
Functional consequences of T-stem mutations in E. coli tRNAThrUGU in vitro and in vivo
Abstract
The binding affinities between Escherichia coli EF-Tu and 34 single and double base-pair changes in the T stem of E. coli tRNAThrUGU were compared with similar data obtained previously for several aa-tRNAs binding to Thermus thermophilus EF-Tu. With a single exception, the two proteins bound to mutations in three T-stem base pairs in a quantitatively identical manner. However, tRNAThr differs from other tRNAs by also using its rare A52–C62 pair as a negative specificity determinant. Using a plasmid-based tRNA gene replacement strategy, we show that many of the tRNAThrUGU T-stem changes are either unable to support growth of E. coli or are less effective than the wild-type sequence. Since the inviable T-stem sequences are often present in other E. coli tRNAs, it appears that T-stem sequences in each tRNA body have evolved to optimize function in a different way. Although mutations of tRNAThr can substantially increase or decrease its affinity to EF-Tu, the observed affinities do not correlate with the growth phenotype of the mutations in any simple way. This may either reflect the different conditions used in the two assays or indicate that the T-stem mutants affect another step in the translation mechanism.
Keywords
INTRODUCTION
The GTP form of bacterial elongation factor Tu (EF-Tu•GTP) binds aminoacyl-tRNAs (aa-tRNAs), and the resulting ternary complex subsequently participates in the multistep decoding reaction that ensures delivery of the correct aa-tRNA to the ribosomal A site (Rodnina and Wintermeyer 2001; Marshall et al. 2008). Several cocrystal structures reveal that the protein contacts the aa-tRNA over a large interface that includes the esterified amino acid and the acceptor and T helices of tRNA (Nissen et al. 1995, 1999). Most of these contacts persist until the aa-tRNA is released by EF-Tu and accommodates into the A site (Schmeing et al. 2009). Experiments with misacylated tRNAs reveal that the identities of both the esterified amino acid and the tRNA body contribute to the overall free energy of formation of the complex (LaRiviere et al. 2001; Asahara and Uhlenbeck 2002; Dale et al. 2004). This has allowed tRNA sequences to evolve to compensate for the differing thermodynamic contributions of the amino acid such that the overall binding free energy of all aa-tRNAs is similar. Biochemical experiments using mutations of the base pairs in the acceptor and T helices of yeast tRNAPhe revealed that the sequence of the 49–65, 50–64, and 51–63 base pairs in the T helix were largely responsible for the sequence-specific binding of this tRNA to Thermus thermophilus EF-Tu (Sanderson and Uhlenbeck 2007a; Schrader et al. 2009). When these three base pairs were transplanted from several Escherichia coli tRNAs into tRNAPhe, the resulting chimeras bound EF-Tu with an affinity similar to that of the parental tRNAs. This suggested that different combinations of these three base pairs could explain the different affinities of all aa-tRNAs to EF-Tu (Schrader et al. 2011a). Although mutagenesis of T. thermophilus EF-Tu confirmed that the three amino acids that directly contact the three base pairs contribute to the sequence specificity, at least two additional amino acids contribute to sequence specificity by an indirect mechanism (Sanderson and Uhlenbeck 2007b,c). Thus, the sequence specificity of EF-Tu is not completely understood and may differ somewhat among aa-tRNAs.
The goal of this paper is to test whether mutations in the T stem that alter the affinity of an aa-tRNA to EF-Tu can affect tRNA function in living E. coli cells. To do this, a plasmid shuffling strategy was used to introduce mutations of E. coli tRNAThrUGU into a strain where the single chromosomal copy of the gene was disrupted (Saks et al. 1998). Such a gene replacement approach permits an evaluation of the overall ability of a given mutant tRNAThrUGU to decode cellular ACA codons. In order to interpret the in vivo data, it was also necessary to measure the binding affinities of different T-stem variants of E. coli tRNAThrUGU with E. coli EF-Tu to determine whether the sequence specificity was similar to that previously found for yeast tRNAPhe with T. thermophilus EF-Tu. Although the cocrystal structures suggest that E. coli and T. thermophilus EF-Tu contact aa-tRNAs in a very similar way and tRNAPhe and tRNAThr are in the same structural class, differences in either the protein or tRNA sequences may affect how sequence-specific recognition is achieved.
RESULTS
Design and EF-Tu binding properties of E. coli tRNAThr variants
The wild-type sequence of E. coli tRNAThrUGU used for mutagenesis in these experiments is shown in Figure 1. The tRNAs are cloned into placZACA, a low-copy plasmid with a p15a replicon and a kanamycin-resistance gene. This plasmid also has a multiple cloning site that is flanked by a tufB promoter and an rrnC terminator so that tRNAs can be expressed in E. coli (W Liu and M Saks, unpubl.). Each tRNA gene is cloned behind a T7 RNA polymerase promoter and includes a BstN I site to permit in vitro run-off transcription so that it can be used for both in vivo and in vitro analyses. Twenty-seven variants of tRNAThrUGU containing single and multiple substitutions of the 49–65, 50–64, and 51–63 base pairs in the T stem were cloned into placZACA (Table 1). The mutations were expected to show a broad range of affinities to EF-Tu based upon previous experiments measuring the binding of T-stem mutants of yeast tRNAPhe and E. coli tRNALeu with T. thermophilus EF-Tu (Sanderson and Uhlenbeck 2007a; Schrader et al. 2009). After in vitro transcription and purification, both the wild-type and all of the variant tRNAThrUGU transcripts were found to aminoacylate to a similar high level (>70%) when high concentrations of ThrRS were used, indicating that the tRNAs had not grossly misfolded. In addition, initial rates of aminoacylation at low enzyme concentration were measured for several of the inviable T-stem mutants (see below) and found to have similar initial rates to the wild-type tRNAThr (data not shown). These results were expected since both the X-ray cocrystal structure and the biochemical experiments show that E. coli ThrRS does not interact with the T stem, but requires proper placement of identity residues in the anticodon and near the 3′ terminus (Hasegawa et al. 1992; Sankaranarayanan et al. 1999).
Dissociation rates of tRNAThrUGU variants from E. coli EF-Tu
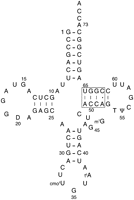
E. coli tRNAThrUGU with a box enclosing the four T-stem base pairs that were mutated in this work. The expected (Czerwoniec et al. 2009; Björk 1995) post-transcriptional modifications are as follows: D, dihydrouridine; T, 5-methyluridine; Ψ, pseudouridine; m7G, 7-methylguanosine, cmo5U, uridine 5-oxyacetic acid; t6A, N6-threonylcarbamoyladenosine.
The dissociation rates of the wild-type and variant [3H]Thr-tRNAThr transcripts from E. coli EF-Tu•GTP were determined using a ribonuclease protection assay (Louie and Jurnak 1985; Vorstenbosch et al. 2000). The buffer conditions and temperature were chosen to allow comparison with previous experiments and to ensure that the dissociation rates were slow enough to permit manual pipetting. Examples of dissociation rate data for wild-type and two variants are shown in Figure 2A and the mean koff values and the standard errors of the mean for all the variants are summarized in Table 1. In each case, sufficient E. coli EF-Tu•GTP was present to ensure virtually complete binding of the aa-tRNA and the complex dissociated with a single exponential rate. The koff values were converted to ΔG° using a constant kon = 0.11 μM−1sec−1, a value that was experimentally determined for E. coli (S Chapman, J Schrader, and O Uhlenbeck, unpubl.) and T. thermophilus EF-Tu (Schrader et al. 2009) using several different aa-tRNAs. The wild-type E. coli tRNAThrUGU transcript has a koff of 24 × 10−4 sec−1, which is similar to the value found for numerous other aa-tRNAs binding to both E. coli and T. thermophilus EF-Tu under similar conditions (Louie and Jurnak 1985; Asahara and Uhlenbeck 2005; Schrader et al. 2011b).

Binding of T-stem variants of unmodified E. coli Thr-tRNAThrUGU to E. coli EF-Tu•GTP. (A) Individual determinations of the dissociation rates of WT (•), 18 (▴), and 5 (▪) in buffer A at 0°C. The lines are single exponential fits with koff = 23 × 10−4 sec−1, 9 × 10−4 sec−1, and 78 × 10−4 sec−1, respectively. (B) Comparison of experimental ΔG° values for tRNAThrUGU variants with multiple base-pair substitutions with ΔG° values calculated from the addition of ΔΔG° values determined for single base-pair substitutions. The best linear fit has a slope of 0.9. (C,D) Relationship between the ΔG° values for the E. coli EF-Tu–Thr tRNAThr complex and the T. thermophilus EF-Tu–Phe-tRNAPhe complex (Schrader et al. 2009) and the identity of the 49–65 (C) and 51–63 (D) base pairs. In both cases, data are for single base-pair substitutions of the wild-type tRNA sequence. The best linear fits have slopes of 0.98 and 0.95 when the G51•U63 data point is excluded.
The data in Table 1 show that the affinity between E. coli Thr-tRNAThrUGU and E. coli EF-Tu•GTP is altered when the identity of the 49–65, 50–64, and 51–63 base pairs of the T stem are changed individually and in combination. For the entire set of variants, the free energy of binding (ΔG°) ranges from ∼0.5 kcal/mol tighter to 1.8 kcal/mol weaker than the wild-type tRNAThrUGU transcript (WT). The sequence at each of the three base pairs contributes to the ΔG°, and it appears that the thermodynamic contribution at each site is independent. As shown in Figure 2B, the ΔG° of variants containing two or three base-pair changes can be accurately predicted from the sum of the ΔΔG° values obtained from corresponding single base-pair changes. Thus, for these three base pairs, no neighbor effects are apparent.
It is interesting to compare how the different base-pair substitutions in the T stem affect the binding of E. coli tRNAThr to E. coli EF-Tu with how the same substitutions affect the binding of yeast tRNAPhe to T. thermophilus EF-Tu (Schrader et al. 2009). Figure 2C,D shows that the agreement is extremely good for both the 49–65 and 51–63 base pairs. Since the slopes of both plots are nearing unity, the thermodynamic effect of the different base-pair substitutions at the two positions is quantitatively the same in the two systems. Although there is only limited tRNAThr data for the 50–64 pair, the agreement with the tRNAPhe data is equally good. For example, the ΔΔG° between C50–G64 and G50–U64 is 1.8 kcal/mol in the case of tRNAThr (Table 1, cf. WT and 31) and tRNAPhe. In light of the strong similarity in the sequence specificity of the two proteins, it is interesting that there is a single exception. The G51–U63 pair in tRNAThr stabilizes the interaction with E. coli EF-Tu-tRNAThr by much more than it does in tRNAPhe interacting with T. thermophilus EF-Tu (Fig. 2D).
Viability of T-stem variants in E. coli
tRNA gene knockout and plasmid shuffling approaches were used to test whether T-stem mutations affect viability in E. coli (Saks et al. 1998). These experiments employed a strain of E. coli where the single copy of the essential chromosomal gene for tRNAThrUGU had been inactivated by lengthening the anticodon stem by one base pair and by deleting three residues from the anticodon loop. To propagate the knockout strain, wild-type tRNAThrUGU was expressed from a maintenance plasmid (pMAK705) that confers chloramphenicol resistance and is thermosensitive for DNA replication (Hamilton et al. 1989). The knockout cells were transformed with each of the placZACA plasmids containing the different T-stem variants. Since placZACA is a low-copy plasmid and the tRNAThrUGU variant was transcribed from a copy of the tufB promoter that normally regulates the expression of wild-type tRNAThrUGU from the chromosome, the steady-state level of the variant tRNA in the knockout strain was only slighter greater than its wild-type counterpart in normal E. coli cells (Saks et al. 1998). The two plasmids have compatible replicons and different selectable markers, making it straightforward to confirm that the transformants contained both plasmids (Fig. 3A). These two-plasmid strains were then used to evaluate the function of the tRNAThrUGU variants in two related experiments.
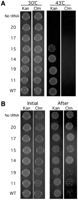
Growth phenotypes and retention of pMAK705 for strains expressing selected tRNAThrUGU variants from the low-copy plasmid (placZACA) that confers kanamycin resistance. Each spot represents equivalent numbers of cells. (A) Growth of E. coli tRNAThrUGU gene knockout strains on plates containing kanamycin (Kan) and chloramphenicol (Clm) at temperatures that are permissive (30°C) or nonpermissive (43°C) for replication of the maintenance plasmid, pMAK705, which confers chloramphenicol resistance. (B) Each strain was continuously grown at 30°C in liquid media containing Kan and the loss of the maintenance plasmid was monitored by testing the initial resistance of each two-plasmid strain to Clm (left panel) and after ∼36 h of growth (right panel).
In the first experiment, each two-plasmid strain was grown at 43°C where pMAK705, the maintenance plasmid producing wild-type tRNAThrUGU, cannot replicate. Thus, cells will only survive if the mutant tRNAThrUGU can function sufficiently well to permit cell growth. Examples of the data from this experiment are given in Figure 3A. The two lanes in the left panel show that each strain contains both placZACA (KanR) and pMAK705 (ClmR) at the outset. In the right panel, the lane marked Kan shows that once pMAK705 is eliminated, strain survival is variant-dependent. Whereas the T-stem sequences of variants 15, 14, 19, 11, and WT are compatible with tRNA function, those of 17 and 20 are not. In all, eight strains did not show any colonies after growth at 43°C. This indicates that their T-stem sequences were incompatible with tRNAThr function. Sixteen strains were able to grow at 43°C and in each case, the cells were sensitive to chloramphenicol, confirming the loss of pMAK705 and indicating that the mutant tRNAThrUGU can support growth of E. coli. Most of the strains showed colony sizes comparable to the wild-type control, but 4 and 14 showed smaller colonies, indicating a slower growth rate. Subsequent experiments showed that the colonies of 4 and 14 are also smaller than those of WT when the strains are grown at 30°C.
A second experiment was used to screen for subtle differences among tRNA variants that were able to support the growth of E. coli. The design of this experiment makes use of the fact that, due to the energetic costs incurred in maintaining extra-chromosomal plasmid DNA, cells tend to lose plasmids over time, unless they confer a counterbalancing benefit (Bouma and Lenski 1988). In this study, plasmid pMAK705 confers two potential benefits when cells are grown at the permissive temperature (30°C): It confers chloramphicol resistance and it also supplies cells with a copy of wild-type tRNAThrUGU. Each two-plasmid strain was grown in liquid media that lacked chloramphenicol but that was supplemented with kanamycin, in order to maintain placZACA, which expresses the T-stem variant. Under these conditions, the strains are expected to lose pMAK705 over time unless the expression of wild-type tRNAThrUGU is advantageous. The strains were grown overnight at 30°C. Fresh cultures were inoculated the following day and were grown to mid-log phase under the same conditions. Equivalent numbers of cells from the strains used for the initial inoculation and from the mid-log cultures were spotted on plates containing either kanamycin or chloramphenicol and incubated overnight at 30°C. Examples of the data from this experiment are given in Figure 3B. The left panel shows that each strain initially is both kanamycin and chloramphenicol resistant, indicating that both plasmids were present. The right panel shows that the strains subsequently differ in their resistance to chloramphenicol, but not kanamycin. The differential retention of chloramphicol resistance, after prolonged growth in chloramphenicol-free liquid media, is indicative that there are strains for which the expression of wild-type tRNAThrUGU from pMAK705 outweighs the energetic costs of maintaining the plasmid. Indeed, the experiments reveal that there are important functional differences among those T-stem variants that are able to support cell growth. One functional category comprises variants such as 19 and WT that are sufficient for E. coli growth without pMAK705 (Fig. 3A) and preferentially lose pMAK705 from the majority of cells such that only a few colonies are formed on media containing chloramphenicol (Fig. 3B). Thus, these variants are fully functional. Seven variants and WT fall into this category (see Fig. 4, green bars). The second functional category comprises variants such as 14 and 15 whose function is sufficient for E. coli growth (Fig. 3A) but that retain pMAK705, if given the option (Fig. 3B). Thus, these variants have a partial functional deficiency that is ameliorated by the co-expression of wild-type tRNAThrUGU. Nine variants fall into this category (see Fig. 4, yellow bars). The third functional category comprises variants such as 20 and 17 which are examples of nonfunctional tRNAThr. They are unable to support E. coli growth in the absence of the WT gene and consequently pMAK705 is retained even when the strains are grown in the absence of chloramphenicol (see the right panels of Fig. 3A,B). Eight of the 24 variants in the study are nonfunctional (see Fig. 4, red bars). In summary, 16 of the 24 variants could not function, or functioned less well than wild type, while only seven variants (and WT) are fully able to replace wild-type tRNAThrUGU.
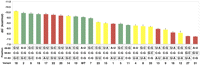
Relationship between cell viability and ΔG° of binding to EF-Tu for tRNAThr variants. Boxes indicate the base pairs that were changed to create each variant. Bar height indicates the ΔG° of binding (Table 1), and bar color indicates the effectiveness of each variant in replacing the inactivated tRNAThrUGU chromosomal gene. Green are fully functional variants where cells spontaneously lose pMAK705. Yellow are moderately functional variants where cells do not need pMAK705 to grow, but do not lose it spontaneously. Red are nonfunctional variants that do not grow without pMAK705 (see text and Fig. 3).
As shown in Figure 4, the relationship between a variant's ability to support growth in E. coli and its affinity for EF-Tu in vitro is not clear-cut. Several variants which bind EF-Tu as well as or tighter than wild type are either unable to support growth (17, 20, 22) or do so less well (14, 18). Most variants that bind less well than WT either do not grow or show impaired function. However, there are two exceptions. Variants 5 and 11 effectively replace tRNAThr even though their binding to EF-Tu is 0.5 kcal/mol weaker than WT. It is unclear why the T-stem sequences of variants 5 and 11 result in effective growth. Only the C50–G64 pair is common between the two variants and this pair is present in nonfunctional variants as well. The lack of a clear relationship between T-stem sequence and cell viability is frequently observed. For example, while the single base-pair variant A51–U63 (1) is inviable, the pair is present in two functional (5, 9) and two nonfunctional (13, 17) variants that have multiple base-pair substitutions. Thus, when base pairs are considered individually, their different identities do not account for the functional differences of the corresponding tRNA variants. Rather, the consequence of a single base-pair substitution can be contingent on the identity of residues at additional sites in the tRNA.
It was possible that some of the variants fail to support growth in E. coli because they either are not properly processed or are rapidly degraded. This was tested by extracting RNA from two-plasmid strains where tRNAThr function was either partially or entirely impaired. Northern blots detected only full-length, correctly processed tRNA in every case. Similarly, dot blot experiments suggested that the steady-state levels of all the variants were roughly similar to wild-type tRNAThr (data not shown). Thus, the lack of growth of the eight inviable strains is unlikely to be due to altered tRNA processing or a change in the steady-state concentration of the mutant tRNA.
Identification and characterization of intragenic revertants
The discovery of mutations of tRNAThr that did not support growth permitted a search for intragenic revertants. The gene for variant 20, which is nonfunctional due to nucleotide substitutions that alter the 49–65 and 51–63 pairs (Fig. 5A), served as the starting point for these experiments. The gene was subjected to error-prone PCR and the resulting second-generation variants were cloned and subsequently transformed into the knockout strain. Those variants that conferred viability, after elimination of the maintenance plasmid, were sequenced, recloned, and their ability to confer viability was confirmed. Three different point mutations converted variant 20 from nonfunctional to functional (Fig. 5A). Two of these, G49A and U65C, were among the group of 16 viable variants characterized previously, thereby confirming the effectiveness of the selection procedure. The third compensatory mutation, A52G, converts the A52–C62 pair present of wild-type tRNAThrUGU into a G–C pair.
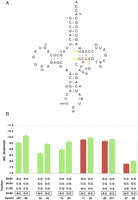
Mutations that restore the function of variant 20. (A) The sequence of variant 20, which fails to support E. coli growth, with the deleterious mutations indicated in red. Arrows indicate three separate point mutations that restore its function. (B) The effect of the A52G mutation on the ΔG° of binding of variants to EF-Tu and on their ability to support growth in the absence of pMAK705. Bar color distinguishes variants that support E. coli growth in the absence of pMAK705 (green) from those that do not (red).
To better understand the role of the A52G mutation in tRNAThr function, two functional variants (33, 15) and four additional nonfunctional variants (1, 16, 17, 27) were inserted into wild-type tRNAThr. When tested for their ability to support growth in the absence of pMAK705, all seven of the A52G-containing variants conferred viability. Thus, not only was the A52G mutation of wild-type tRNAThr able to support growth, but A52G successfully reverted the phenotype of all of the inviable mutations tested.
The effect of introducing the A52G mutation on EF-Tu binding was tested with wild-type tRNAThr, two functional, and three nonfunctional variants (Table 2). The binding data are presented together with the viability data in Figure 5B. For wild-type tRNAThr and the two functional variants, the A52G mutation stabilizes the ΔG° of binding by ∼0.5 kcal/mol. However, in the case of all three nonfunctional variants, the A52G mutation has little or no stabilizing effect on EF-Tu binding. These experiments indicate that the A52–C62 pair present in wild-type tRNAThr reduces the affinity to EF-Tu. In addition, in the case of the A52G mutation, there appear to be sequence-context effects not previously observed in EF-Tu binding studies.
Dissociation rates of tRNAThrUGU variants from E. coli EF-Tu
DISCUSSION
The free energies of binding of 34 different variants of E. coli Thr-tRNAThrUGU to E. coli EF-Tu•GTP determined here provide an overview of the sequence specificity of this tRNA–protein interaction. When these data are compared to the similarly large data set measuring the binding of variants of yeast tRNAPhe with T. thermophilus EF-Tu•GTP, it is clear that the two systems show nearly identical sequence specificity despite the fact that both the protein and tRNA used in the experiments were different. Not only were the ΔΔG° values resulting from different single base-pair substitutions quantitatively identical in the two systems, but the effects at the three T-stem sites were thermodynamically additive, making it possible to accurately predict the effect of multiple base-pair substitutions in both cases. A similar conclusion was recently reached with a more limited set of E. coli tRNAVal mutants binding to E. coli EF-Tu (Schrader et al. 2011b). Although the two proteins are only 71% identical, cocrystal structures show the same set of amino acid side chains interacting with aa-tRNA (Nissen et al. 1995, 1999; pdb 10B2). Indeed, almost all of the amino acids at the interface that make thermodynamically important contacts are conserved in all bacteria (Sanderson and Uhlenbeck 2007b). This suggests that all bacterial EF-Tu proteins interact with aa-tRNA in a very similar manner.
Despite the similarity of the two data sets, there were two features of the Thr-tRNAThr–E. coli EF-Tu interaction that had not been encountered in earlier studies of tRNA binding to T. thermophilus EF-Tu. First, despite the fact that all other mutations of the 51–63 base pair behaved identically in the two systems, the G51–U63 pair stabilizes the binding of tRNAThr to E. coli EF-Tu by 0.4 kcal/mol more than it stabilizes the binding of tRNAPhe or tRNALeu to T. thermophilus EF-Tu (Sanderson and Uhlenbeck 2007c; Schrader et al. 2009). It is known that in both E. coli and T. thermophilus EF-Tu, a stabilizing hydrogen bond forms between a glutamate and the amino group of G at either the 51 or 63 position of several tRNAs (Roy et al. 2007; Sanderson and Uhlenbeck 2007a; Schrader et al. 2011b). Thus, the amount that this hydrogen bond contributes to the affinity not only depends upon the identity of the 51–63 pair, but also the tRNA context. The second new feature of the tRNAThr–E. coli EF-Tu interaction is the 52–62 base pair's contribution to the sequence specificity. Here, binding to EF-Tu is tightened when the wild-type A52–C62 pair of tRNAThrUGU is changed to G52–C62, but these two pairs bind equivalently to T. thermophilus EF-Tu when they were tested in tRNAPhe (Schrader et al. 2009). Both these context-dependent features of the interaction between tRNAThr and EF-Tu are likely to be due to the A52–C62 pair. Since the phosphodiester backbone of the 52–62 pair contacts the protein and contributes to ΔG° (Pleiss and Uhlenbeck 2001; Sanderson and Uhlenbeck 2007a), the mismatched A–C pair may result in a more dynamic or altered backbone conformation that not only weakens binding but somehow modifies how the adjacent G51–U63 mutant is recognized. When the A–C pair is changed to G–C, the reduced dynamics could tighten binding and alter the positioning of the adjacent 51–63 pair. This model could also reconcile the differing effects on binding when the A52G mutant is introduced into different T-stem variants as shown in Figure 5.
The A52–C62 pair in E. coli tRNAThr can be considered to be a “negative” specificity element that evolved to weaken binding to EF-Tu in order to offset the positive elements in the T stem that stabilize the complex. The C1–A72 pair in E. coli tRNAfMet also acts as a negative element to help to ensure that the initiator tRNA does not bind EF-Tu (Seong and RajBhandary 1987). There is evidence that the A1–U72 and U1–A72 pairs in several E. coli elongator tRNAs also act as negative elements, possibly due to their propensity to fray (L Behlen and O Uhlenbeck, unpubl.). Thus, while bacterial EF-Tu appears to use a common set of amino acids to recognize aa-tRNA, the thermodynamic details of how particular aa-tRNAs contact the protein can be somewhat different. While all tRNAs appear to use the sequence of three T-stem base pairs as important specificity determinants, some, such as tRNAThr, use additional sequence elements.
It is interesting that the A52–C62 pair is extremely rare, occurring in <1% of bacterial tRNAs and primarily appearing in only three tRNA species: tRNAThrUGU, tRNAProGGG, and tRNAProUGG. The A52–C62 pair in tRNAThrUGU primarily occurs in γ-proteobacteria, whereas other bacteria most often use G52–C62. Thus, by choosing to work on E. coli tRNAThr, we have uncovered a relatively rare EF-Tu recognition element.
The function of tRNA T-stem variants was assayed in vivo using a tRNA gene replacement strategy in combination with a tRNAThrUGU knockout strain. These experiments determined whether a variant tRNA can support growth on its own and, if it can, whether growth is aided by the presence of wild-type tRNA. With the exception of recent experiments using a highly engineered heterologous suppressor tRNA (Guo et al. 2009), this study is the first systematic analysis of how T-stem mutations affect tRNA function in vivo. Since all of the mutations were base-pair changes expected to minimize improper tRNA folding, it was striking that a large proportion of the mutations led to a detectable growth defect. Eight of the 24 mutations could not replace the wild-type tRNAThrUGU at all, while nine showed sufficiently reduced function that E. coli preferred the fitness cost of retaining an additional plasmid with a wild-type copy of the gene. This result supports the notion that the T-stem sequence of tRNAThr (and presumably all other E. coli tRNAs) is carefully “tuned” by evolution for optimal function. Since seven T-stem mutations were able to replace wild-type tRNAThrUGU, it appears that multiple T-stem sequences are functionally equivalent. However, it remains possible that some or all of these viable variants function slightly less efficiently than wild-type since competitive growth experiments over many generations were not performed.
It is interesting that the T stem of tRNAThrUGU from Chromobacterium violaceum (Brazilian National Genome Project Consortium 2003) has a sequence identical to that of our nonfunctional variant 22. This emphasizes that the evolution of T-stem sequences is highly idiosyncratic for a given tRNA and that tRNA function also may require additional sequence elements outside of the T stem. An important goal is to understand this complex tuning process that governs the sequence of the T stem.
Biochemical experiments have found that a potentially important role of the T-stem sequence in translation is to tune the affinity of each tRNA to EF-Tu to a value that offsets the variable contribution of the esterified amino acid. Two different steps in translation collaborate in the evolution of these tuning elements (Schrader et al. 2011b). If the ΔG° for a given aa-tRNA is too weak, the concentration of the corresponding ternary complex would be too low for efficient delivery to the ribosome. If the ΔG° for an aa-tRNA is too tight, recent experiments have shown that the rate of release of aa-tRNA from EF-Tu•GDP limits the rate of peptide bond formation (Schrader et al. 2011b). Based on these experiments, one might expect that either increasing or decreasing the affinity of tRNAThrUGU to EF-Tu could lead to a defect in growth. Although we have shown that many of the base-pair changes in the T stem that lead to tighter or weaker binding do indeed cause growth defects, Figures 4 and 5B show that a clear correlation between EF-Tu affinity and growth does not exist. Several variants that bind either weaker or tighter than wild-type remain viable. One straightforward explanation for this disparity is that since the buffer and temperature used for the in vitro binding experiments are so different from conditions inside cells, the correlation between viability and affinity is lost. It is known that the affinity of EF-Tu to aa-tRNA at 37°C is much weaker than at 0°C where the affinities are usually measured (Vorstenbosch et al. 2000) and this temperature difference could affect the sequence specificity as well. For example, the higher temperature could magnify the destabilizing effect of the mismatched A52–C62 pair in tRNAThr on EF-Tu binding, thereby altering the relative effects of mutations at other T-stem positions. Unfortunately, the lability of the aminoacyl bond and the very rapid koff values at higher temperatures make it difficult to assay the mutants at temperatures where cells grow. In addition, the unknown intracellular ionic environment and the potential contributions of molecular crowding complicate a quantitative comparison of the two sets of data.
An alternate explanation for the lack of correlation between the growth and binding data is that growth also may be affected by a different step in the translation pathway. Although the T stem of tRNA does not make many contacts with the ribosome in the A, P, and E sites, there are a few that could potentially lead to sequence-specific effects (Selmer et al. 2006; Jenner et al. 2010). In addition, the T-stem sequence may influence intermediates in translation for which no structural data are available. The availability of a large set of T-stem mutations that affect growth to differing degrees should be valuable reagents for future, more detailed biochemical studies examining their properties in translation.
MATERIALS AND METHODS
EF-Tu binding experiments
The gene for each tRNAThrUGU variant was cloned into placZACA (W Liu and M Saks, unpubl.) and the intended sequence changes were verified by sequencing using BigDye terminator (ACGT, Inc.). DNA templates for in vitro run-off transcription with the P266L mutant of T7 RNA polymerase (Guillerez et al. 2005) were obtained from the cloned genes by PCR amplification using Vent polymerase (New England BioLabs) followed by restriction endonuclease digestion with BstN 1 or with Bsa 1 (New England BioLabs) in the cases where the tRNA variant had an internal BstN 1 site. The RNA products were purified on 8% denaturing polyacrylamide gels. The tRNA variants were aminoacylated with [3H]Thr (20 ci/mmol, American Radiolabeled Chemicals), ethanol-precipitated, and stored at −20°C in 5 mM NaOAc pH 5.2.
E. coli EF-Tu was purified using an expression clone that contained an N-terminal His6 tag and a TEV protease site. After expression in BL21-Gold (DE3) cells (Agilent), the protein was purified by Ni-NTA chromatography cleaved with TEV protease and further purified as described in Sanderson and Uhlenbeck (2007b). EF-Tu, stored in the GDP-bound form, was activated to its GTP-bound form by incubating at 37°C for 30 min in buffer A (50 mM HEPES-KOH pH 7.0, 20 mM MgCl2, 0.5 M NH4Cl, 5 mM DTT) with 10 μM GTP, 3 mM phosphoenolpyruvate, and 12.5 U/mL pyruvate kinase. Ternary complexes were formed by incubation of 1 μM activated EF-Tu•GTP and <0.2 μM [3H]Thr-tRNAThr at 0°C. The dissociation rate constant (koff) of each [3H]Thr-tRNAThr variant from EF-Tu•GTP was determined in buffer A using a ribonuclease protection assay at 0°C and analyzed as described (Schrader et al. 2009).
Cell growth experiments
Details about the construction of the tRNAThrUGU gene knockout strain, including the use of homologous recombination and P1 transduction for chromosomal manipulation and the strategy used to inactivate the chromosomal copy of the wild-type gene for tRNAThrUGU, are given in Saks et al. (1998). Details concerning the construction of placZACA, the low-copy plasmid that is used to express the tRNA variants, in vivo, and that confers kanamycin (Kan) resistance, are given in W Liu and M Saks (unpubl.). PCR mutagenesis was performed as described (Svetlov and Cooper 1998).
The knockout strain, which expresses tRNAThrUGU from pMAK705 (ClmR) was transformed with placZACA (KanR) expressing the different tRNA variants. The presence of both plasmids was verified by growing cells at 30°C (the permissive temperature) on two types of selective plates (LB/Clm [25 μg/mL] and LB/Kan [20 μg/mL]). Prior to each experiment, strains were grown in LB supplemented with kanamycin overnight at 30°C, to bring them to similar physiological states and to prevent the loss of placZACA bearing the test tRNAs. Fresh cultures were inoculated at 0.05 A600/mL, grown to approximately mid-log phase at 30°C in LB/Kan, and diluted so that similar numbers of cells were used in the growth assays, regardless of genotype.
ACKNOWLEDGMENTS
We thank Jared Schrader for useful discussions and Dr. Emine Yikilmaz and Stephen Chapman for purified E. coli EF-Tu. This work was funded by National Institutes of Health Grant GM037552 (to O.C.U.) and by National Institutes of Health Grant GM059779 (to M.E.S.).
Footnotes
-
Reprint requests to: Margaret E. Saks, Department of Molecular Biosciences, Northwestern University, 2205 Tech Drive, Hogan 2-100, Evanston, IL 60208, USA; e-mail: m-saks{at}northwestern.edu; fax: (847) 491-5444; or Olke C. Uhlenbeck, Department of Molecular Biosciences, Northwestern University, 2205 Tech Drive, Hogan 2-100, Evanston, IL 60208, USA; e-mail: o-uhlenbeck{at}northwestern.edu; fax: (847) 491-5444.
-
Article published online ahead of print. Article and publication date are at http://www.rnajournal.org/cgi/doi/10.1261/rna.2427311.
- Received October 12, 2010.
- Accepted March 22, 2011.
- Copyright © 2011 RNA Society








