
Structure of the dengue virus RNA promoter
- Corresponding author: varani{at}uw.edu
Abstract
Dengue virus, a single-stranded positive sense RNA virus, is the most prevalent mosquito-borne pathogen in the world. Like all RNA viruses, it uses conserved structural elements within its genome to control essential replicative steps. A 70 nt stem–loop RNA structure (called SLA), found at the 5′-end of the genome of all flaviviruses, functions as the promoter for viral replication. This highly conserved structure interacts with the viral polymerase NS5 to initiate RNA synthesis. Here, we report the NMR structure of a monomeric SLA from dengue virus serotype 1, assembled to high-resolution from independently folded structural elements. The DENV1 SLA has an L-shaped structure, where the top and side helices are coaxially stacked, and the bottom helix is roughly perpendicular to them. Because the sequence is highly conserved among different flavivirus genomes, it is very likely that the three-dimensional fold and local structure of SLA are also conserved among flaviviruses and required for efficient replication. This work provides structural insight into the dengue promoter and provides the foundation for the discovery of new antiviral drugs that target this essential replicative step.
Keywords
INTRODUCTION
Flaviviruses such as dengue (DENV), West Nile (WNV), yellow fever (YFV), and Zika virus (ZIKV) cause severe human diseases. Among them, dengue fever is the most prevalent mosquito-borne viral disease in humans (Clyde et al. 2006; Gubler 2006). It is estimated that up to 400 million people become infected with it each year (Murray et al. 2013). Any of the four dengue virus serotypes (DENV1 to DENV4) can produce clinical symptoms ranging from a flu-like syndrome to the severe and even fatal dengue hemorrhagic fever (Clyde et al. 2006; Gubler 2006). Despite the significant impact of dengue infection on human health, effective vaccines are not yet available after >70 yr of effort (Dyer 2017), and small molecule treatment is only beginning to show promise (Raut et al. 2015; Lin et al. 2017; Saleem et al. 2019).
Dengue, like all flaviviruses, is a single-stranded positive sense RNA virus with a genome of about 11 kb, with a 5′-type I cap but without a polyadenylated tail (Selisko et al. 2014; Kamau et al. 2019; Colavita et al. 2020; Dang et al. 2020; Pascalis et al. 2020). The viral genome encodes a long polyprotein that is subsequently processed by both host and viral proteases to generate 10 mature viral proteins. Three structural proteins, capsid (C), envelope (E), and prM, assemble new viral particles, while seven nonstructural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) are responsible for viral RNA replication (Klema et al. 2015; Garcia-Blanco et al. 2016; Ng et al. 2017; Mazeaud et al. 2018; Neufeldt et al. 2018). The NS5 protein consists of an amino-terminal methyltransferase (MTase) domain which is involved in capping, and a carboxy-terminal RNA-dependent RNA polymerase (RdRp) domain which is responsible for RNA synthesis (Ferrer-Orta et al. 2006; Yap et al. 2007; Zhao et al. 2015; Neufeldt et al. 2018).
The coding sequence is flanked by 5′- and 3′-UTRs, which contain highly conserved cis-acting RNA elements that are important for translation of viral proteins, RNA synthesis, encapsidation, and genome dimerization (Gebhard et al. 2011; Ng et al. 2017). In particular, the highly conserved stem–loop A (SLA) in the 5′-UTR, close to the 5′-end of the genome, functions as a “promoter” for the NS5 protein to initiate minus-strand synthesis (Filomatori et al. 2006; Gebhard et al. 2011; Choi 2021). The interaction between SLA and NS5 is critical for RNA synthesis and was confirmed to be direct by biochemical studies; removal of SLA and mutation of certain nucleotides drastically decreases RNA replication (Filomatori et al. 2006; Lodeiro et al. 2009). Consistent with its essential functional role, the sequence and secondary structure of the SLA are highly conserved (Fig. 1).
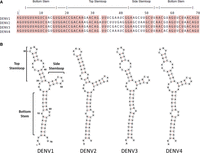
The sequence and secondary structure of the SLA promoter element that recruits the NS5 protein to initiate minus-strand RNA synthesis is very highly conserved among the four dengue virus serotypes (DENV1-4). (A) Alignment of the sequences of the first 70 nt of DENV1, DENV2, DENV3, and DENV4. The regions corresponding to the predicted secondary structure elements (bottom helix, top and side stem–loops) are indicated at the top. Conserved nucleotides are shaded. (B) Predicted RNA secondary structure of the SLA for the four DENV serotypes, as confirmed by SHAPE (Dethoff et al. 2018).
The current model for DENV RNA synthesis involves cyclization of the genome, mediated by long-range RNA–RNA interactions through inverted complementary sequences near the opposite ends of the RNA, which position the 5′- and 3′-UTRs in close proximity (Alvarez et al. 2005b; Filomatori et al. 2006; Villordo et al. 2010; Filomatori et al. 2011). This interaction allows the SLA-bound NS5 to transfer to the 3′-stem–loop (3′-SL) at the end of the 3′-UTR and initiate minus-strand synthesis (Hodge et al. 2016; Mazeaud et al. 2018). Removal of the SLA or 3′-SL leads to a significant reduction of viral replication (Alvarez et al. 2005a; Filomatori et al. 2006; Yu et al. 2008), suggesting that a small molecule targeting these RNAs could have antiviral activity.
In all flaviviruses, the SLA forms a Y-shaped secondary structure, consisting of a bottom helix with a conserved U-rich bulge, a short side stem–loop and an apical stem–loop interrupted by an internal loop (Fig. 1; Gritsun and Gould 2007; Dethoff et al. 2018). A crystal structure of DENV2 SLA was determined using a chimeric RNA, wherein the DENV2 SLA was inserted into the anticodon loop of a human tRNA (Lee et al. 2021). It revealed a large L-shaped dimer, where the side stem–loop of each monomer is base-paired with another molecule in the crystal. It is unclear whether dimerization is an artifact of crystallization, or a relevant functional state, perhaps transient, of the virus.
Here we report the NMR structure of the monomeric DENV1 SLA. Because of the severe peak overlap caused by the large size of the RNA, we use a divide-and-conquer approach to assemble a high-resolution structure from three independently folded structural elements corresponding to two of the three stem–loops and the three-way junction (Fig. 2), which were then assembled to generate the full SLA RNA through NMR analysis of the complete element (Barnwal et al. 2016). The SLA has an L-shaped structure, consistent with the crystal structure, where the top and side stem–loops are coaxially stacked and the bottom helix is roughly perpendicular to it. We only observe formation of a dimeric conformation at near mM NMR concentrations, above 0.5 mM; suggesting that if a dimer does indeed form in the cell, it is not because of its thermodynamic stability. Our structure will facilitate understanding of the mechanism of dengue virus replication and provides the foundation for the discovery of new antiviral drugs, which is already underway in the group.
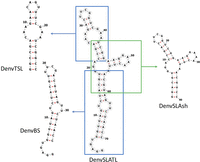
The sequences and secondary structures of each RNA segment studied in this work. Three smaller constructs, corresponding to independently folded secondary structure domains, were prepared to facilitate structure determination: DenvBS represents the bottom helix; DenvTSL the top stem–loop; and DenvSLAsh the three-way junction. Secondary structures for each segment were predicted using the UNAFold web server (http://www.unafold.org) (Markham and Zuker 2008; Zuker 2003) and verified by the NMR assignments.
RESULTS
Construct design
The spectra of the complete SLA RNA are characterized by extensive spectral overlap and broad lines because of its relatively large size, 70 nt, causing difficulties in obtaining unambiguous peak assignment and, especially, collecting a large number of constraints for structure determination (Supplemental Fig. S1). Thus, we adopted the strategy of reconstructing the complete structure by analyzing smaller, independently folded structural elements (divide-and-conquer) (Barnwal et al. 2016). This can be done because RNA structure is modular, and its folding hierarchical; in other words, secondary structural elements generally fold independently outside of the context of the complete RNA (Tinoco and Bustamante 1999). This is very different from proteins, where secondary and tertiary structure folding generally cannot be separated. Nevertheless, the validity of this approach has to be experimentally verified in each case, which we do as described below.
We divided the DENV1 SLA into three structural segments that overlap to generate the complete promoter: the bottom helix (DenvBS), the top stem–loop (DenvTSL), and the three-way junction (DenvSLAsh) (Fig. 2; Supplemental Fig. S2). A few additional G–C base pairs were added at the end of the sequences to improve in vitro transcription and stabilize the local secondary structure, if needed. The addition of tetraloops stabilizes RNA structure and, especially, improves linewidth with multiple RNAs we have studied in the past (Barnwal et al. 2016; Sharma and Varani 2020; Walker et al. 2020). Thus, a UUCG tetraloop was added to the top of DenvBS, while UUCG and GAAA tetraloops were added to the top and side stem–loop of DenvSLAsh, respectively. For the complete SLA, we also incorporated UUCG and GAAA tetraloops into the top and side stem–loop of DENV1 SLA, respectively, to generate a stabilized SLA which we named DenvSLATL (Fig. 2; Supplemental Figs. S1, S2), and to avoid dimer formation through the side stem–loop, as described next. The top terminal loop is an important site for SLA function (Filomatori et al. 2006, 2011; Lodeiro et al. 2009); thus the top loop of DenvTSL was retained as wild type.
The structure of the individual domains recapitulates what is observed in the full SLA
The divide-and-conquer approach is only warranted if the structure of individual domains faithfully recapitulates what is observed in the complete RNA. For each of the smaller structural segments, imino proton peaks of base-paired residues predicted from the secondary structure were observed in H2O NOESYs, except for fast exchanging imino protons for unpaired nucleotides and the base pairs at the end of helical stretches (Supplemental Fig. S3). Assignments of these imino protons allowed us to establish and verify the predicted secondary structures for each domain.
High-quality NMR spectra could be collected for a tetraloop-stabilized SLA model that avoids dimerization through the side stem–loop (see below, DenvSLATL), and peak assignment was facilitated by the assignments of each segment. Reassuringly, we find that each NH in the smaller segments has a similar chemical environment to its corresponding NH in the complete RNA, thus presenting similar chemical shifts and verifying that the structure observed in the complete SLA is retained in the individual fragments. Namely, overlaying the NOESY NMR spectra of the segments on the DenvSLATL spectra revealed high similarities in chemical shifts and NOE patterns (Fig. 3), which confirms the secondary structure of the fragments coincide with what is seen, for the corresponding domain, in the complete RNA.
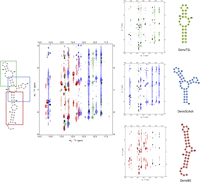
Overlay of the imino region of NOESY spectra for DenvSLATL (black) and the three smaller constructs which were prepared to facilitate structure determination. Black color corresponds to DenvSLATL, red to DenvBS, the bottom helix; green corresponds to DenvTSL, the top stem–loop, and blue corresponds to DenvSLAsh, the three-way junction. 1H NOESY spectra were recorded in 10 mM potassium phosphate buffer (pH 6.5, 90% H2O/10% D2O) at 25°C. Despite small differences, a high similarity of chemical shifts and NOE patterns were observed, allowing the transfer of peak assignments from each segment to the SLA model oligonucleotide (Supplemental Fig. S3).
NOESY spectra in D2O were then collected to assign nonexchangeable protons. Extensive overlaps in the sugar proton region were relieved by deuteration of H3′, H4′, H5′, H5″, and H5 protons (Supplemental Figs. S4–S6), even if this approach prevented assignments of most of those protons. 15N- and 13C-labeled samples were prepared for DenvTSL to record 1H-15N HSQC and 1H-13C HSQC to distinguish ambiguous peaks in NOESY spectra, as well. Through deuteration, we were able to observe characteristic sequential NOEs in helices and assign nonexchangeable protons for each of the separate Denv structural segments (Supplemental Figs. S4–S6).
The existence of a single dominant and monomeric conformation was confirmed in each case from the number of Ura and Cyt H5–H6 peaks in TOCSY spectra, which was in all cases consistent with the sequence (Supplemental Fig. S7). Formation of base-paired double helices were confirmed by cross-strand NOEs involving imino resonances, as well as sequential NOEs involving both exchangeable and nonexchangeable protons (Supplemental Figs. S4–S6). Thus, DenvBS and DenvTSL were confirmed to form stem–loop structures as expected; the monomeric/dimeric state of the three-way junction is discussed below.
NMR structures of the individual structural elements of DENV1 SLA RNA
Once assignments were completed, NOE distance constraints were systematically tabulated for structure determination, and the constrain list was refined by multiple rounds of structure calculations. For each of the three structural segments, structure calculations were performed independently using distance and torsion angle restraints derived from NMR experiments, as summarized in Supplemental Tables S1–S3. Once the segments were completed, restraints from corresponding nucleotides in each segment were also added to the restraint table for structure calculation of the complete monomeric SLA (see below).
RNA structures were calculated with Xplor-NIH using torsion angle dynamics and simulated annealing starting from randomized coordinates against the restraint table (Schwieters et al. 2003). A total of 1000 independent calculations were executed, and the lowest energy structure was further refined by an extended simulated annealing calculation against the experimental restraints. Four hundred structures were eventually generated, and the 10 lowest scoring structures were selected for presentation for each RNA molecule. The resulting structures converge well, with no NOE restraint violation greater than 0.5 Å. Representative structures for DenvSLA segments were presented with PyMol (Fig. 4), and the overall heavy atom RMSDs are listed in Supplemental Tables S1–S3 as well.
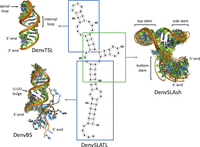
Superposition of the 10 NMR structures of lowest energy for each independently folded RNA segment studied in this work. DenvBS represents the bottom helix and forms a well-defined double helix with an unstructured 5′-tail (G1–U5, in cyan) and a U-rich bulge; DenvTSL represents the top stem–loop and forms a short helix interrupted by an internal loop, capped by a partially flexible loop; DenvSLAsh represents the three-way junction and forms an L-shaped structure, where the top and side stem helices stack coaxially, and the bottom helix is roughly perpendicular to the coaxial stack; this last image was generated by superposing only the two stacked helices to emphasize the coaxial stack and partial flexibility at the three-way junction.
As predicted, DenvBS and DenvTSL form double helical stems interrupted by internal loops. In both cases, no direct evidence of noncanonical base-pair formation in the internal loops was found, but sequential aromatic NOE correlations (H6/H8–H6/H8) in those regions were observed (Supplemental Figs. S4, S5), together with other sugar-base NOEs, implying the retention of helical stacking throughout the internal loop. Many of the unpaired nucleotides are therefore stacked inside the helices, and potentially base-paired, but we lack the information to impose such constraints, perhaps because any noncanonical base pairs are only transiently formed.
The results provide insight into the local structure in these regions. In the U-rich internal loop of DenvBS, Ura12 and Ura30 are coplanar and Ura29 stacks between Gua28 and Ura30 (Fig. 5A). Nucleotides in the internal loop of DenvTSL (Cyt7 to Ade9 and Gua20 to Ade22) also stack within the helical stem, retaining coaxial stacking (Fig. 5B). Although the terminal loop of DenvTSL varies somewhat within the 10 calculated structures, the topology is clearly established. The Gua15 base points toward the major groove, while Ade14 and Ura16 point outward, in the direction of the solvent, and could conceivably provide direct interactions with the RdRP or other accessory proteins (Fig. 5C). The significance of each region in dengue SLA has been validated by biological studies and is discussed further in the Discussion section below.
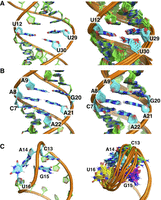
(A) Close-up view of the U-UU bulge in DenvBS, drawn from the lowest energy structure (left) and superposition of 10 representative NMR structures (right). Ura12 and Ura30 are coplanar, potentially forming an unstable base pair (no NH peak is visible in 2D spectra), while Ura29 stacks between Gua28 and Ura30, retaining continuous stacking. (B) A close-up view of the internal loop in DenvTSL, taken from the lowest energy structure (left) and the superposition of 10 representative NMR structures (right). Nucleotides Cyt7–Ade9 and Gua20–Ade22 across the internal loop stack within the helix. Although no evidence of noncanonical base-pair formation in this region was observed in NMR spectra, uninterrupted sequential NOEs were observed (Supplemental Fig. S5). (C) A close-up view of the terminal loop in DenvTSL from the lowest energy structure (left) and the superposition of 10 representative NMR structures (right). In this last image, nucleotides are colored differently for clarity (C13 in green, A14 in cyan, G15 in magenta, and U16 in yellow). In all 10 structures, Gua15 points toward the major groove while Ade14 and Ura16 point outward toward the solvent, and Cyt13 is poorly defined.
Structure of the three-way junction
The key structural element in SLA is the three-way junction which organizes the complete element; because a dimer is observed in the DENV2 crystal structure, it remains unclear what the three-dimensional organization of the full SLA would be. In order to investigate the three-way junction in DENV1 SLA with the necessary resolution, we prepared an RNA containing the sequence of the three-way junction and side stem–loop, but with shortened top and bottom helical stems to reduce spectra overlap. Furthermore, two RNAs were synthesized, one containing the wild-type side stem–loop (SLAshCUUG) and the other containing a GAAA-stabilized side stem–loop (DenvSLAsh) (Fig. 2; Supplemental Fig. S1). This was done because we observed extra base pairs in the NMR spectra of SLAshCUUG. The signal intensity for those extra base pairs is concentration dependent, which indicates a monomer-dimer equilibrium at the 0.5–1.5 mM concentration of our NMR experiments (Supplemental Fig. S8A). By substituting CUUG with a GAAA tetraloop, dimerization was eliminated (Supplemental Fig. S8B).
In the NOESY spectra of SLAshCUUG, we did not observe NH corresponding to base-paired nucleotides for the side stem–loop or cross-strand peaks involving the AH2s. Nevertheless, sequential H1′–H6/H8 and H2′–H6/H8 were observed for the helical sections of the side stem–loop, which indicate stacking and perhaps unstable base pairing. We presume that with a less stabilizing loop sequence, the side stem–loop in monomeric SLAshCUUG forms an unstable short helix, allowing the dimer to form through RNA–RNA interactions in the side stem–loop at the mM concentrations of NMR experiments (Supplemental Fig. S8C). Considering the low copy number of viral RNAs in the cell, it is very unlikely that the dimer is the thermodynamically favored SLA structure, and therefore we collected NMR spectra of the monomeric DenvSLAsh for structural analysis. However, we cannot exclude the possibility that dimer formation occurs at intermediate steps of viral replication; this is further discussed below.
Base-pair imino NOEs and characteristic sequential NOE patterns for helical stem nucleotides were observed in NOESY spectra for the three double helical stems (Supplemental Figs. S3, S6), validating the secondary structure prediction. For nucleotides Gua7, Gua8, and Ade9, only sequential NOEs were observed, but no other interstranded or internucleotide NOEs were identified. NOE interactions corresponding to Gua23H8–Ade24H8, Ura33H6–Ade34H8, and Ade34–Cyt10H6 were observed in NOESY spectra as well (Fig. 6A), consistent with continuous stacking between the top and side stem–loops, and suggestive of coaxial stacking of the two helices.
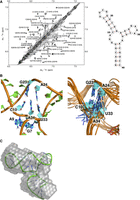
(A) NOESY spectrum of DenvSLAsh showing H6/H8–H6/H8 interactions; the secondary structure of DenvSLAsh is shown on the right. This spectrum was recorded at 25°C in 10 mM potassium phosphate buffer (pH 6.5, 90% H2O/10% D2O). The following NOE interactions, G23H8–A24H8, U33H6–A34H8 and A34–C10H6, within the three-way junction establish coaxial stacking of the top and side stem–loops. (B) A close-up view of the three-way junction in DenvSLAsh from the lowest energy structure (left) and superposition of 10 representative NMR structure (right). Ade34 stacks within the coaxial helix, between Cyt10 at the end of top stem and Ura33 at the end of the side stem–loop. (C) The lowest energy structure of DenvSLAsh is shown in cartoon representation superposed on the SAXS model; the SAXS results were not used for NMR refinement and therefore provide independent validation of the structure.
The calculated structure of DenvSLAsh demonstrates formation of an L-shaped three-way junction, where the top and side stem–loops stack coaxially, and the bottom helix is roughly perpendicular to the coaxial stack (Fig. 4). Ade34 in the three-way junction stacks within the coaxial helix, between Cyt10 in the top stem and Ura33 in the side stem. Gua7, Gua8, and Ade9 are instead single stranded, giving the three-way junction some flexibility, and thus allowing the angle of the bottom helix to wiggle relative to the top coaxial stack (Fig. 6B).
The topology of the three-way junction structure was independently validated by SAXS analysis (Fig. 6C; Supplemental Fig. S9). Both SLAshCUUG and DenvSLAsh exhibit L-shaped SAXS envelopes, which agree very well with the L-shaped NMR structure and provide independent validation. They also demonstrate that the structure is fully monomeric below 0.1 mM concentration.
Structure of the Denv1 promoter
By examining each of the independently folded secondary structure elements separately, we were able to establish the local structures for the two stem–loops and three-way junction and side stem–loop that form Denv1 SLA. Spectral and structural information collected from these spectra were used to assist peak assignments and structure determination for the complete SLA-structure.
A single conformation of DenvSLATL was confirmed by the number of Ura and Cyt H5–H6 peaks in TOCSY spectra (Supplemental Fig. S10). Overlay of the NOESY and TOCSY spectra of DenvSLATL with those of its structural segments revealed very similar chemical shifts and NOE patterns (Fig. 3; Supplemental Fig. S10), which allowed us to safely transfer the much larger number and more confidently assigned restraints obtained from the individual segments. Structure calculations were then performed for DenvSLATL using distance and torsion angle restraints derived from experimental NMR data of DenvSLATL and each of the structural segments, as was done for each of the elements in the structure. The 10 lowest structures converge with no NOE restraint violation greater than 0.5 Å, and then independent SAXS analysis was performed to validate the structure. Importantly, the topology of the structure of DenvSLATL was independently validated by SAXS analysis, and the SAXS envelope agrees very well with the L-shaped NMR structure (Fig. 7). The structural statistics are summarized in Supplemental Table S4.
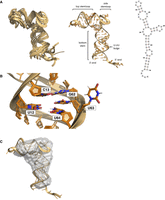
(A) Superposition of the 10 NMR structures of DenvSLATL (left), the structure with the lowest calculated energy (middle), and the secondary structure (right) are also shown. (B) A close-up view of the U-rich bulge of DenvSLATL. Two conformations of U-UU bulge were observed in the calculated NMR structures, as shown in the image. (C) The lowest energy structure of DenvSLATL in cartoon representation superposed on the SAXS envelope; the SAXS results were not used for refinement and therefore provide independent validation of the NMR structure.
The calculated structure of DenvSLATL reveals an L-shaped RNA, where the top and side stem–loops are coaxial and the bottom stem roughly perpendicular to it (Fig. 7A), as per the smaller three-way junction. The arrangement of nucleotides in the three-way junction is as observed in DenvSLAsh: Ade56 stacks between the base-pairs of Cyt22–Gua45 and Ade46–Ura66, which are at the end of the top and side stem–loop, respectively, whereas nucleotides in the longer internal junction loop (Gua19 to Ade21) are single stranded, with fewer NOE interactions, and thus the orientation of the bottom stem is less well defined. The U-rich bulge in the bottom stem is important for SLA function. In our NMR structures, Ura12 and Ura64 are coplanar and stack within the helix. However, two conformations of Ura63 were observed in the calculated structures: one with the base within the helix and another with the base pointing outward (Fig. 7B). Eight out of the lowest 10 structures have Ura63 stacked within the helix and two structures have Ura63 pointing outward, suggesting conformational flexibility.
DISCUSSION
The Denv SLA structure functions as an RNA “promoter” for the viral RNA-dependent RNA polymerase enzyme NS5 and is essential to viral replication. NS5 is recruited to the viral RNA through this element, which has high conservation in both sequence and secondary structure across flaviviruses (Filomatori et al. 2006; Gebhard et al. 2011; Choi 2021). The presence of a U-rich bulge in the bottom helix is essential for SLA function, and the high conservation of the three-way junction supports a functional role for the overall 3D shape of the RNA, which is determined by the topology of the junction. We observe a rigid conformation for the three-way junction, with coaxial stacking of the top and side stem–loops, to generate a well-defined L-shaped structure, with the bottom helix emanating at a nearly 90° angle, but with some flexibility because of the single-stranded nucleotides linking the bottom and top helices (Fig. 7A).
Flexibility could be the result of insufficient experimental information (e.g., due to spectral overlap) or intrinsic dynamics. One way to look at dynamics without recording relaxation data, which would be very challenging for an RNA this size, is to examine the conformation of the sugar pucker, as deduced from the TOCSY spectra. For 3′-endo sugar pucker, the H1′–H2′ coupling is very small and generally no peak is observable, especially for an RNA this size. The 2′-endo conformation has strong H1′–H2′, medium H1′–H3′ peaks and very weak/unobservable H1′–H4′ peaks. For a mixture of 3′- and 2′-endo conformations, which is a sign of dynamics, peaks from H1′ to H2′, H3′ and H4′ are observed. According to the TOCSY spectra (Supplemental Fig. S12), Ura29 in the U-rich bulge occupies the 2′-endo conformation; Ade9 and Gua8 in the three-way junction are 2′-endo as well. This is interesting, since the 2′-endo conformation extends the phosphate-phosphate distance. Unfortunately, we cannot establish the conformation of Ade34 in the loop between the side and bottom stems, because of spectral overlap around 4.4 ppm; this is an important nucleotide to determine the orientation of the lower helix with respect to the rest of the structure. The cytosine in the top loop is 2′-endo. Only the sugar of the adenine that follows occupies a mixture of 2′ and 3′-endo conformations, which implies a dynamic structure for this nucleotide. It is possible that conformational flexibility plays a role in SLA function, since RNA-binding proteins often exploit induced fit (Leulliot and Varani 2001), but this remains to be investigated.
In the structure, Ura63 in the U-rich bulge occupies two conformations; the base can point inward or outward, but the two conformations do not affect the rest of the RNA structure (Fig. 7B); in the recently reported crystal structure of a dimeric Denv2 SLA, the same Uracil points outward (Lee et al. 2021). The fact that DENV3 has a single U bulge in the bottom stem and mutational studies of the U-UU bulge implies that Ura63 is critical for SLA promoter function, but the two remaining nucleotides are not. Deletion or mutations of the U-rich bulge largely impair viral RNA replication in vivo, but the effect on the binding affinity to RdRp and its in vitro activity is insignificant (Lodeiro et al. 2009; Filomatori et al. 2011). This suggests that the U-rich bulge interacts with another protein, which is important for viral replication in infected cells.
The apical loop is important for SLA function and highly conserved; the CAG(X)U sequence is found in all four dengue serotypes (Fig. 1), and mutations impair both viral replication in vivo and RdRp activity in vitro (Filomatori et al. 2006, 2011; Lodeiro et al. 2009). Interestingly, mutations in the terminal loop do not significantly affect RdRp binding, suggesting the terminal loop might play an important role in post-binding steps to promote polymerase activity (Filomatori et al. 2011). In the structure, we observe that Gua32 points toward the major groove and faces the other two loop nucleotides Ade31 and Ura33, which point outward (Fig. 5C). The orientation of the loop nucleotides could be important for RdRp activity. The top stem, however, is less conserved among dengue serotypes. In vivo biological assays have shown that the presence of an internal loop in the top stem has no significant effect on SLA function either (Lodeiro et al. 2009), yet shortening the helix significantly decreases viral replication and RdRp binding (Lodeiro et al. 2009; Filomatori et al. 2011). This implies that, although the helical stem is unlikely to provide direct interactions with the NS5 protein, it is important to space the terminal loop relative to the three-way junction and the U-rich bulge.
The three-way junction is likely to provide a structural framework to orient the SLA and different domains of NS5, giving it an L-shaped conformation confirmed by the SAXS analysis. The top and side stem–loops are coaxially stacked, as expected, since no single-stranded nucleotide is found at the junction between them (Lescoute and Westhof 2006). The stacking of Ade56 in the linker between side and bottom stem constrains the direction of the bottom stem to be roughly perpendicular to the coaxially stacked helix.
The recently reported crystal structure of Denv2 SLA also presents an L-shaped structure, but this is generated by dimerization in the crystal created by the unpaired side loop which forms “kissing loop” interactions (Lee et al. 2021). In the crystal structure, the side loop was open and engaged in loop–loop interactions. We observe two conformations for a construct containing the wild-type loop sequence, consistent with monomer-dimer equilibrium. Replacement of the wild-type loop with GAAA stabilizes the monomer and allowed us to establish the structure of a monomeric SLA RNA. It remains to be seen whether SLA dimerization is of functional relevance. Because we observe increased dimer peak intensity at the NMR concentrations, 0.5–1.5 mM, and no dimer below 0.5 mM, we presume that under cellular conditions, the SLA RNA will be entirely monomeric. However, this conclusion does not preclude the possibility that the dimeric structure is transiently present as a result of protein binding or dimerization of the genome, and in fact it could be an elegant way for the virus to regulate its promoter activity.
We expect the 3D structure of the three-way junction to be conserved in flaviviruses (Fig. 1; Supplemental Fig. S13), because no nucleotides are predicted to be present in the junction between top and side stem in all flavivirus sequences, and the linker between side and bottom stem is always short, whereas the longest linker occurs invariably between bottom and top helices. It follows that the shape of the SLA and the orientation of the top and bottom stems are likely important for SLA to be recognized by the viral polymerase.
In summary, we have established the 3D structure of the thermodynamically favored, monomeric form of the SLA promoter from dengue virus serotype 1; which is very likely to be representative of all other serotypes, and indeed it is very likely all flaviviruses will share the same global structural arrangement. In addition to providing a framework for interpreting biochemical data and NS5 activity, this structure also lays the groundwork to identify small molecule inhibitors that target the highly conserved three-way junction, which are being actively pursued in our group.
MATERIALS AND METHODS
RNA preparation
All RNA molecules were synthesized by in vitro transcription with in house purified T7 RNA polymerase using synthetic DNA oligonucleotide templates (purchased from Integrated DNA Technologies) and standard methods (Milligan et al. 1987). Partially deuterated RNA molecules (deuteration of H5, H3′, H4′, H5′, and H5″ protons) were synthesized using selective deuterated NTPs (from Cambridge Isotopes). 15N- and 13C-labeled samples were synthesized using isotope-labeled NTPs (from Cambridge Isotopes). The sequence and secondary structures of all the RNAs, as verified by NMR, are shown in Supplemental Figure S2.
The RNA oligonucleotides were purified by denaturing polyacrylamide gel electrophoresis (PAGE), electroeluted and concentrated by ethanol precipitation (Gubser and Varani 1996). After extensive dialysis into 10 mM potassium phosphate buffer (pH 6.5), the RNAs were annealed by heating briefly to 90°C followed by snap cooling in an ice bath. Final RNA concentrations used for NMR studies were 0.6 ∼ 1.2 mM. For experiments studying nonexchangeable protons, samples were lyophilized to dryness and dissolved into D2O. Samples used to study exchangeable protons were dissolved in H2O:D2O (9:1).
NMR spectroscopy
All NMR spectra were collected at 25°C on Avance III 600 MHz, Avance III 700 MHz or Avance III 800 MHz spectrometer equipped with cryogenic probes (600 and 800). The 1D 1H spectra were recorded using the excitation sculpting pulse sequence. 2D total correlation spectroscopy (TOCSY) spectra were recorded with mixing times of 80 msec. The exchangeable and nonexchangeable 2D NOESY spectra were recorded with various mixing times (100, 200, and 300 msec) to assist spectral assignments and quantitative evaluation of internuclear distances by comparison with peak intensities for pairs of protons with fixed distances. Spectra for selectively deuterated samples were collected in the same manner. 2D 1H-15N and 1H-13C HSQC spectra were recorded on isotope labeled samples, if needed to confirm assignments. All NMR data were processed with TopSpin (Bruker) and analyzed in NMRFAM-SPARKY (Lee et al. 2015). Assignments of RNA spectra were guided by predicted RNA chemical shift values and based on well-established double-helical sequential NOE patterns (Varani and Tinoco 1991; Varani et al. 1996).
Experimental restraints and structure determination
Interproton distance restraints were derived from NOE cross-peaks in 2D 1H NOESY spectra and sorted into strong (2.5 ± 0.7 Å), medium (3.5 ± 1.2 Å), and weak (4.5 ± 1.5 Å) bins based on peak intensities, relative to fixed distances (e.g., H5–H6 = 2.5 Å, H3′–H6/H8 = 3.5 Å). Base-pair planarity and hydrogen-bonding restraints were used for unambiguously established base pairs as identified by 2D NOESY involving NH protons. Hydrogen bond, planarity and dihedral restraints were included for base-paired nucleotides that were surrounded by base pairs conforming to A-form helical structures, as established from the pattern of NOE cross-peaks (Varani and Tinoco 1991; Varani et al. 1996).
We have often found within the group that flexible loops capping stem–loops lead to loss of spectral quality, most likely due to nonspecific aggregation or multimerization at NMR concentrations (Varani et al. 1991; Barnwal et al. 2016); this was the case for this RNA as well, with the added complication of dimerization through kissing loop interactions which created multiple conformations (Supplemental Fig. S1). Thus, UUCG and GAAA tetraloops were used to replace the dynamic apical loop or added to the end of the bottom helix to improve spectra quality (Varani et al. 1991; Jucker and Pardi 1995; Banas et al. 2010).
Experimental constraints for structure calculation of the complete DenvSLATL were compiled by dividing the RNA into three segments: DenvBS, DenvTSL, and DenvSLAsh (Fig. 2), corresponding to the bottom helix, apical stem–loop and three-way junction and side stem–loop. Overlay of 2D 1H–1H NOESY spectra from the segments, with a spectrum of the complete dengue 1 SLA model (called DenvSLATL), showed strong similarities in the chemical shifts, which allowed the transfer of segments assignments to DenvSLATL for assignments for structure calculations (Fig. 3; Supplemental Fig. S3). Restraints derived from DenvBS spanned nucleotides G1–U18 and A57–A70; those obtained from DenvTSL spanned nucleotides C22–G45; DenvSLAsh contributed information around the three-way junction, namely A15–A25 and U42–U60; obviously, there is overlap which allowed us to further verify that the divide-and-conquer approach was warranted. The NMR experimental constraints are summarized in Supplemental Tables S1–S4.
RNA structures were calculated with Xplor-NIH using torsion angle dynamics and simulated annealing from a single extended RNA starting template (Schwieters et al. 2003). Compiled experimental restraints were used in a simulated annealing procedure, initially undergoing high-temperature (2500 K to 298 K) torsional angle dynamics, where incremental decreases in temperature were generated by progressively introducing Van der Waals terms and increasing force constants for angles, dihedral angles, NOEs, and the Van der Waals repulsive term. After the final cooling step, the RNA underwent two sequential refinement steps, first in torsional angle space then in Cartesian space. A total of 1000 independent calculations were executed for each RNA, and the lowest total energy structure calculated by the Xplor energy function was further refined by an extended simulated annealing calculation against the constraints listed above to generate 400 structures. The 10 structures with the lowest total energy, based on the Xplor energy function, were chosen as the representative structures to calculate RMSD. The structure quality analysis was conducted using MolProbity (Williams et al. 2018). Representative structures of DenvBS, DenvTSL, DenvSLAsh, and DenvSLATL were deposited in the Protein Data Bank with accession codes: 7K4L, 7UME, 7UMD, and 7UMC, respectively.
Small-angle X-ray scattering (SAXS)
RNA samples for SAXS were prepared similarly to samples made for NMR spectroscopy, but at different concentrations (1–5 mg/mL), dissolved in 20 mM Tris, 100 mM NaCl, and 0.1 mM EDTA (pH 6.5). SAXS experiments were recorded on an in-house state-of-art SAXS instrument (BioSAXS-2000) at Argonne National Laboratory. The data were processed using RAW, and particle distance distribution function P(r) plots were calculated using GNOM (Svergun 1992) and used for low resolution ab initio shaped reconstruction with DAMMIN (Svergun 1999). Processed SAXS data curves of DenvSLAsh, SLAshCUUG, and DenvSLATL are shown in Supplemental Figures S14–16. A total of 20 models were generated with DAMMIN using the ATSAS online server (https://www.embl-hamburg.de/biosaxs/) (Manalastas-Cantos et al. 2021). The representative model was selected with a suite of software tools (DAMSEL, SAMSUP, DAMAVER and DAMFILT) for comparison and fitting to the NMR structure (Volkov and Svergun 2003).
DAMSEL compares models, finds the most probable model and identifies outliers; DAMSUP aligns all models with the most probable model; DAMAVER averages these aligned models and computes a probability map; and DAMFILT filters the average model at a given default cut-off volume, which is the expected volume of the generated PDB file. DAMFILT removes loosely defined and lower occupancy atoms and generates a most probable compact model.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
COMPETING INTEREST STATEMENT
G.V. is cofounder of Ithax Pharmaceuticals and Ranar Therapeutics.
ACKNOWLEDGMENTS
We wish to thank all members of the Varani group for discussions and support and Dr. Greg Olsen for help with final preparation of the manuscript. The study was supported by National Insitutes of Health (NIH) grant 1 R35 GM126942. This project has also been funded in part with federal funds from the National Institute of Allergy and Infectious Diseases, National Institutes of Health, Department of Health and Human Services, under contract no. HHSN272201700059C. We acknowledge the use of the SAXS Core facility of the Center for Cancer Research, National Cancer Institute (NCI) of National Institutes of Health (NIH). The SAXS core resource has been funded in whole or in part with federal funds from NCI under contract 75N91019D00024 and the Intramural Research Program of the NIH, National Cancer Institute, Center for Cancer Research. The content of this publication does not necessarily reflect the views or policies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organizations imply endorsement by the U.S. Government. The SAXS data were collected at beamline 12-ID-B of the Advanced Photon Source (APS) of Argonne National Laboratory (ANL). We thank Dr. Yu-Xing Wang (NCI) and Dr. Lixin Fan (FNLCR/Leidos) and Dr. Xiaobing Zuo (ANL) for their support. Use of the APS was supported by the U.S. Department of Energy, Office of Science, Office of Basic Energy Sciences, under contract no. DE-AC02-06CH11357.
Author contributions: Y.-T.S.: Conceptualization, methodology, investigation, writing, reviewing and editing, and graphics. G.V.: Experimental design, data analysis, writing, reviewing and editing.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079197.122.
- Received April 14, 2022.
- Accepted June 15, 2022.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is a new editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Yi-Ting (Tina) Sun is the first author of this paper, “Structure of the dengue virus RNA promoter.” Tina is currently a PhD candidate in the Varani Group in the Department of Chemistry, University of Washington. The focus of her research is the structural biology of RNA molecules.
What are the major results described in your paper and how do they impact this branch of the field?
In this paper, we report the high-resolution NMR structure of a monomeric form of the promoter SLA from dengue virus serotype 1, which is critical for its viral replication. Because of sequence conservation, this structure is very likely to be representative of all other dengue serotypes. It is also very likely that all flaviviruses will share the same global structural arrangement. This structure can potentially help us understand its function and provide a base for small molecule drug discovery, which is already underway in our group.
What led you to study RNA or this aspect of RNA science?
The research of RNA structural biology and functions is still a very new field compared to peptides and proteins. RNAs are related to many human diseases and virus infection, yet our understanding of them is still limited. I find this area of research very challenging, but also very interesting and promising.
During the course of these experiments, were there any surprising results or particular difficulties that altered your thinking and subsequent focus?
Structure determination of an RNA of this size (70 nt) using NMR spectroscopy is itself challenging. We used a divide-and-conquer strategy to overcome the challenge and successfully assemble the high-resolution structure of DNEV1 SLA. Another difficulty we faced was the dimerization due to the high concentration of RNA in NMR samples. The dimerization caused unexpected extra peaks, which I originally thought were caused by contamination of the sample. It took us some time to figure out that the real reason is dimerization.
If you were able to give one piece of advice to your younger self, what would that be?
Don't be afraid to fail because you can always learn more from a failure than a success.
What are your subsequent near- or long-term career plans?
I will graduate from my PhD program this year. I like to do experiments and research. Thus, I am looking for a research position in the biological or pharmaceutical industry where I can continue my interest in research and contribute to human health after graduating from school.








