
Pseudouridine formation in archaeal RNAs: The case of Haloferax volcanii
- Ian K. Blaby1,4,5,
- Mrinmoyee Majumder2,4,
- Kunal Chatterjee2,
- Sujata Jana2,
- Henri Grosjean3,
- Valérie de Crécy-Lagard1,6 and
- Ramesh Gupta2,6
- 1Department of Microbiology & Cell Science, University of Florida, Gainesville, Florida 32611-0700, USA
- 2Department of Biochemistry and Molecular Biology, Southern Illinois University, Carbondale, Illinois 62901-4413, USA
- 3Université Paris 11, IGM, CNRS, UMR 8621, Orsay, F 91405, France
-
↵4 These authors contributed equally to this work.
Abstract
Pseudouridine (Ψ), the isomer of uridine, is commonly found at various positions of noncoding RNAs of all organisms. Ψ residues are formed by a number of single- or multisite specific Ψ synthases, which generally act as stand-alone proteins. In addition, in Eukarya and Archaea, specific ribonucleoprotein complexes, each containing a distinct box H/ACA guide RNA and four core proteins, can produce Ψ at many sites of different cellular RNAs. Cbf5 is the core Ψ synthase in these complexes. Using Haloferax volcanii as an archaeal model organism, we show that, contrary to eukaryotes, the Cbf5 homolog (HVO_2493) is not essential in this archaeon. The Cbf5-deleted strain of H. volcanii completely lacks Ψ at positions 1940, 1942, 2605, and 2591 (Escherichia coli positions 1915, 1917, 2572, and 2586) of its 23S rRNA, and contains reduced steady-state levels of some box H/ACA RNAs. Archaeal Cbf5 is known to have tRNA Ψ55 synthase activity in vitro but we could not confirm this activity in vivo in H. volcanii. Conversely, the Pus10 (previously PsuX) homolog (HVO_1979), which can produce tRNA Ψ55, as well as Ψ54 in vitro, is shown here to be essential in H. volcanii, whereas the corresponding tRNA Ψ55 synthases, Pus4 and TruB, are not essential in yeast and E. coli, respectively. Finally, we demonstrate that HVO_1852, the TruA/Pus3 homolog, is responsible for the pseudouridylation of position 39 in H. volcanii tRNAs and that the corresponding gene is not essential.
Keywords
INTRODUCTION
Pseudouridine (Ψ), the post-transcriptional C5-ribosyl isomer of uridine, is one of the most common modifications identified in a wide range of RNA species and in all domains of life (Charette and Gray 2000; Grosjean 2009; Cantara et al. 2011). Ψ modifications are very abundant in tRNAs. In addition to the near ubiquitous Ψ55 found in tRNAs of Archaea, Bacteria, Eukarya, and most of the organelles, Ψ is found at several other positions of tRNA (Juhling et al. 2009). Ψ modifications are plentiful in rRNAs, particularly in eukaryotes with ∼100 identified in mammals and ∼50 found in Saccharomyces cerevisiae (Cantara et al. 2011). Ψ residues appear to be located specifically at functionally important positions (Ofengand et al. 2001b; Decatur and Fournier 2002; Karijolich and Yu 2010).
Although the biological role of many Ψ residues remains unknown, the modification is thought to be required for RNA structure and stability. Indeed, the ability of Ψ residues to form an additional hydrogen bond compared to uridine, coupled with the property of augmenting local RNA stacking, causes the localized phosphodiester structure to become more rigid, contributing to overall RNA structure stabilization (Davis 1995; Yarian et al. 1999). Structural studies have suggested that, while the presence of pseudouridine modifications does not affect the overall three-dimensional structure of tRNA (Harrington et al. 1993; Arnez and Steitz 1994), the local structure near the site of modification is affected (Auffinger and Westhof 1997). The presence of Ψ in the anticodon helps foster proper codon–anticodon interactions, which may increase translational accuracy, and the presence of Ψ at positions 38/39 of tRNA has a direct influence on codon reading and frameshifting (Zerfass and Beier 1992; Lecointe et al. 2002; Namy et al. 2005). Ψ modifications in rRNAs can affect both ribosome biogenesis and translation accuracy (Decatur and Fournier 2002; Baudin-Baillieu et al. 2009).
Six distinct pseudouridine synthase families have been identified to date on the basis of protein sequence similarity: TruA, TruB, TruD, RluA, RsuA, and Pus10 (Mueller and Ferre-D'Amare 2009). Predicting the substrate specificity of any given pseudouridine synthase is, however, difficult. Different proteins of the same family can produce Ψ at different positions (Hamma and Ferre-D'Amare 2006; Mueller and Ferre-D'Amare 2009). The annotation of Ψ modification genes is all the more difficult given that examples of nonorthologous displacements are frequent. In Escherichia coli and S. cerevisiae pseudouridylation of position 55 of tRNA is catalyzed by TruB and Pus4, respectively (Nurse et al. 1995; Becker et al. 1997), both members of Cluster of Orthologous Group (COG) 0130 (Tatusov et al. 2003). In Archaea however the unrelated Pus10 family of proteins belonging to COG 1258 seems to modify both positions 54 and 55 of tRNA (Roovers et al. 2006; Gurha and Gupta 2008).
Specific Ψ residues in eukaryal and archaeal rRNAs and in eukaryal snRNAs can also be introduced by box H/ACA RNPs. The H/ACA RNPs comprise four core proteins and a modification-specific guide RNA containing between one and three hairpins (Dennis and Omer 2005; Yu et al. 2005; Meier 2006; Karijolich and Yu 2008; Li 2008; Hamma and Ferre-D'Amare 2010; Kiss et al. 2010). Cbf5, a member of the TruB family (COG 0130), is the pseudouridine synthase in the RNP, which together with Nop10, L7Ae (Nhp2 in Eukarya), and Gar1 forms a site-specific complex. Box H/ACA RNA-guided in vitro pseudouridylation systems using recombinant Pyrococcus furiosus, Pyrococcus abyssi, and Methanocaldococcus jannaschii proteins have been established (Baker et al. 2005; Charpentier et al. 2005; Gurha et al. 2007; Muller et al. 2008). However, Cbf5, in addition to its RNA-guided activity, also produces Ψ at position 55 in tRNAs and in a 23S rRNA fragment in vitro in a guide RNA-independent manner. This guide RNA-independent activity of Cbf5 either requires or is enhanced by association with Gar1 and Nop10 (Roovers et al. 2006; Gurha et al. 2007; Muller et al. 2008).
In Eukarya the Cbf5 encoding gene is essential (Jiang et al. 1993; Giordano et al. 1999), which may be due to multiple roles of H/ACA RNPs in ribosome biogenesis, pre-mRNA splicing, and telomere maintenance (Meier 2005; Karijolich and Yu 2008). Indeed, Cbf5 (Nap57 in mice) was originally identified in S. cerevisiae as a low affinity centromere binding protein, which is essential for cell viability (Jiang et al. 1993; Meier and Blobel 1994; Heiss et al. 1998). Orthologs of cbf5 are essential in yeast and Drosophila melanogaster, and its disruption is embryonic lethal in mice (Jiang et al. 1993; Phillips et al. 1998; Giordano et al. 1999; He et al. 2002).
Most of the information on the role of Cbf5 and protein-alone Ψ synthases in Archaea is based on bioinformatic, biochemical, or structure-based data. The essentiality and function of these proteins in vivo is therefore still undetermined in this domain of life. We set out to determine the in vivo role of Cbf5 in Ψ synthesis as well as of other Ψ synthases using the model archaeon H. volcanii. Indeed, sequences of the nearly complete set of tRNAs with associated modifications have been determined in this archaeon (Gupta 1984, 1986) and the positions of Ψs in 23S rRNA can be inferred by comparison with other haloarchaea (Ofengand and Bakin 1997; Del Campo et al. 2005; Kirpekar et al. 2005 and http://people.biochem.umass.edu/fournierlab/3dmodmap/heneqlsu.php). H. volcanii 16S rRNA does not contain Ψ (Kowalak et al. 2000). Ψ synthases for most of these Ψ have also been predicted in the H. volcanii genome using a bioinformatic approach (Grosjean et al. 2008). In this work, we achieved the deletion of cbf5 (HVO_2493) (H. volcanii ORFs are as annotated in http://archaea.ucsc.edu and Hartman et al. 2010) and demonstrated that Cbf5 is responsible for the introduction of Ψ at multiple positions in the 23S rRNA, but for none of the Ψs at common positions of tRNAs. We also made efforts to create genomic-deletions of certain other Ψ synthases, to determine their role in producing these non-Cbf5-dependent Ψs in tRNAs. We show that pus10 (HVO_1979), the gene for tRNA Ψ54 and Ψ55 synthase (Gurha and Gupta 2008), is essential in H. volcanii. We also show that the H. volcanii TruA/Pus3 homolog (HVO_1852) is indeed responsible for the introduction of Ψ39 in tRNA. Finally we show that Cbf5 is needed for stabilization of box H/ACA RNAs that are predicted to guide Ψ formation in 23S rRNA.
RESULTS AND DISCUSSION
HVO_2493, encoding the Cbf5 homolog, is not essential in H. volcanii
The H. volcanii cbf5 homolog HVO_2493 was deleted using a published method (Allers et al. 2004), modified slightly to expedite the preliminary cloning steps (Blaby et al. 2010). Deletion of HVO_2493 was confirmed by PCR using two primer pairs, one designed to anneal outside the deleted fragment and the other within (Fig. 1B,C), and independently verified by Southern hybridization (Fig. 1D). These data indicate that cbf5 is dispensable in H. volcanii. However, as shown in Figure 1E, deletion of cbf5 caused a slight decrease in growth rate (from 0.03 h−1 in the wild-type [WT] strain to 0.02 h−1 in the mutant), although the final growth yield is not affected. To rescue the effects of the cbf5 deletion, a derivative of the Δcbf5 strain was created (Fig. 1A,D), which contained a plasmid-borne copy of cbf5. The growth rate was restored to WT levels when cbf5 was expressed in trans in the mutant strain (Fig. 1E). A well-established role of Ψ residues is to maintain tertiary structure. If the absence of Cbf5 influenced the stability of the rRNA molecule, the H. volcanii Δcbf5 strain could become more sensitive to altered salt concentrations or to temperature extremes. However, deletion of cbf5 did not lead to any obvious phenotype when analyzing growth on solid medium at 20, 30, 37, 44, and 50°C or with total salt concentrations ranging from 12% (1.812 M NaCl, 65.28 mM MgCl2, 63 mM MgSO4, and 41.64 mM KCl) to 25% (3.775 M NaCl, 136 mM MgCl2, 131.25 mM MgSO4, and 86.75 mM KCl) (compared to the standard YPC media with a final concentration of 18% salt; data not shown). It is presently unclear whether slower growth of the cbf5-deleted strain is due to the absence of Cbf5 itself or due to the loss of Cbf5-catalyzed modifications.
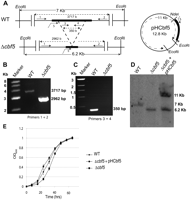
cbf5-deleted strain of H. volcanii is viable. Deletion of the cbf5 gene was confirmed by three independent methods. (A) Schematics to show HVO_2493 (thick lines) in the genomes of wild-type (WT) and cbf5-deleted (Δcbf5) strains and on plasmid pHCbf5. EcoRI and NdeI (in plasmid) sites are marked. Positions of primers 1–5 (Cbf5_Ext_Fwd, Cbf5_Ext_Rev, Cbf5_Int_Fwd, Cbf5_Int_Rew, and HVCBF5-F, respectively) are shown by arrows. The arrow direction indicates 5′ to 3′ direction of the primers. Segment between broken lines is deleted in the Δcbf5 strain. The His-tag at the C terminus of the Cbf5 protein is indicated by an asterisk. Approximate lengths of the segments are indicated but are not drawn to a scale. (B) PCR products using primers designed to anneal outside the gene (primers 1 and 2 in A) confirm a genomic rearrangement of correctly predicted sizes in WT and mutant strains. (C) PCR products using primers designed to anneal within the target gene (primers 3 and 4 in A) show the absence of the HVO_2493 segment in the mutant and its presence in the WT strain. (D) Southern hybridization is consistent with loss of the gene in mutant strain. Blots of EcoRI-digested DNAs of WT and Δcbf5 strains, and Δcbf5 strain transformed with pHCbf5 were probed with [5′-32P]-labeled HVCBF5-F primer. The predicted sizes of the hybridized fragments are indicated beside the panel. (E) Growth curves of WT, Δcbf5, and Δcbf5+pHCbf5 strains of H. volcanii are shown. OD600 was measured using a Bioscreen C apparatus. Each point is the mean of three independent cultures; error bars represent standard error. H26 and VDC2364 were used as WT and Δcbf5 strains.
The nonessentiality of cbf5 in H. volcanii is in stark contrast to eukaryotes, where the gene is essential and point mutations lead to pleiotropic effects. Point mutations in yeast cbf5 can abolish in vivo pseudouridylation of rRNA (Zebarjadian et al. 1999) and mutations in NAP57/dyskerin in mice decrease pseudouridylation of rRNA and assembly of box H/ACA RNP (Ruggero et al. 2003; Grozdanov et al. 2009). RNAi silencing of Trypanosoma brucei Cbf5 depleted box H/ACA RNAs, and caused defects in rRNA processing and trans-splicing (Barth et al. 2005). Partial loss-of-function mutations in the D. melanogaster ortholog minifly result in small size and developmental delay (Giordano et al. 1999). Mutations in the human ortholog dyskerin result in the disorder dyskeratosis congenita (Heiss et al. 1998; Knight et al. 1999; Mitchell et al. 1999). The number of Ψ residues produced by Cbf5 is much larger in eukaryotes than in Archaea. Therefore, the essentiality of cbf5 in complex eukaryotic systems may be due to the vital role of certain Cbf5-mediated Ψ residues in biological processes, e.g., translation, pre-mRNA splicing, telomere maintenance, etc. Alternatively Cbf5 may have some other functions in addition to pseudouridylation in eukaryotes that may be essential. At least in yeast the essential nature of Cbf5 is not due to its pseudouridylation activity, because cells containing a mutant Cbf5 with the catalytic aspartate mutated to alanine lack Ψ in their rRNA, but show a temperature-sensitive growth phenotype (Zebarjadian et al. 1999). Another possibility is a role of Cbf5 in maintaining essential H/ACA RNAs in eukaryotes. U17/E1/snR30 is a nonpseudouridylating H/ACA snoRNA that is essential for rRNA processing (and consequently cell growth) (Meier 2006). A Cbf5 depletion can destabilize this RNA. If the only activity of Cbf5 in “simpler” archaeal systems is to produce Ψ, then it might not be absolutely necessary under normal growth conditions.
Cbf5 is needed for pseudouridylation of the large subunit rRNA residues of H. volcanii
Ψ residues are found at four positions in the 23S rRNA of Halobacterium salinarium (Halobacterium halobium) (Ofengand and Bakin 1997) corresponding to positions 1940, 1942, 2591, and 2605 in H. volcanii. The 23S rRNA of Haloarcula marismortui contains only three Ψ (Del Campo et al. 2005; Kirpekar et al. 2005). Position 2607 of H. marismortui (corresponding to H. volcanii 2591) contains an unmodified U. In E. coli the corresponding residues in the 23S rRNA are m3Ψ1915, Ψ1917, A2572, and U2586 (http://people.biochem.umass.edu/fournierlab/3dmodmap/heneqlsu.php). Based on the correlation with E. coli and H. marismortui 23S rRNAs, H. volcanii residues 1940 and 1942 are in the loop of helix 69 of domain IV, residue 2605 is between helix 90 and 93 of domain V, and residue 2591 is in a bulge in the stem of helix 90 (http://people.biochem.umass.edu/fournierlab/3dmodmap/heneqlsu.php). We sought to determine which of the four H. volcanii positions contained Ψ, and which of these were Cbf5-mediated. Both CMCT and U-reaction based approaches were used to determine whether a Ψ or an unmodified U is present at each of these four positions in the WT strain. Furthermore, when Ψ is present at a specific position, we determined whether it was replaced by an unmodified U in the cbf5-deleted strain and, if so, whether it reverts to Ψ in the complemented strain and is thus Cbf5-mediated.
Figure 2A,B shows that positions 1940 and 1942 of 23S rRNA contain Ψ in the WT strain. However, the same positions contain unmodified U residues in the Δcbf5 strain, which reverts back to Ψ when cbf5 is added back in trans. These results suggest that production of Ψ at both positions 1940 and 1942 is Cbf5-mediated. Similar data presented in Figure 2C,D suggest that Cbf5 is also responsible for the formation of Ψ at position 2605. Presence of Ψ at these three positions is in agreement with the results of H. salinarium and H. marismortui (Ofengand and Bakin 1997; Del Campo et al. 2005; Kirpekar et al. 2005). These three Ψ modifications were predicted to be catalyzed by Cbf5 in an RNA-guided manner (Grosjean et al. 2008). The results presented here confirm that Cbf5, at least, is critical in the production of these three Ψ residues.
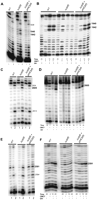
Pseudouridylation of H. volcanii 23S rRNA is Cbf5-mediated. (A) U-specific analyses to determine the modification status of U1940 and U1942 of 23S rRNA were done using primer HVLSUR1 and total RNAs of WT and Δcbf5 strains, and Δcbf5 strain transformed with pHCbf5. (Lanes 1,2) Primer extensions without and with U-specific reactions, respectively. Positions of certain U in 23S rRNA are indicated on the side; those in bold indicate Cbf5-mediated modification while that in italic indicates an unmodified U (used as indicator for the positions). (B) CMCT-primer extension analyses to determine the modification status of U1940 and U1942 of 23S rRNA were done using primer HVLSUR1 and total RNAs of WT and Δcbf5 strains, and Δcbf5 strain transformed with pHCbf5. Total RNAs were treated with (+) or without (−) CMCT for the indicated time (in minutes), followed by alkali (OH−) treatment (+) or no treatment (−). Positions of Cbf5-mediated modifications are indicated on the side. (C,D) Analyses similar to those in A and B, respectively, using primer HVLSUR2, to determine the modification status of U2605 of 23S rRNA. (E,F) Analyses similar to those in A and B, respectively, using primer HVLSUR3, to determine the modification status of U2591 of 23S rRNA.
Analysis for the presence of Ψ at position 2591 in 23S rRNA is less clear. Based on the data presented in Figure 2E,F, it appears that the formation of Ψ2591 is Cbf5-mediated as it is present only if the cbf5 gene is present in cis or trans. We believe that conversion of U to Ψ at this position is not 100% for the following reasons. Neither U nor Ψ should produce a strong stop in untreated samples (see lanes 1 in Fig. 2A,C). However, presence of stops in lanes 1 of WT and Δcbf5+pHCbf5 (Fig. 2E) suggests that position 2591 contains a residue other than U or Ψ, such as a secondary modification of Ψ (see below). On the other hand lanes 2 (U-reaction) of the same strains should not show stops unless there is an unmodified U (or an unknown modification of U that behaves like an unmodified U in aniline/hydrazine treatment) at position 2591. Similar incomplete conversion to Ψ has also been observed in certain H. volcanii tRNAs (Gupta 1984) as well as in other RNAs (discussed in Grosjean 2005). Recently it has been shown that pseudouridylation of certain residues of S. cerevisiae U2 snRNA can be conditionally induced in response to nutrient deprivation and heat shock (Wu et al. 2011). At present we cannot be certain whether 23S rRNA with and without Ψ2591 in our RNA preparations reflect a mixed population of rRNAs in each cell or a mixed population of cells, each containing only one type of rRNA. In the former case, it would raise questions as to whether each cell has two or more types of differentially modified ribosomes, each one representing different translation machinery or translating different groups of mRNAs.
Finally a secondary modification of Ψ seems to occur at position 2591. Untreated RNAs of WT and Δcbf5+pHCbf5 strains, but not of Δcbf5 strain, show a band (primer-extension stop) at position 2591 (Fig. 2E, lanes 1). Primer-extension stops with untreated RNAs do not appear to be due to the presence of a strong secondary structure in this region of the RNAs, as the untreated RNA of the Δcbf5 strain does not show this stop, and Ψ normally does not inhibit the extension. So far only four modifications of Ψ have been identified, 1-methylpseudouridine (m1Ψ) in Archaea, 1-methyl-3-(3-amino-3-carboxypropyl) pseudouridine (m1acp3Ψ) in Eukarya, 3-methylpseudouridine (m3Ψ) in Bacteria, and 2′-O-methylpseudouridine (Ψm) in both Archaea and Eukarya (http://rna-mdb.cas.albany.edu/RNAmods/ and http://modomics.genesilico.pl/modifications/). The first three are base modifications, whereas the last one is a ribose modification. These base modifications require U to Ψ conversion, before production of secondarily modified Ψ. It is difficult to conclude at this point whether the secondary modification of Ψ2591 in H. volcanii 23S rRNA is one of the three known base modifications of Ψ or is a new modification.
As mentioned above, H. salinarium also contains Ψ (Ψ 2590) at the equivalent position to H. volcanii Ψ2591 (Ofengand and Bakin 1997). It is possible that H. salinarium may also have this secondary modification of Ψ, since CMCT reactions alone may not be able to distinguish between Ψ and certain modified Ψ. Again as mentioned above, H. marismortui is not reported to contain Ψ at position 2607 (equivalent to H. volcanii 2591) (Del Campo et al. 2005; Kirpekar et al. 2005). However, Kirpekar's laboratory (Kirpekar et al. 2005) does see a medium strong stop at position 2607 of H. marismortui in their CMCT reactions but could not confirm the presence of Ψ at this position by MALDI-MS study. It is possible that, like H. volcanii, H. marismortui may also contain a subpopulation of 23S rRNAs that contain either a Ψ or a secondarily modified Ψ at their position 2607, while the rest contain unmodified U at that position.
Position 2619 in H. marismortui 23S rRNA has been shown to contain 3-methyluridine (m3U) (Kirpekar et al. 2005), which inhibits primer extension. Residues at its equivalent positions in E. coli and human 23S rRNAs are U2584 and m3U4500, respectively (http://people.biochem.umass.edu/fournierlab/3dmodmap/heneqlsu.php). Lack of this m3U modification in H. salinarium has been shown to confer sparsomycin resistance (Lazaro et al. 1996; Kirpekar et al. 2005). Its equivalent position in H. volcanii is 2603, which shows strong primer-extension stops across all lanes in all strains (Fig. 2C,D). Taken together, this suggests that H. volcanii also has m3U2603 in its 23S rRNA.
H. volcanii tRNA Ψ55 residues are produced even in the absence of Cbf5
Positions of Ψ residues in nearly all tRNAs of H. volcanii have been determined and compiled (Gupta 1984, 1986; Grosjean et al. 2008). However, determination of the presence or absence of these Ψ in native tRNAs by primer-extension-based methods is technically challenging. Strong secondary structure of tRNAs in general and presence of certain modified residues in single-stranded regions pause or halt reverse transcriptase reactions (Motorin et al. 2007). Furthermore, the presence of partial modifications at a given site (Gupta 1984) can make primer extension reactions difficult to interpret. Therefore, a combination of strategies was used to determine the effect of cbf5-deletion in production of Ψs at known positions of tRNAs.
Two panels in Figure 3A reflect nucleotide composition of total tRNA extracted from WT and Δcbf5 cells. The amount of Ψ appears to be the same in both strains. Most of the Ψ in the tRNA population comes from Ψ at positions 55 (in all tRNAs), and 13 and 39 (in several tRNAs) (Grosjean et al. 2008). Therefore, it appears that Ψ modifications at these three positions are not Cbf5-mediated.

cbf5 deletion does not significantly reduce the amount of Ψ in H. volcanii total tRNAs and at tRNA position 55. Nuclease P1 digests of uniformly labeled tRNAs were resolved by thin layer chromatography. pA, pC, pG, pU, pΨ, and pm1Ψ indicate 5′-phosphorylated A, C, G, U, Ψ, and m1Ψ, respectively. (A,B) Digests of total tRNA and tRNATrp, respectively, from WT (upper panels) and Δcbf5 strains (lower panels). Ratio of Ψ and m1Ψ is indicated in each panel.
Cbf5 either alone or in association with other H/ACA RNP core proteins has been shown to produce Ψ55 in a guide RNA-independent manner in vitro in full-size tRNAs or tRNA fragments containing TΨC stem–loop (Roovers et al. 2006; Gurha et al. 2007; Muller et al. 2007; Zhou et al. 2011). To test whether this reaction occurs in vivo, we analyzed H. volcanii tRNATrp. This tRNA contains only one Ψ (Ψ55) and one modified Ψ (m1Ψ54) (Gupta 1984). The Ψ/m1Ψ ratio of tRNATrp remains the same even after deletion of cbf5, suggesting that Cbf5 is not the primary enzyme producing tRNA Ψ55 (Fig. 3B) in vivo. As attempts to delete Pus10 were not successful (see below) the question whether Ψ55 might be modified in vivo by Cbf5, in absence of active Pus10, remains open.
Cbf5 belongs to the TruB/Pus4 family of Ψ synthases that produce tRNA Ψ55 in Bacteria/Eukarya. Apparently archaeal Cbf5 protein, although specialized for RNA-guided activity, seems to have retained its capacity to produce tRNA Ψ55, though it does not seem to be its normal function. It is possible that there was a common ancestral mechanism for recognition of tRNA for Ψ55 modification by TruB/Pus4 and archaeal Cbf5. A similar example exists for archaeal and eukaryal tRNA splicing endonucleases (Fabbri et al. 1998; Fruscoloni et al. 2001), where the enzymes are homologous but have evolved to use different features of the pre-tRNAs to determine their cleavage sites (which are generally symmetric in Archaea and asymmetric in Eukarya). However, the eukaryal enzyme has retained the ability to use archaeal recognition signals. Alternatively, archaeal Cbf5 may produce Ψ in rRNAs in vivo in both RNA-guided and guide RNA-independent manners, the latter reaction recognizing a structure somewhat similar to the TΨC stem–loop of tRNA. Structures similar to this stem–loop are indeed present in rRNAs (Nagaswamy and Fox 2002; Krasilnikov and Mondragon 2003), and U2603 in a fragment of P. abyssi 23S rRNA has been shown to be converted by Cbf5 along with other H/ACA core proteins in a guide RNA-independent manner (Muller et al. 2008). The sequence surrounding this U2603 can indeed be folded into a stem–loop similar to the TΨC stem–loop of tRNA.
Pus10, a tRNA Ψ55 and Ψ54 synthase, is essential in H. volcanii
Another archaeal protein, Pus10, has been shown to produce Ψ54 and Ψ55 in tRNA in vitro and Ψ55 in heterologous in vivo conditions (Roovers et al. 2006; Gurha and Gupta 2008). Despite several attempts, we failed to delete the H. volcanii pus10 homolog, HVO_1979, using our standard protocol. This led us to postulate that the gene might be essential. To address this, the gene was placed under control of the inducible Ptna promoter. A strain carrying the chromosomal HVO_1979 deletion could then be constructed in the presence of the gene in trans as described in the Methods section (Fig. 4). This strain required tryptophan for growth, hence providing strong evidence for pus10 essentiality in H. volcanii (Fig. 4). The in vitro data that showed Pus10 can introduce Ψ residues at positions 54 and 55 in tRNA combined with evidence presented above that Ψ55 is present in tRNAs of cbf5-deleted strains suggest that Pus10 is primarily responsible for the production of Ψ55 and Ψ54 in cellular tRNAs.
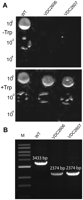
pus10 is essential in H. volcanii. pus10 (HVO_1979) could only be deleted from the chromosome when expressed in trans. (A) H26 (WT) and two independent isolates VDC2606 and VDC2607 (ΔHVO_1979 with plasmid-borne copy of HVO_1979 under Ptna control) were serially diluted from 100 to 10−4 and spotted onto Hv-Min media plates, plus and minus 500 μM tryptophan. Significant growth of the mutant is only observed in the presence of tryptophan. (B) Deletion of the chromosomal copy of HVO_1979 was further verified by PCR using primers Pus10_Ext_Fwd and Pus10_Ext_Rev. Predicted sizes of the amplicon are indicated above each band.
The essentiality of pus10 was unexpected as genes responsible for the insertion of Ψ55 in the other domains of life, at least truB and pus4, are dispensable in E. coli and yeast, respectively (Becker et al. 1997; Gutgsell et al. 2000). The strains lacking truB and pus4 are viable and have unmodified U55 in the tRNAs. This suggests that the essential nature of Pus10 in H. volcanii may be either due to its role in tRNA Ψ54 synthesis or in Ψ production at a presently unidentified position in other cellular RNAs. Alternatively Pus10 may be essential because of a function not related to pseudouridylation such as a possible chaperone function during tRNA maturation or being part of a multiprotein complex having some nonpseudouridylation function. Furthermore, Pus10 might not be essential in other Archaea. The in vitro Ψ54 activity of Pus10 from M. jannaschii is more robust than of Pus10 from P. furiosus, which may be a reflection of their in vivo functions (Gurha and Gupta 2008). Indeed, Thermococcales seem to have mostly 5-methyluridine (m5U or rT) as 5-methyl-2-thiouridine (m5s2U or s2T) in tRNA and few Ψ54 (or m1Ψ54) (Edmonds et al. 1991; Kowalak et al. 1994; Constantinesco et al. 1999; Urbonavicius et al. 2008). If this were the case, then it would be interesting to test if pus10 is essential in Pyrococcus sp.
Cbf5 is not needed for most of the tRNA Ψ residues of H. volcanii, but TruA/Pus3 homolog is needed for Ψ39
We determined the Ψ status at certain other positions of tRNAs in Cbf5-deleted strains of H. volcanii, in addition to the Ψ55 mentioned above, by primer extension following U-specific and CMCT reactions with RNAs of WT and Δcbf5 strains using elongator tRNAMet-specific primers (Fig. 5). Both strains show the presence of Ψ13 and Ψ22 in tRNAMet (Fig. 5A,B), suggesting that neither the formation of Ψ13 nor Ψ22 are Cbf5-mediated. Probably Ψ13 is produced by TruD/Pus7 because a homolog is present in H. volcanii (Grosjean et al. 2008), and P. abyssi Pus7 can produce Ψ13 in vitro (Muller et al. 2009).
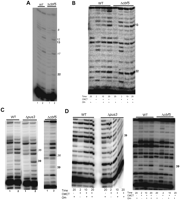
H. volcanii tRNA Ψ13, Ψ22, and Ψ39 are not Cbf5-mediated and Ψ39 is TruA/Pus3-mediated. (A) U-specific analyses to determine the modification status of U13 and U22 of elongator tRNAMet were done using primer HVMETCR2 and total tRNAs of WT and Δcbf5 strains. (Lanes 1,2) Primer extensions without and with U-specific reactions, respectively. Positions of certain Us and Ψ13 and Ψ22 in tRNA are indicated on the side. (B) CMCT-primer extension analyses to determine the modification status of U13 and U22 of tRNAMet were done using primer HVMETCR2 and total tRNAs of WT and Δcbf5 strains. Total tRNAs were treated with (+) or without (−) CMCT for the indicated time (in minutes), followed by alkali (OH−) treatment (+) or no treatment (−). Positions of Ψ13 and Ψ22 are indicated on the side. (C,D) Analysis similar to those in A and B, respectively, using primer VM5/6R, to determine the modification status of U39 of tRNAMet in WT (H26), Δpus3 (VDC2465), and Δcbf5 (VDC2364) strains.
Similar analysis using CMCT reactions suggests that Ψ39 in elongator tRNAMet is also not Cbf5-mediated (Fig. 5C,D) thus is not guide RNA dependent, whereas 2′-O-methylation of U at the same position 39 in tRNATrp of H. volcanii is guide RNA dependent (Clouet d'Orval et al. 2001; Singh et al. 2004). Ψ39 in tRNAs of Bacteria and Eukarya is produced by TruA and Pus3, respectively. Therefore, we deleted the gene encoding their H. volcanii homolog, HVO_1852 (data not shown). Primer extension following U-specific and CMCT reactions using WT and ΔHVO_1852 strains (Fig. 5C,D) shows that indeed as for Bacteria and Eukarya the TruA/Pus3 homolog in H. volcanii catalyzes the formation of Ψ39 in tRNA, further confirming that it is not Cbf5-mediated. Furthermore, these results show that pus3 in H. volcanii is not essential.
In addition to Ψ at positions 13, 39, and 55 (and m1Ψ54), Ψ is also present at four other positions in H. volcanii tRNAs and only in one tRNA for each of these additional positions. These are Ψ22 in elongator tRNAMet, Ψ28 in tRNAIle, Ψ38 in tRNAPro, and Ψ52 in tRNALys, the last three being only in the major isoacceptor for these amino acids (Gupta 1984; Grosjean et al. 2008). While we have demonstrated above that Ψ22 is not Cbf5-mediated, we were unable to obtain clear evidence as to whether Cbf5 has any role in the production of Ψ28. We did not attempt to determine Ψ38 and Ψ52 production in this analysis, because both of these show partial modification of tRNAs (Gupta 1984). The ratio of U vs. Ψ can vary from a tRNA preparation to another, which can affect the analysis of the data. However, based on the activities of E. coli TruA and S. cerevisiae Pus3 (Kammen et al. 1988; Nurse et al. 1995; Lecointe et al. 2002), we can assume that Ψ38 is also produced by their homolog in H. volcanii. At present it is difficult to predict any Ψ synthase for the other three positions (22, 28, and 52) with any certainty.
cbf5 deletion reduces the amounts of guide box H/ACA RNAs in H. volcanii
Two box H/ACA guide RNAs have been proposed for production of Ψ in H. volcanii 23 rRNA: one for U2605 and another one for both U1940 and U1942 (Grosjean et al. 2008). Both of these RNAs are present in the cell as shown by Northern hybridizations of WT RNA using appropriate probes (Fig. 6B). Interestingly, signals for the probes were greatly reduced in the Δcbf5 strain, but the signals return to WT levels when the mutant strain is supplemented with a plasmid-borne copy of cbf5. This suggests that, in the absence of Cbf5, H/ACA RNAs are destabilized and get degraded. This would be expected if these RNAs normally formed H/ACA RNP complexes with Cbf5 and other proteins and were thereafter protected against cellular RNA nucleases. Similar situations are observed in yeast, where genetic depletion of Cbf5 leads to depletion of all box H/ACA snoRNAs (Lafontaine et al. 1998) and in T. brucei, where RNAi-mediated depletion of Cbf5 destabilized box H/ACA snoRNAs (Barth et al. 2005). Our results in Figures 2A–D and 6B together suggest that Ψ1940, 1942, and 2605 in 23S rRNA are produced by H/ACA RNPs. This is confirmed for the position 2605 by our in vitro pseudouridylation assays using appropriate guide RNAs, and a 23S rRNA fragment containing U2605 and recombinant M. jannaschii proteins (M Majumder and R Gupta, unpubl.).
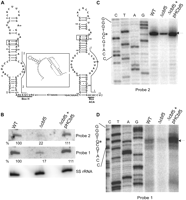
Levels of a double stem–loop box H/ACA guide RNA are reduced in the absence of Cbf5. (A) Primary sequence and predicted secondary structures of two H/ACA guide RNAs proposed (Grosjean et al. 2008) to be responsible for guiding pseudouridylation of U2605 and U1940/U1942 of H. volcanii 23S rRNA. The 59-base region that separates the two RNAs in the genome can potentially form a pseudoknot (inset). Pseudoknot is drawn according to pknotsRG-mfe (http://bibiserv.techfak.uni-bielefeld.de/pknotsrg/). Conserved box H and box ACA are underlined. G•A pairs of K-turns are boxed. Positions for probe 1 (HV1940/42HAR) and probe 2 (HV2605HAR) are indicated by lines along the sequences. (B) Northern blot analysis of total RNA isolated from WT and Δcbf5 strains, and Δcbf5 transformed with pHCbf5. Blot was hybridized using probes 1 and 2 (shown in A). Both probes hybridized to an ∼200-base RNA. 5S rRNA-specific probe was used as a loading control. Quantity of RNA in each hybridized band is indicated. (C) Primer extension and sequencing reactions using probe 2 to determine 5′ ends of the RNAs. Equal amounts of total RNAs from WT and Δcbf5 strains and Δcbf5 transformed with pHCbf5 were used in the reactions. Sequence in the region of primer stop is indicated on the side. Asterisked G indicates the 5′ end of the RNA. (D) Reactions similar to C, using probe 1.
A double stem–loop box H/ACA RNA in H. volcanii
Both probes used for Northern analyses seemed to hybridize to one RNA of ∼200 bases (Fig. 6B). The two sRNA sequences are located near each other in the H. volcanii genome (http://archaea.ucsc.edu) and are separated by a 59-base spacer region between the two stem–loops. Therefore, we used the same two probes to determine the 5′ ends of the two RNAs. As seen in Figure 6C,D, both RNAs have the same 5′ end, suggesting that both RNAs are part of the same molecule. Reduced signals in the Δcbf5 strain (Fig. 6C,D) again confirm the reduced level of this box H/ACA RNA in the mutant strain and the recovery to original levels when the strain is supplemented with a plasmid-borne copy of cbf5. Archaeal H/ACA RNAs mostly have either one or three stem–loops, though two stem–loops have also been observed, albeit rarely (Dennis and Omer 2005; Muller et al. 2008). This H. volcanii RNA is a rare case of archaeal H/ACA RNA containing two stem–loops. Normally spacer regions between the stem–loops of archaeal H/ACA RNAs are very short (Grosjean et al. 2008; Muller et al. 2008). However, this double stem–loop RNA has a spacer of 59 bases (or 56 bases excluding ACA of the stem–loop on the 5′ side). The spacer length between homologs of these two stem–loops among haloarchaea varies from 21 to 142 bases, but other Archaea have smaller spacers from 0 to 5 bases (Grosjean et al. 2008). This spacer of H. volcanii potentially can form a pseudoknot structure (inset in Fig. 6A). Interestingly, this spacer of all haloarchaeal species can also be folded into a pseudoknot structure (data not shown). Generally pseudoknots have some kind of regulatory role but it is not yet known if this structure between the two H/ACA RNAs forms in vivo and whether it has any role there.
Conclusions
In conclusion, we present here the analysis and confirmation of several proteins previously predicted to be involved in pseudouridylation of ribonucleic acids in H. volcanii. We demonstrate that Cbf5 is responsible for making Ψ at multiple positions of the large subunit of rRNA. Contrary to suggestions made by previous in vitro based assays Cbf5 does not appear to be involved in tRNA modification. Furthermore, while essential in yeast, cbf5 is dispensable in H. volcanii, although its deletion does result in a slight growth defect. However, another protein suspected to be involved in pseudouridylation of at least U55 of tRNAs, Pus10, is necessary for viability. Finally we demonstrate that HVO_1852, the TruA homolog, is responsible for Ψ39 in tRNA.
It is important to note that the results reported here might be specific to halophilic Archaea. Compared to E. coli or yeast, H. volcanii clearly contains a lower number of RNA modifications (both in tRNA and rRNA) and comparative genomics analysis suggests the number of RNA modifications in certain halophilic Archaea could be reduced even more. Indeed, as far as tRNAs are concerned, H. volcanii has already lost the tyw1 signature gene for the imG-14 modification (Waas et al. 2005) that is present in H. salinarium (de Crécy-Lagard et al. 2010). Haloquadratum walsbyi, which lives in an environment with a much more stable salinity level than H. volcanii (Burns et al. 2007), has lost not only the tyw1 gene like H. volcanii but also the tgtA gene involved in the formation of archaeosine (Watanabe et al. 1997) and the gene encoding COG 1444 that has been shown to be involved in ac4C formation (El Yacoubi et al. 2009). A possibility is that high salt concentration can replace specific modifications in these organisms (discussed in Grosjean et al. 2008). It is therefore crucial to repeat the experiments reported here with nonhalophilic Archaea before any generalization is made.
MATERIALS AND METHODS
Strains, plasmids, oligonucleotides, media, and transformation procedures
All strains, plasmids, and oligonucleotides used in this study are detailed in Supplemental Tables S1, S2, and S3, respectively. H. volcanii H26 was received from Julie Maupin-Furlow as well as from Thorsten Allers. Plasmids pJAM202 (Kaczowka and Maupin-Furlow 2003) and pMDS132, a derivative of pMLH32 (Holmes and Dyall-Smith 2000), were gifts from Julie Maupin-Furlow and Michael Dyall-Smith, respectively. H26 and VDC2364 (prepared in this work) were used in this study as WT and Δcbf5 strains of H. volcanii. E. coli was routinely grown in LB (Fisher) or LB agar (Fisher) at 37°C, supplemented when required with ampicillin (100 μg/mL), isopropyl β-D-1-thiogalactopyranoside (0.2 mM), and bromo-chloro-indolyl-galactopyranoside (40 μg/mL). H. volcanii cells were routinely grown at 42°C–44°C (unless specified) in Hv-YPC, Hv-CA, and Hv-Min as described in the HaloHandbook (http://www.haloarchaea.com/resources/halohandbook/Halohandbook_2008_v7.pdf) or in a standard medium as described previously (Gupta 1984). Transformations of E. coli were performed by standard protocols (Sambrook and Russell 2001) or as described by the manufacturer (Invitrogen). Transformation of H. volcanii was performed as described in the HaloHandbook using the “standard PEG-mediated transformation of haloarchaea” protocol. Briefly, spheroplasts were made by adding 100 μL 0.5 M EDTA (pH 8.0) to 1 mL concentrated cells, incubating at room temperature for 10 min; 2 μg plasmid DNA was then added to 100 μL spheroplasts and incubated for 5 min; 100 μL 60% (v/v) PEG600 was added and mixed gently, and the mixture incubated at room temperature for 30 min. Cells were recovered by adding recovery solution, as described in the HaloHandbook, and incubated for 4 h at 42°C–44°C before plating onto the appropriate media. Phenotypes (growth rate, and growth at different salt concentrations and temperature) were tested as described previously (Blaby et al. 2010).
Deletion strain and plasmid construction
Plasmids to delete H. volcanii genes were constructed as described previously (Blaby et al. 2010). Briefly, regions of ∼600 bp upstream of and downstream from the gene to be deleted, along with a small portion of the gene at both ends to avoid affecting neighboring genes, were PCR-amplified from H. volcanii genomic DNA and recombined between the XhoI and EcoRI sites of pTA131 using InFusion (Clontech); 0.2 μL of the reaction mixture was transformed to TOP10 chemically competent cells (Invitrogen) and plated onto LB plus ampicillin (100 μg/mL), isopropyl β-D-1-thiogalactopyranoside (0.2 mM), and bromo-chloro-indolyl-galactopyranoside (40 μg/mL). Clones were screened by PCR using M13 primers (which anneal on either side of the multiple cloning site of pTA131) using 5′ Master Mix (Eppendorf) as per the directions, and confirmed by sequencing using the UF Sequencing facility.
The plasmid generated to disrupt HVO_2493 (pIKB219) was constructed as described above using oligonucleotide pairs HVO2493_N_IfKO_Fwd, HVO2493_N_IfKO_Rev, HVO2493_C_IfKO_Fwd, and HVO2493_C_IfKO_Rev. The plasmid constructed to delete HVO_1979 (pIKB180) was made using oligonucleotides HVO_1979_N_IfKO_Fwd, HVO_1979_N_IfKO_Rev, HVO_1979_C_IfKO_Fwd, and HVO_1979_C_IfKO_Rev. The plasmid used to delete HVO_1852 (pIKB199) was made using HVO_1852_N_IfKO_Fwd, HVO_1852_N_IfKO_Rev, HVO_1852_C_IfKO_Fwd, and HVO_1852_C_IfKO_Rev.
Once obtained, deletion plasmids were passed through a dam− E. coli (INV110; Invitrogen), transformed into H. volcanii H26 (or derivatives) as described above. Deletion of the targeted locus was selected in a two-step process as described previously (Allers et al. 2004). Briefly, insertion of the deletion plasmid by a single crossover event was selected for by ability to grow on Hv-CA (i.e., in the absence of uracil). Subsequent excision of the integrated plasmid and target gene by a second recombination event was selected for by plating onto Hv-CA supplemented with uracil (10 μg/mL) and 5-fluoroorotic acid (5-FOA; 50 μg/mL). Gene deletion candidates were screened using a PCR-based method as follows. One pair of primers (denoted by the suffix _Ext_Fwd and _Ext_Rev) was designed to anneal within the flanking regions of the gene to be deleted, and the amplicon compared to WT and predicted size. To confirm loss of the gene, a second pair of primers (suffixed _Int_Fwd and _Int_Rev) was designed to anneal within the gene to be deleted. Gene deletions were verified by amplification of the fragment in the WT, but inability in mutant strains.
Other plasmid and strain constructions
pHCbf5 was generated by replacing ∼420 bp NdeI-EcoRI fragment of pMDS132 with an ∼890 bp PCR fragment produced by amplifying HVO_2493 using HVCBF5ND-F2 and HVCBF5-R2 primers. An amplified gene contains six His codons just before the stop codon. The reaction mixture was used to transform DH5α cells. Plasmids were verified by PCR, restriction digestion, and sequencing, and then passed through E. coli GM2929 and transformed into H. volcanii strain VDC2364 (Δcbf5 strain). Transformants were selected for by plating onto Hv-YPC containing novobiocin (0.8 μg/mL).
A fragment from the H. volcanii genome containing the Ptna promoter was PCR-amplified using primers Ptna_5 and rev-Ptna, and inserted between the BlpI and XbaI sites of pTA131, to generate pPT002. pus10 was subsequently PCR-amplified using primers Pus10_tna_Nde_Fwd and Pus10_tna_Rev from H. volcanii genomic DNA and inserted between the NdeI and EcoRV sites of pPT002 to generate pIKB487. Subsequent deletion of the chromosomal copy of pus10 was performed as described by Allers and colleagues (Allers et al. 2004; Large et al. 2007) using pIKB180 to disrupt the gene in the strain transformed with pIKB487.
Mapping the sites of Ψ in RNAs
Presence of Ψ in RNA was analyzed by the 1-cyclohexyl-3-(2-morpholinoethyl) carbodiimide metho-p-toluenesulfonate (CMCT) modification technique (Ofengand et al. 2001a; Motorin et al. 2007). Briefly, 20 μg total RNA or 10 μg gel-purified tRNA was treated with CMCT for 2, 10, and 20 min each at 37°C, while an untreated sample was simply incubated at 37°C for 20 min as control. After precipitation of the RNA, alkali treatment was done for 3 h at 37°C for all samples except for the ones treated with CMCT for 2 min. Again after precipitation, the RNA was used for primer extension using [5′-32P]-labeled primers that hybridized close to and on the 3′ side of the position to be mapped for Ψ. M-MLV reverse transcriptase (Promega) was used for primer extension according to the manufacturer's instructions. The extension stops at one residue before the CMCT modified Ψ. A dark band in CMCT followed by alkali treatment lanes, with an increased intensity in the 20 min treatment lane, indicates the presence of Ψ at that position. The position of the band in the gel that corresponds to Ψ is determined by its distance from the end of the primer and sometimes the presence of darker bands for U and G in 2 min CMCT but no alkali treatment lane, and correlation with the known sequence of the RNA.
U-specific sequencing reactions (Peattie 1979; Gupta 1984) followed by primer extension were used to determine the positions of U (absence of Ψ) in the RNAs. Briefly, 20 μg total RNA or 10 μg gel-purified tRNA was treated with 50% hydrazine for 20 min on ice and precipitated. The RNA was then treated with acidified aniline at 60°C for 20 min. Precipitated RNA was then used for primer extension as above for the CMCT reaction. Here also the position of the primer extension band in the gel is one residue before the expected U. A dark band in the hydrazine-aniline treated lane but not in the untreated lane indicates the presence of an unmodified U at that position. A correlation between the position of the dark band in the gel relative to the end of the primer and positions of unmodified Us in the known sequence of the RNA determines the position of Ψ.
Quantitation of Ψ residues in tRNAs
Uniformly labeled total tRNA was prepared as described before (Gupta 1984; Joardar et al. 2008). When needed, uniformly labeled tRNATrp was isolated from this total tRNA according to previously published procedures (Joardar et al. 2008). Labeled tRNAs were digested with nuclease P1 and digests were resolved by two-dimensional thin-layer chromatography on cellulose plates (EM Science) using isobutyric acid/0.5 N NH4OH (5:3, v/v) in the first dimension and isopropanol/HCl/H2O (70:15:15, v/v/v) in the second dimension (Gupta 1984). Radioactivity in the plates was revealed and quantified by phosphorimaging. The amount of Ψ per tRNA was determined by the ratio of radioactivities in Ψ vs. m1Ψ nucleotide spots. The reasons for using m1Ψ spot for comparison are because nearly all H. volcanii tRNAs have one m1Ψ residue per tRNA (Gupta 1984, 1986), Ψ for this m1Ψ is produced by Pus10 (Gurha and Gupta 2008) not by Cbf5, and this nucleotide is well separated in our chromatography system.
Southern and Northern hybridizations, and mapping the 5′ ends of guide RNAs
The genomic deletion of cbf5 was checked by Southern hybridization of EcoRI-digests of DNA using standard procedures (Sambrook and Russell 2001). The blots were hybridized with [5′-32P]-labeled HVCBF5-F primer. For Northern analyses, total RNA was isolated from the cells using TRI reagent (Molecular Research Center, Inc). The RNA was separated by a 6% denaturing PAGE. The blots were prepared and hybridized to [5′-32P]-labeled probes according to standard procedures (Sambrook and Russell 2001). The blot was stripped (incubated in 10 mM Tris-Cl pH 7.4, 0.2% SDS at 70°C for 2 h) after hybridization for use with a second probe. Radioactivity was detected and quantified (when needed) by phosphorimaging. The amount of H/ACA RNA in Northern blots was quantitated by using a band for 5S rRNA in each lane as a loading control and normalizing each H/ACA RNA band in that lane against these controls. The amount of RNA in WT is considered 100% in each case. To determine 5′ ends of the guide RNAs the primers used in Northern analyses were 5′-32P labeled and used with total RNA for extension reactions.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
H. volcanii H26 was received from Julie Maupin-Furlow (University of Florida) as well as from Thorsten Allers (University of Nottingham, UK). Plasmids pJAM202 and pMDS132 were gifts from Julie Maupin-Furlow and Michael Dyall-Smith (Charles Sturt University, Australia), respectively. We thank Wayne Decatur for the help in correlating the positions of Ψ in H. volcanii with those of other haloarchaea and E. coli. We thank Patrick Thiaville for the construction of pTA002. H.G. holds a CNRS position of Emeritus Scientist at the University Paris-South in Orsay-France. This work was funded in part by National Science Foundation grant number MCB-05169448 to V.d.C.-L. and by NIH grant number GM055945 to R.G.
Footnotes
-
↵6 Corresponding authors.
E-mail rgupta{at}siumed.edu.
E-mail vcrecy{at}ufl.edu.
-
Article published online ahead of print. Article and publication date are at http://www.rnajournal.org/cgi/doi/10.1261/rna.2712811.
- Received March 9, 2011.
- Accepted April 15, 2011.
- Copyright © 2011 RNA Society








