pyrophosphate at the 1 position of the inositol ring to generate 5-IP7 (Pascual-Ortiz et al. 2018; Dollins et al. 2020; Benjamin et al. 2022). Siw14, a member of the cysteinyl phosphatase family, cleaves the pyrophosphate at the 5 position to form 1-IP7 (Wang et al. 2018). Aps1, a Nudix family enzyme, can hydrolyze the β-phosphate at either the 1 or 5 pyrophosphate positions (Safrany et al. 1999; Kilari et al. 2013; Zong et al. 2021). None of the three pyrophosphatase activities is essential per se for fission yeast vegetative growth, that is, the pyrophosphatase-defective asp1-H397A strain and the aps1Δ and siw14Δ null strains grow well on YES agar at 20°C to 37°C (Figs. 5A, 12A). However, asp1-H397A is synthetically lethal with aps1Δ (Sanchez et al. 2019), suggesting that simultaneous ablation of these two pyrophosphatases results in accumulation of toxic levels of IP8.
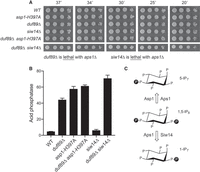
Duf89 genetic interactions with fission yeast inositol pyrophosphatases. (A) S. pombe wild-type, single-mutant, and double-mutant strains with the indicated duf89, asp1, and siw14 alleles were spot tested for growth at the temperatures specified. aps1Δ was synthetically lethal with duf89Δ and siw14Δ. (B) The indicated fission yeast strains were assayed for Pho1 acid phosphatase activity. (C) Inositol pyrophosphatases convert 1,5-IP8 to either 5-IP7 or 1-IP7 as shown.
Epistasis of Duf89 with the three pyrophosphatases with respect to cell growth and Pho1 expression was queried by attempting to construct all double mutant strains via mating and sporulation. We thereby found (by random spore analysis; see Materials and Methods) that duf89Δ asp1-H397A and duf89Δ siw14Δ strains were viable and grew well on YES agar (Fig. 5A). In contrast, the duf89Δ and aps1Δ alleles were synthetically lethal; to wit, (i) we were unable to obtain viable double mutants after screening a large population of haploid progeny of the genetic cross; and (ii) wild-type progeny and the differentially marked duf89Δ (on chromosome III) and aps1Δ (on chromosome I) single mutants were recovered at the expected frequencies. This result signifies that the Duf89 and the Aps1 pyrophosphatase have essential but redundant functions in fission yeast, and it raises the possibility that Duf89 might participate in inositol pyrophosphate metabolism or inositol pyrophosphate signaling. We also found that siw14Δ and aps1Δ were synthetically lethal (Fig. 5A).
Analysis of Pho1 expression in the viable single and double mutants grown under phosphate-replete conditions is shown in Figure 5B. Whereas duf89Δ and asp1-H397A single mutations derepressed Pho1 by ninefold and 12-fold, respectively, compared to the wild-type control, siw14Δ had no effect per se on Pho1 expression. The Pho1 activity of the duf89Δ asp1-H397A double mutant was virtually the same as that of the asp1-H397A single mutant, that is, there was no additive effect of duf89Δ on asp1-H397A. The duf89Δ siw14Δ strain exhibited a higher level of Pho1 derepression (15-fold vs. wild-type) than duf89Δ alone (Fig. 5B).
Transcriptome profiling of duf89Δ cells
We performed RNA-seq on poly(A)+ RNA isolated from duf89Δ cells and from the parental wild-type strain. cDNAs obtained from three biological replicates (using RNA from cells grown to mid-log phase in YES medium at 30°C) were sequenced for each strain. In the data sets, 97%–98% of the reads were mapped to unique genomic loci (Supplemental Fig. S1). Read densities (RPKM) for individual genes were highly reproducible between biological replicates (Pearson coefficients of 0.977 to 0.987; Supplemental Fig. S2). A cutoff of plus or minus twofold change in normalized transcript read level and an adjusted P-value of ≤0.05 were the criteria applied to derive an initial list of differentially expressed annotated loci in the duf89Δ mutant versus the wild-type control. We then focused on differentially expressed genes with average normalized read counts ≥100 in either the duf89Δ or wild-type strains to eliminate transcripts that were expressed at very low levels in vegetative cells. We thereby identified a set of 15 annotated protein-coding genes that were up-regulated by these criteria in duf89Δ cells, which includes all three phosphate homeostasis genes: tgp1 (up 7.3-fold), pho1 (up 3.3-fold), and pho84 (up 2.5-fold) (Supplemental Fig. S3). The ecl3 gene, up-regulated by fourfold, is located on chromosome II, adjacent to and in opposite orientation to the prt2 lncRNA gene of the phosphate-regulated prt2–pho84–prt–pho1 gene cluster; ecl3 is coregulated with the PHO genes in several mutant genetic backgrounds (Sanchez et al. 2019). One other gene involved in phosphate acquisition was also up-regulated: SPBPB2B2.06c encoding an extracellular 5′ nucleotidase (up fivefold). Most pertinent is that 9/15 (60%) of the mRNAs that were overexpressed in duf89Δ cells were also included in the set of 19 mRNAs found to be overexpressed in aps1Δ cells that lack the Nudix-family pyrophosphatase Aps1 (Sanchez et al. 2019), that is, the very genetic background in which duf89Δ is synthetically lethal. The RNA-seq experiment identified 23 protein-coding genes that were down-regulated by greater than or equal to twofold in duf89Δ cells (Supplemental Fig. S4), 14 of which (61%) were also down-regulated in aps1Δ cells (Sanchez et al. 2019). The overlapping transcriptional signatures hint that Duf89 might be a functional analog of Aps1.
Derepression of pho1 expression by duf89Δ depends on CPF subunits, Rhn1, and Pin1
The fission yeast cleavage and polyadenylation factor (CPF) is a 13-subunit protein assembly responsible for the 3′ processing of nascent Pol2 transcripts that precedes Pol2 transcription termination (Vanoosthuyse et al. 2014). Five of the CPF subunits (Ctf1, Ssu72, Dis2, Ppn1, and Swd22) are dispensable for growth. Dis2 and Ssu72 are phosphoprotein phosphatase enzymes. Rhn1 is an inessential CTD-binding Pol2 termination factor that recognizes the Thr4-PO4 CTD mark (Jasnovidova et al. 2017). Pin1 is a peptidyl prolyl isomerase that abets the function of Ssu72, a cis-proline-dependent CTD phosphatase. Loss-of-function mutants of Pin1, Rhn1, or any of the five inessential CPF subunits result in hyperrepression of pho1 RNA expression and Pho1 acid phosphatase activity in phosphate-replete cells (Sanchez et al. 2018a, 2020). Moreover, CPF/Rhn1/Pin1 mutations erase or attenuate the derepression of pho1 by genetic maneuvers that promote precocious prt lncRNA 3′ processing/termination, for example, by pyrophosphatase mutations asp1-H397A and aps1Δ, by Pol2 CTD-S7A, by erh1Δ, and by rad24Δ (Sanchez et al. 2018a, 2019, 2020; Schwer et al. 2020; Garg et al. 2022). If duf89Δ were exerting its derepressive effects on the pho1 via precocious lncRNA termination, then we would expect derepression of pho1 in duf89Δ cells to be reversed by CPF/Rhn1/Pin1 mutations.
To address this issue, we performed pairwise mating of the duf89Δ strain with dis2Δ, ctf1Δ, ssu72-C13S (a phosphatase-dead allele), ppn1Δ, swd22Δ, rhn1Δ, and pin1Δ strains, which yielded viable haploid double mutants in every case. The duf89Δ CPF/rhn1/pin1 strains grew well on YES agar at all temperatures (Fig. 6A). Indeed, the temperature sensitivity of rhn1Δ at 37°C (Sanchez et al. 2018a) was alleviated by duf89Δ (Fig. 6A). The duf89Δ CPF/rhn1/pin1 double mutants were assayed for Pho1 acid phosphatase activity during growth in phosphate-replete medium. The derepression of Pho1 by duf89Δ was effaced in rhn1Δ, ssu72-C13S, ppn1Δ, swd22Δ, and pin1Δ cells and was attenuated in ctf1Δ and dis2Δ cells (Fig. 6B). Thus, the increase in Pho1 expression by duf89Δ requires CPF subunits, Pin1, and Rhn1, consistent with the precocious lncRNA termination model.
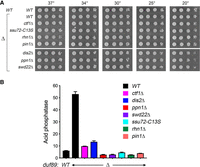
Derepression of Pho1 expression by duf89Δ depends on CPF subunits, Rhn1, and Pin1. (A) S. pombe duf89+ (WT) cells and cells bearing the duf89Δ (Δ) allele in combination with CPF subunit, Rhn1, and Pin1 mutations were spot tested for growth at the temperatures specified. (B) The indicated strains were grown in liquid culture at 30°C and assayed for acid phosphatase activity.
Lethality of duf89Δ aps1Δ is rescued by CPF, Pin1, Rhn1, Asp1 kinase, and CTD-T4A mutations
The key question raised by the results of the preceding two sections is whether the lethality of the duf89Δ aps1Δ strain arises from unconstrained precocious transcription termination. If so, then it might be expected that the lethality would be ameliorated by mutations in the 3′-processing/termination machinery. To test this idea, we crossed our duf89Δ CPF/pin1/rhn1 double-mutants to aps1Δ then sporulated the resulting diploids and screened random spores for each of the differentially marked loci of interest. In this way, we recovered viable duf89Δ aps1Δ dis2Δ, duf89Δ aps1Δ ppn1Δ, duf89Δ aps1Δ swd22Δ, duf89Δ aps1Δ ctf1Δ, duf89Δ aps1Δ ssu72-C13S, and duf89Δ aps1Δ pin1Δ haploid strains that grew on YES agar at all temperatures (Fig. 7A). The viable duf89Δ aps1Δ rhn1Δ triple mutant grew well at 30°C–37°C but displayed a cold-sensitive growth defect at 20°C–25°C (Fig. 7A). These results suggest that the synthetic lethality of duf89Δ aps1Δ is a consequence of IP8-driven precocious termination that depends on CPF, Pin1, and Rhn1. The duf89Δ aps1Δ dis2Δ and duf89Δ aps1Δ ctf1Δ strains expressed higher levels of Pho1 than the duf89Δ and aps1Δ single mutants (Fig. 7B). In the five other triple-mutants, Pho1 expression was intermediate between that of wild-type and aps1Δ cells, with rhn1Δ being the most effective at reversing the derepression of Pho1 elicited by duf89Δ aps1Δ (Fig. 7B).
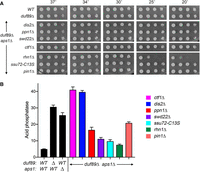
Lethality of duf89Δ aps1Δ is rescued by CPF, Pin1, and Rhn1 mutations. (A) S. pombe cells with indicated genotypes were spot tested for growth at the temperatures specified. (B) The indicated strains were grown in liquid culture at 30°C and assayed for acid phosphatase activity.
If, as suggested, the synthetic lethality of duf89Δ aps1Δ is a function of IP8-dependent precocious 3′-processing/termination of one or more essential fission yeast genes, then the synthetic lethality should be suppressed by the kinase-dead asp1-D333A allele. Indeed, after crossing aps1Δ asp1-D333A and duf89Δ strains and random spore analysis, we obtained viable duf89Δ aps1Δ asp1-D333A cells that grew as well as wild-type on YES agar (Fig. 8A). The hyperrepressed Pho1 expression observed in the aps1Δ asp1-D333A double mutant was maintained in the duf89Δ aps1Δ asp1-D333A triple-mutant strain (Fig. 8B).
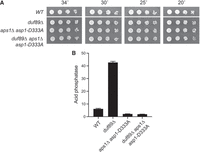
Lethality of duf89Δ aps1Δ is suppressed by the kinase-dead asp1-D333A allele. (A) S. pombe wild-type, single-mutant, double-mutant, and triple-mutant strains with the indicated duf89, asp1, and aps1 alleles were spot tested for growth at the temperatures specified. (B) The indicated strains were grown in liquid culture at 30°C and assayed for acid phosphatase activity.
Having found that CTD-T4A erases Pho1 derepression in duf89Δ cells (Fig. 1B), it was of interest to test if CTD-T4A (which negatively affects 3′-processing/termination) would suppress the lethality of duf89Δ aps1Δ. After crossing duf89Δ CTD-T4A and aps1Δ strains and screening by random spore analysis, we obtained viable triple mutant duf89Δ aps1Δ CTD-T4A cells that grew as well as the duf89Δ CTD-T4A strain on YES agar at 25°C to 34°C and slightly better than duf89Δ CTD-T4A at 20°C (Supplemental Fig. S5A). Pho1 expression in duf89Δ aps1Δ CTD-T4A cells was intermediate between that of the duf89Δ and duf89Δ CTD-T4A strains (Supplemental Fig. S5B).
Lethality of duf89Δ CTD-S7A is rescued by CPF, Pin1, and Rhn1 mutations
Crosses of duf89Δ to CTD-S7A CPF/pin1/rhn1 double mutants (Sanchez et al. 2018a) yielded viable triple mutant haploids duf89Δ CTD-S7A dis2Δ, duf89Δ CTD-S7A ppn1Δ, duf89Δ CTD-S7A swd22Δ, duf89Δ CTD-S7A ctf1Δ, duf89Δ CTD-S7A ssu72-C13S, duf89Δ CTD-S7A pin1Δ, and duf89Δ CTD-S7A rhn1Δ that grew on YES agar at 30°C–34°C and, in some cases, displayed ts or cs growth phenotypes (Fig. 9A). We surmise that the synthetic lethality of duf89Δ rpb1-CTD-S7A is a consequence of precocious 3′-processing/termination. Pho1 acid phosphatase activities in the duf89Δ CTD-S7A ctf1Δ and duf89Δ CTD-S7A dis2Δ mutants were similar to those of duf89Δ and CTD-S7A single mutants, respectively (Fig. 9B). In contrast, introduction of the ppn1Δ, swd22Δ, ssu72-C13S, and pin1Δ alleles into the duf89Δ CTD-S7A background reduced Pho1 expression to less than the wild-type level (Fig. 9B).
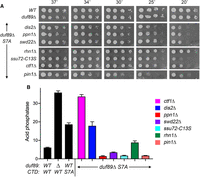
Lethality of duf89Δ CTD-S7A is rescued by CPF, Pin1, and Rhn1 mutations. (A) S. pombe cells with indicated genotypes were spot tested for growth at the temperatures specified. (B) The indicated strains were grown in liquid culture at 30°C and assayed for acid phosphatase activity.
Lethality of dis2Δ ssu72-C13S is rescued by duf89Δ
The synthetic lethality accompanying simultaneous inactivation of both protein phosphatase subunits of the CPF complex (these being dis2Δ and ssu72-C13S) is posited to be the consequence of a 3′-processing/termination defect impacting the expression of essential S. pombe genes. We reported previously that the lethality of the dis2Δ ssu72-C13S double-mutant was ameliorated by a third mutation that exerts an opposite effect, that is, the asp1-H397A allele that promotes precocious termination of the lncRNAs that control phosphate homeostasis (Sanchez et al. 2019). Here we queried whether duf89Δ might also suppress the lethality of dis2Δ ssu72-C13S. After crossing duf89Δ dis2Δ and ssu72-C13S strains and screening by random spore analysis, we recovered viable duf89Δ dis2Δ ssu72-C13S triple mutant haploids. The duf89Δ dis2Δ ssu72-C13S triple mutant strain grew less well than wild-type on YES agar at 30°C but similarly to the ssu72-C13S single mutant (as gauged by colony size) (Fig. 10A). With respect to Pho1 expression, the duf89Δ dis2Δ ssu72-C13S triple mutant phenocopied the duf89Δ ssu72-C13S double mutant in erasing the derepression of Pho1 caused by duf89Δ (Fig. 10B).
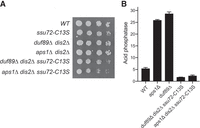
Lethality of dis2Δ ssu72-C13S is rescued by duf89Δ and aps1Δ. (A) S. pombe cells with indicated genotypes were spot tested for growth at 30°C. (B) The indicated strains were grown in liquid culture at 30°C and assayed for acid phosphatase activity.
In light of the genetic evidence that Duf89 overlaps functionally with the Nudix pyrophosphatase Aps1, we queried whether the lethality of dis2Δ ssu72-C13S might be similarly overcome by deleting aps1. After crossing aps1Δ dis2Δ and ssu72-C13S strains (Sanchez et al. 2019) we obtained viable aps1Δ dis2Δ ssu72-C13S cells that grew slowly on YES agar at 30°C (Fig. 10A) and were hyperrepressed with respect to Pho1 expression (Fig. 10B).
duf89Δ derepression of Pho1 depends on Spx1
Deleting the inositol pyrophosphate-binding RING-domain ubiquitin ligase Spx1 (or alanine mutations of its inositol pyrophosphate-binding site or zinc-binding cysteines) results in hyperrepression of Pho1 in phosphate-replete cells (Schwer et al. 2022). Moreover, the spx1Δ and the inositol pyrophosphate-binding or Zn-binding mutations erase the derepression of Pho1 elicited by asp1-H397A and suppress the lethality of asp1 alleles STF6 and STF9 (Schwer et al. 2022). Spx1 is proposed to be a mediator of the inositol pyrophosphate signal that leads to precocious transcription termination. Here we constructed a duf89Δ spx1Δ double mutant, which grew as well on YES agar as the duf89Δ single mutant (Supplemental Fig. S6A). spx1Δ erased the derepression of Pho1 by duf89Δ (Supplemental Fig. S6B).
duf89Δ and rad24Δ additively derepress Pho1 expression
Deleting the 14-3-3 protein Rad24 results in a 21-fold derepression of Pho1 expression under phosphate-replete conditions (Fig. 11B); this is the most potent Pho1 derepressive single mutation identified to date. Mating differentially marked rad24Δ and duf89Δ strains yielded rad24Δ duf89Δ double mutant haploids that grew as well on YES agar at 30°C and 34°C as the rad24Δ single mutant (Fig. 11A). duf89Δ exerted an additive effect on Pho1 expression in the rad24Δ background, whereby acid phosphatase activity was derepressed by 35-fold vis-à-vis wild-type cells (Fig. 11B).
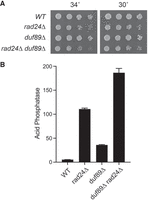
duf89Δ and rad24Δ additively derepress Pho1 expression. (A) Wild-type, duf89Δ, rad24Δ, and rad24Δ duf89Δ cells were spot tested for growth at 30°C and 34°C. (B) The indicated strains were grown in liquid culture at 30°C and assayed for acid phosphatase activity.
Duf89 mutation D252A does not result in Pho1 derepression
A pertinent question is whether the catalytic activity of Duf89 is required for its function in maintaining the repressed state of the PHO regulon in phosphate-replete cells. To address this issue, we replaced the duf89+ chromosomal allele with the phosphatase-dead mutant D252A, flanked by a downstream hygromycin-resistance cassette. We had shown previously that changing Asp252 to Ala abolished the capacity of recombinant Duf89 protein to hydrolyze p-nitrophenylphosphate and inorganic pyrophosphate (Sanchez et al. 2022). Asp252 is the nucleophile that attacks the phospho-substrate to form an enzyme-(aspartyl)–phosphate intermediate in the two-step phosphohydrolase reaction (Sanchez et al. 2022). As a control, the hygromycin-resistance cassette was introduced downstream from the wild-type duf89 gene. The marked wild-type duf89 and D252A mutant strains grew equally well on YES agar (Fig. 12A). Counter to our expectations, the D252A mutation did not derepress Pho1 acid phosphatase expression à la duf89Δ (Fig. 12B), signifying that Duf89 protein, but not its phosphatase activity, is central to proper PHO homeostasis under phosphate-rich conditions. In the same vein, the duf89-D252A allele differed from duf89Δ in that it was not synthetically lethal with aps1Δ (Fig. 12A). The Pho1 expression was derepressed to the same extent in aps1Δ duf89-WT and aps1Δ duf89-D252A cells (Fig. 12B). The Asp290 side chain of Duf89 coordinates the essential metal cofactor for the phosphohydrolase reaction; mutating Asp290 to alanine abolished Duf89's p-nitrophenylphosphatase and inorganic pyrophosphatase activities (Sanchez et al. 2022). We found that the phosphatase-dead duf89-D290A allele did not synergize with aps1Δ, that is, as was the case for duf89-D252A, a genetic cross of single mutants yielded a viable duf89-D290A aps1Δ double-mutant strain that grew as well as wild-type on YES agar at all temperatures (not shown).
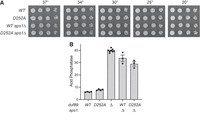
Duf89 mutation D252A does not result in Pho1 derepression or synthetic lethality with aps1Δ. (A) Fission yeast strains bearing marked duf89-WT or duf89-D252A alleles were spot tested for growth on YES agar at the temperatures specified. (B) duf89-WT, duf89Δ, and duf89-D252A strains were assayed for acid phosphatase activity.
Increasing duf89 gene dosage hyperrepresses Pho1 expression
Given that ablating duf89 derepresses Pho1 expression, it was of interest to test whether increasing duf89 gene dosage might have the opposite effect. Accordingly, we introduced into wild-type fission yeast a multicopy pTIN plasmid bearing duf89, either wild-type or the D252A mutant. The pTIN expression vector (Garg 2020) places the Duf89 open reading frame under the transcriptional control of the tgp1 promoter, which is situated adjacent to the transcription unit specifying the nc-tgp1 lncRNA driven by the thiamine-repressible nmt1 promoter. In the absence of thiamine, lncRNA transcription interferes with firing of the tgp1 promoter. In the presence of thiamine, lncRNA synthesis is turned off and expression of the downstream mRNA—encoding Duf89 in this case—is turned on (Garg 2020). Comparing Pho1 acid phosphatase activity of thiamine-replete cells bearing the pTIN-Duf89-WT or pTIN-Duf89-D252A plasmid (cell-normalized activity values of 1.06 and 1.11, respectively) versus cells bearing the empty pTIN vector (activity 4.86) showed that increased gene dosage hyperrepressed Pho1 expression (Fig. 13). Moreover, the derepression of Pho1 in duf89Δ cells bearing the empty pTIN vector (activity 52.9) was erased completely by the pTIN-Duf89-WT and pTIN-Duf89-D252A plasmids (activity values of 1.09 and 1.01, respectively) (Fig. 13). Finally, the derepression of Pho1 in aps1Δ cells bearing the empty pTIN vector (activity 43.9) was partially blunted by the pTIN-Duf89-WT and pTIN-Duf89-D252A plasmids (activity values of 28.7 and 31.5, respectively, which, by pairwise t-tests versus empty vector were significantly lower; P values of 0.019 and 0.027, respectively) (Fig. 13).
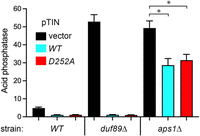
Increased duf89-WT or duf89-D252A gene dosage hyperrepresses Pho1 expression. Single colonies (≥20) of wild-type, duf89Δ, and aps1Δ cells bearing pTIN plasmids encoding wild-type or mutant Duf89, or the empty pTIN vector, respectively, were pooled and grown in Leu- ePMG with 15 µM thiamine. Aliquots of exponentially growing cultures were assayed for acid phosphatase activity. The data are averages (±SEM) of three independent biological replicates. Pairwise t-test showed significant differences between aps1Δ cells bearing the empty pTIN plasmid and pTIN-Duf89-WT or pTIN-Duf89-D252A (P-values of 0.019 and 0.027, respectively, denoted by asterisks).
Effect of Aps1 pyrophosphatase active site mutations
The results in the preceding section raised the question of whether the pyrophosphatase activity of Aps1 is required for its function in PHO regulation. To address this issue, we replaced the wild-type aps1+ gene with mutated versions—E89A, E93A, or E89A-E93A—in which one or both of the essential metal-binding glutamates of the Aps1 Nudix motif (Yang et al. 1999) were changed to alanine. The aps1-Ala strains grew as well as wild-type or aps1Δ cells on YES agar at 30°C (Supplemental Fig. S7A) and the active site mutations elicited derepression of Pho1 to the same extent as did aps1Δ (Supplemental Fig. S7B). We conclude that Aps1 pyrophosphatase activity is required for repression of Pho1 in phosphate-replete cells.
To see if Aps1 pyrophosphatase activity is pertinent to the synthetic lethality of aps1Δ with duf89Δ, asp1-H397A, and CTD-S7A, we crossed the aps1-E89A and aps1-E93A mutants to differentially marked duf89Δ, asp1-H397A, and CTD-S7A strains and screened for double-mutants by random spore analysis. The aps1-Ala active site mutations phenocopied aps1Δ with respect to synthetic lethality with asp1-H397A, consistent with the idea that inactivation of these two pyrophosphatases at once results in the accumulation of toxic levels of IP8 (Sanchez et al. 2019). The noteworthy findings were that aps1-E89A and aps1-E93A were viable, albeit quite slow growing, in combination with duf89Δ and CTD-S7A (Supplemental Fig. S8), signifying that these pyrophosphatase-dead versions of Aps1 retain some level of biological activity in a subset of genetic backgrounds in which the absence of Aps1 is lethal.
DISCUSSION
The results herein provide genetic and biochemical evidence that fission yeast Duf89 is an antagonist of precocious 3′-processing and transcription termination. Since the original report by the Wykoff laboratory that phosphate-replete duf89Δ cells overexpress Pho1 acid phosphatase (Henry et al. 2011), it has been established that: (i) the fission yeast PHO regulon genes are subject to lncRNA-mediated transcriptional interference; and (ii) the interference phenomenon is tunable by genetic manipulations that enhance or diminish precocious lncRNA termination in response to cis-acting PAS and DSR elements within the lncRNAs. duf89Δ joins a roster of mutant alleles that elicit Pho1 derepression by increasing the probability of precocious lncRNA termination (Fig. 14A), from which it is inferred that Duf89—like Rad24, Erh1, the Ser5-Pro6-Ser7 “word” in the CTD code, and the catabolism of IP8 by Asp1 and Aps1 pyrophosphatases—functions to dampen precocious termination. duf89Δ derepression of Pho1 correlates with squelching the production of full-length prt lncRNA and is erased or attenuated by: (i) DSR/PAS mutations in prt; (ii) loss-of-function mutations in components of the 3′-processing and termination machinery; (iii) elimination of the CTD Thr4-PO4 mark: (iv) interdicting CTD prolyl isomerization by Pin1; (v) inactivating the Asp1 kinase that synthesizes IP8; and (vi) loss of the putative IP8 sensor Spx1. Each of the aforementioned genetic maneuvers is pho1 hyperrepressive on its own, as is overexpression of duf89 via increased gene dosage (Fig. 14B). That duf89Δ is synthetically lethal with pho1-derepressive mutations CTD-S7A and aps1Δ—and that such synthetic lethality is rescued by CTD-T4A, CPF/Rhn1/Pin1 mutations, and spx1Δ—implicates Duf89 more broadly as a collaborator in cotranscriptional regulation of essential fission yeast genes. This point is underscored by our finding that duf89Δ rescues the synthetic lethality of the dis2Δ ssu72-C13S strain, that is, the “yin” of precocious 3′-processing/termination caused by duf89Δ compensates for the “yang” of defective processing/termination when the two protein phosphatase subunits of CPF are inactivated.
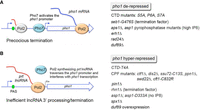
lncRNA control of Pho1 expression is tunable by genetic maneuvers affecting 3′-processing/termination. prt lncRNA synthesis represses pho1 mRNA transcription in phosphate-replete conditions. States of pho1 derepression (A) and hyperrepression (B) at the prt-pho1 locus are achieved by changes in prt lncRNA 3′-processing/termination efficiency. The genetic perturbations that elicit these effects are indicated on the right.
Synthetic lethality of duf89Δ with aps1Δ was especially provocative given that Duf89 and Aps1 are both phosphoesterase enzymes and that duf89Δ and aps1Δ single deletions evoke highly concordant perturbations of their mRNA transcriptomes. Aps1 orthologs are Nudix hydrolases implicated in the catabolism of inositol pyrophosphates and inorganic polyphosphates (Lonetti et al. 2011; Márquez-Moñino et al. 2021; Zong et al. 2021). Duf89 is a cobalt/nickel-dependent phosphomonoesterase and inorganic pyrophosphatase that acts via a covalent enzyme–aspartyl-phosphate intermediate (Sanchez et al. 2022). Although the physiological substrate(s) for Duf89 are not known, the genetics initially pointed toward inositol pyrophosphates and a scenario in which simultaneous inactivation of two pyrophosphatases (Aps1 and putatively Duf89) results in IP8 toxicity, akin to what happens when Aps1 is deleted and the pyrophosphatase domain of Asp1 is mutated (Sanchez et al. 2019). However, this model is vitiated by our finding that the duf89-D252A mutation, which abolishes Duf89 phosphohydrolase activity (Sanchez et al. 2022), in no way phenocopies duf89Δ, either with respect to derepression of Pho1 expression or synthetic lethality with aps1Δ. Thus, the duf89Δ phenotype is a consequence of Duf89 protein absence, rather than absence of Duf89 catalysis. This extends to the ability of Duf89 overexpression, either wild-type or D252A, to curb Pho1 expression. In this vein, it is noteworthy that we did not detect Duf89 hydrolysis of IP8 in vitro when assayed under conditions permissive for hydrolysis of inorganic pyrophosphate.
How might Duf89 function noncatalytically to antagonize precocious 3′-processing and termination? It is conceivable that Duf89 can bind and sequester IP8, without needing to hydrolyze it, and thereby limit IP8 availability for its action as an agonist of precocious termination. Alternatively, binding of some other phosphometabolite to Duf89 might enable its function as a 3′-processing/termination antagonist. Or, even more simply, Duf89 protein is a 3′-processing/termination antagonist independent of phospho-ligand binding, for example, via a putative physical interaction of Duf89 with some component(s) of the transcription and processing machinery.
MATERIALS AND METHODS
Deletion of duf89 and siw14
PCR amplification and standard cloning methods were used to construct plasmids in which the duf89 gene (SPCC1393.13) from nucleotides +5 to +1412 (relative to the translational start codon +1) and the siw14 gene (SPBC17A3.03c) from +5 to +1107 were replaced by a hygMX antibiotic resistance cassette. The duf89 and siw14 disruption cassettes were excised from the plasmid and transfected into diploid S. pombe cells. Hygromycin-resistant transformants were selected and analyzed by Southern blotting to confirm correct integration at one of the duf89 loci (thereby deleting amino acids 2–435 of the 442-aa Duf89 protein) or one of the siw14 loci (deleting amino acids 2–287 of the 287-aa Siw14 protein). Hygromycin-resistant duf89Δ and siw14Δ haploids were isolated upon sporulation of the heterozygous diploids. Nourseothricin-resistant and G418-resistant duf89Δ and siw14Δ strains were generated by marker switching (Hentges et al. 2005).
Tests of mutational synergies
duf89Δ haploids were mixed on malt agar with haploids of the opposite mating type bearing differentially marked mutations in genes involved in inositol pyrophosphate metabolism (asp1, aps1, siw14) and RNA 3′-processing and Pol2 transcription termination (ctf1, dis2, ppn1, swd22, ssu72, rhn1), CTD prolyl isomerization (pin1), or pan-heptad mutations in the Pol2 CTD (T4A, S7A) to allow mating and sporulation. The mixture was then subjected to random spore analysis. Spores (∼1000) were plated on YES agar and on medium selective for the marked mutant alleles and the plates were incubated at 30°C. At least 500 viable drug-resistant progeny were screened by replica-plating for the presence of the second drug resistance marker gene, or by sequentially replica-plating from YES to different drug-selective media. Wild-type (unmarked) and the differentially marked single mutant alleles were recovered at the expected frequencies. A finding that no haploids with both marker genes were recovered after 6 to 8 d of incubation at 30°C was taken to indicate synthetic lethality. Growth phenotypes of viable double-mutant strains were assessed in parallel with the individual single mutants and wild-type cells at different temperatures (20°C to 37°C) by spotting as follows. Fission yeast cultures were grown in YES liquid medium at 30°C until A600 reached 0.6–0.9. The cultures were adjusted to a final A600 of 0.1, and 3 µL aliquots of serial fivefold dilutions were spotted on YES agar. The plates were photographed after incubation for 2 d at 34°C, 2.5 d at 30°C and 37°C, 4 d at 25°C, and 6 d at 20°C.
Cell-surface acid phosphatase activity
Cells were grown at 30°C in YES medium. Aliquots of exponentially growing cultures were harvested, washed, and resuspended in water. To quantify acid phosphatase activity, reaction mixtures (200 µL) containing 100 mM sodium acetate (pH 4.2), 10 mM p-nitrophenylphosphate, and cells (ranging from 0.01 to 0.1 A600 units) were incubated for 5 min at 30°C. The reactions were quenched by addition of 1 mL of 1 M sodium carbonate, the cells were removed by centrifugation, and the absorbance of the supernatant at 410 nm was measured. Acid phosphatase activity is expressed as the ratio of A410 (p-nitrophenol production) to A600 (cells). The data are averages (±SEM) of at least three assays using cells from three independent cultures.
Northern blot analyses
Total RNA was extracted via the hot phenol method (Herrick et al. 1990) from 6 A600 units of yeast cells that had been grown exponentially in YES + G418 (for selection of kanMX plasmids) to A600 of 0.3 to 0.5 at 30°C. Aliquots (10 µg) of total RNA were resolved by electrophoresis through a 1.2% agarose/formaldehyde gel. After photography under UV light to visualize ethidium bromide-stained rRNAs and tRNAs, the gel contents were transferred to a Hybond-XL membrane (GE Healthcare). Hybridization was performed with a 5′ 32P-labeled ssDNA complementary to nucleotides 160–202 or 84–115 downstream from the prt or pho1 transcription start sites, respectively, using a commercial hybridization buffer (Invitrogen ULTRAhyb-Oligo). Hybridized probes were visualized by autoradiography.
Transcriptome profiling by RNA-seq
RNA was isolated from S. pombe wild-type and duf89Δ cells that were grown in liquid YES medium at 30°C to an A600 of 0.5 to 0.6. Cells were harvested by centrifugation and total RNA was extracted via the hot phenol method. The integrity of total RNA was gauged with an Agilent Technologies 2100 Bioanalyzer. The Illumina TruSeq stranded mRNA sample preparation kit was used to purify poly(A)+ RNA from 500 ng of total RNA and to carry out the subsequent steps of poly(A)+ RNA fragmentation, strand-specific cDNA synthesis, indexing, and amplification. Indexed libraries were normalized and pooled for paired-end sequencing performed by using an Illumina HiSeq 4000 system. FASTQ files bearing paired-end reads of length 51 bases were mapped to the S. pombe genome (ASM294v2.28) using HISAT2-2.1.0 with default parameters (Kim et al. 2015). The resulting SAM files were converted to BAM files using Samtools (Li et al. 2009). Count files for individual replicates were generated with HTSeq-0.10.0 (Anders et al. 2015) using exon annotations from Pombase (GFF annotations, genome-version ASM294v2; source “ensembl”). RPKM analysis and pairwise correlations (Supplemental Figs. S1, S2) were performed as described previously (Schwer et al. 2014). Differential gene expression and fold change analysis was performed in DESeq2 (Love et al. 2014). Cut-off for further evaluation was set for genes that had an adjusted P-value (Benjamini–Hochberg corrected) of ≤0.05 and were up or down by at least twofold in duf89Δ versus wild-type. Genes were further filtered on the following criteria: (i) genes that were greater than or equal to twofold up and the average normalized read count for the duf89Δ strain was ≥100; and (ii) genes that were greater than or equal to twofold down and the average normalized read count for the wild-type strain was ≥100.
Allelic replacement at the duf89 locus
We constructed strains harboring marked wild-type and mutated duf89 alleles as follows. First, we generated a pKS-based plasmid carrying a duf89 integration cassette marked with hygMX. The cassette consisted of the following elements, proceeding from 5′ to 3′: (i) a 620-bp segment of genomic DNA 5′ of the duf89+ start codon; (ii) a 1436-bp segment encompassing the duf89 ORF + intron and 44-bp of 3′-flanking genomic DNA; (iii) a 266-bp segment harboring the nmt1+ transcription termination signal; (iv) a hygMX gene conferring resistance to hygromycin; and (v) a 579-bp segment of genomic DNA 3′ of the duf89+ stop codon. The integration cassette for duf89-D252A was generated by replacing a restriction fragment spanning the Asp252 codon in the wild-type integration cassette with a restriction fragment containing the D252A missense mutation. All fragments were sequenced to exclude the presence of unwanted mutations. The integration cassettes (WT and D252A) were excised from plasmids and transfected into haploid S. pombe cells. Hygromycin-resistant transformants were selected and analyzed by Southern blotting to verify marker integration at the duf89 locus. The duf89-WT-hygMX and duf89-D252A-hygMX alleles were amplified by PCR and sequenced to confirm the presence of the desired mutation.
pTIN plasmid-based expression of Duf89
pTIN plasmids (Garg 2020) encoding the wild-type Duf89 or Duf89-D252A were transfected by the lithium acetate method into S. pombe cells. Control transfections were performed with the empty pTIN vector. Transformants were selected on Leu− enhanced pombe minimal glutamate (ePMG) 2% agar medium with 15 µM thiamine (Benjamin et al. 2022). Cells bearing pTIN plasmids were grown at 30°C in Leu− ePMG liquid medium with 15 µM thiamine. Aliquots of exponentially growing cultures were harvested, washed with water, and resuspended in water, and assayed for cell surface acid phosphatase activity. The data are averages (±SEM) of three independent biological replicates.
Allelic replacement at the aps1 locus
Strains harboring marked wild-type and mutated aps1 alleles were constructed as follows. The integration cassette for wild-type aps1 consisted of five elements in series from 5′ to 3′: (i) a 594-bp segment of genomic DNA 5′ of the aps1+ start codon; (ii) an open reading frame encoding wild-type Aps1; (iii) a 266-bp segment including polyA/termination signals from the nmt1+ gene; (iv) a kanMX gene conferring resistance to G418; and (v) a 543-bp segment of genomic DNA 3′ of the aps1+ stop codon. Two-stage PCR overlap extension with mutagenic primers was used to introduce missense mutations into the aps1 ORF and mutated DNA restriction fragments were inserted into the integration cassettes in lieu of the wild-type aps1 ORF. All inserts were sequenced to exclude the presence of unwanted mutations. The integration cassettes (WT, E89A, E93A, and E89A-E93A) were excised from plasmids and transfected into haploid S. pombe cells. G418-resistant transformants were selected and correct integrations at the target locus were confirmed by Southern blotting. A segment of the aps1-kanMX locus was PCR-amplified and sequenced to verify that the desired mutations were present.
DATA DEPOSITION
The RNA-seq data in this publication have been deposited in NCBI's Gene Expression Omnibus and are accessible through GEO Series accession number GSE216299.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
This work was supported by National Institutes of Health (NIH) grants R01-GM134021 (B.S.) and R35-GM126945 (S.S.). The content of this paper is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. A.M.S. is supported by National Science Foundation (NSF) graduate research fellowship 1746057.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079595.123.
- Received January 12, 2023.
- Accepted February 13, 2023.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Ana Sanchez is the first author of this paper, “Duf89 abets lncRNA control of fission yeast phosphate homeostasis via its antagonism of precocious lncRNA transcription termination.” Ana is a fourth year PhD student in Stewart Shuman's laboratory in the Molecular Biology program at Memorial Sloan Kettering Cancer Center in New York. The main focus of her thesis research is to investigate the biochemical and structural properties of fission yeast pyrophosphatase enzymes and elucidate their roles in regulating 3′ end processing and transcription termination by using the prt-pho1 system of transcriptional interference.
What are the major results described in your paper and how do they impact this branch of the field?
Previous studies by Wykoff and colleagues had implicated Duf89 (a domain of unknown function protein) in negatively regulating the expression of pho1, a gene that encodes a cell surface acid phosphatase, which is actively repressed in phosphate-replete conditions by the transcription in cis of the 5′-flanking prt long noncoding RNA (lncRNA). We show in this paper that Duf89 impacts pho1 expression by preventing premature termination of the prt lncRNA in a manner dependent on (i) cis-acting elements within prt lncRNA that are important for termination of the lncRNA transcript; (ii) an intact 3′ processing/termination machinery; (iii) the Thr4 phospho-site in the carboxy-terminal domain (CTD) of the largest subunit of RNA polymerase II Rpb1; and (iv) synthesis of the metabolite inositol pyrophosphate 1,5-IP8. Genetic interactions between Duf89, cleavage and polyadenylation complex (CPF) subunits, transcription termination factor Rhn1, the RNA polymerase II CTD, and inositol pyrophosphate metabolizing enzymes Asp1 and Aps1, show that Duf89 plays a broader role in 3′ processing/termination. We show that combining single mutations rpb1-CTD-S7A and duf89Δ that promote precocious termination is synthetically lethal, signifying that Duf89 and the CTD-Ser7-PO4 mark have redundant functions in avoiding precocious termination. This lethality can be bypassed by introducing an additional mutation that diminishes termination, such as rhn1Δ. Whereas combining CPF mutations ssu72-C13S and dis2Δ that lead to reduced termination is synthetically lethal, this lethality can be suppressed by duf89Δ that exerts an opposing termination effect. Previous work from our laboratory, in which we characterized the biochemical and structural properties of two fission yeast DUF89 paralogs (Duf89 and Duf8901), showed that Duf89 is a metal-dependent phosphatase/pyrophosphatase and that Duf89 catalytic activity requires the metal-binding aspartates Asp252 and Asp290. The present paper shows that the catalytic activity of Duf89 is not required for its function in vivo. In sum, our work establishes a new role for the Duf89 protein in 3′ processing and transcription termination.
What led you to study RNA or this aspect of RNA science?
3′ end processing (cleavage and polyadenylation) and transcription termination are essential steps in the expression of genes transcribed by RNA polymerase II. I was interested in studying this aspect of RNA biology because we still do not have a full understanding of how these steps impact cell physiology. My studies in fission yeast have shown that the prt-pho1 system of lncRNA-mediated transcription interference is sensitive to changes in 3′ processing/termination; thus, I am exploiting this platform to uncover new regulators of these events.
During the course of these experiments, were there any surprising results or particular difficulties that altered your thinking and subsequent focus?
My initial hypothesis was that Duf89 enzymatic activity was required for its effects on fission yeast phosphate homeostasis, and that accumulation of phosphorylated substrate(s) led to cytotoxicity in duf89Δ aps1Δ and duf89Δ rpb1-CTD-S7A cells. The surprising (and hypothesis-rejecting) result obtained during the course of my experiments was that the duf89-D252A mutation that abolishes phosphohydrolase activity had no consequence on any of the phenotypes tested. This shifted my thinking toward a noncatalytic function of the Duf89 protein, above and beyond its activity as a phosphatase.
What are some of the landmark moments that provoked your interest in science or your development as a scientist?
I am fascinated by the complexity of transcriptional regulation of gene expression. Our studies on the individual letters of the carboxy-terminal domain of RNA polymerase II and their role in governing distinct gene expression programs in fission yeast were critical to my development as a scientist.
If you were able to give one piece of advice to your younger self, what would that be?
Don't be afraid to step out of your comfort zone, and remember that failure is an option that teaches you more valuable lessons than success.








