
A fast selenium derivatization strategy for crystallization and phasing of RNA structures
- Vincent Olieric1,5,
- Ulrike Rieder2,5,
- Kathrin Lang2,
- Alexander Serganov3,
- Clemens Schulze-Briese1,
- Ronald Micura2,
- Philippe Dumas4,6 and
- Eric Ennifar4,6
- 1Swiss Light Source, Paul Scherrer Institute, CH-5232 Villigen, Switzerland
- 2Institute of Organic Chemistry, Center for Molecular Biosciences, Leopold Franzens University, A-6020 Innsbruck, Austria
- 3Structural Biology Program, Memorial Sloan-Kettering Cancer Center, New York, New York 10021, USA
- 4Architecture et Réactivité de l'ARN, Institut de Biologie Moléculaire et Cellulaire, Université Louis Pasteur/CNRS UPR 9002, F-67084 Strasbourg, France
-
↵5 These authors contributed equally to this work.
Abstract
Site-specific 2′-methylseleno RNA labeling is a promising tool for tackling the phase problem in RNA crystallography. We have developed an efficient strategy for crystallization and structure determination of RNA and RNA/protein complexes based on preliminary crystallization screening of 2′-OCH3-modified RNA sequences, prior to the replacement of 2′-OCH3 groups with their 2′-SeCH3 counterparts. The method exploits the similar crystallization properties of 2′-OCH3- and 2′-SeCH3-modified RNAs and has been successfully validated for two test cases. In addition, our data show that 2′-SeCH3-modified RNA have an increased resistance to X-ray radiolysis in comparison with commonly used 5-halogen-modified RNA, which permits collection of experimental electron density maps of remarkable quality.
Keywords
INTRODUCTION
The growing number of biologically important RNA sequences demands a fast and reliable method for crystallization and de novo three-dimensional structure determination of RNA. 2′-methylseleno (2′-SeCH3) labeling of nucleic acids has been developed as an alternative to two broadly used phasing techniques, to the halogenation of pyrimidines and to soaking crystals in heavy atom salt solutions (Du et al. 2002; Teplova et al. 2002; Carrasco et al. 2004; Jiang et al. 2007; Pallan and Egli 2007). The 2′-SeCH3-labeling method, suited for the determination of RNA structures, relies on the crystallization of RNA molecules that have natural nucleoside(s) substituted for their 2′-SeCH3-modified counterparts during chemical RNA synthesis. The 2′-SeCH3 labeling allows use of the powerful multiwavelength anomalous dispersion (MAD) technique for structure determination and, in contrast to the halogenation of pyrimidines, can be utilized for the modification of all four standard nucleosides (adenosine, guanosine, uridine, and cytidine) (Hobartner and Micura 2004; Hobartner et al. 2005; Moroder et al. 2006; Puffer et al. 2008). Moreover, the combination of the 2′-SeCH3 labeling with enzymatic ligation expands the ∼40 nucleotide (nt) limit of chemically synthesized Se-labeled RNA to large biologically relevant RNA molecules (Hobartner et al. 2005). The Se-labeling strategy was successfully applied to determine several nucleic acid structures, including the structure of a 49-nt Diels–Alder ribozyme determined by single-wavelength anomalous dispersion (SAD) (Hobartner et al. 2005; Serganov et al. 2005).
Due to the high costs of preparative Se-labeled RNA synthesis, initial crystallization trials have to be performed using unmodified RNA. However, the substitution of the 2′-OH group for the 2′-SeCH3 group can alter crystal contacts, thereby resulting in the deterioration of crystal quality and in interference with crystallization conditions established for unmodified RNA (Hobartner et al. 2005; Moroder et al. 2006). As a consequence, crystallization conditions either cannot be reproduced or require the synthesis of several RNA sequences with 2′-SeCH3 modifications at various positions (Hobartner et al. 2005; Moroder et al. 2006). To obtain a reproducible transition from the nonmodified RNA to the Se-labeled RNA, we propose a new crystallization strategy based on a broad preliminary crystallization screening of the RNA sequences with 2′-OH group(s) substituted for 2′-OCH3 group(s) (Fig. 1), followed by focused crystallization and structure determination using crystallizable RNA variants with 2′-OCH3 group(s) substituted for 2′-SeCH3 group(s) (Fig. 2). The 2′-OCH3 group is chemically analogous to the 2′-SeCH3 group (Fig. 1). However, the incorporation of the 2′-OCH3-modified nucleoside into RNA is less expensive and easier. As we demonstrate in the current study, RNAs derivatized by either 2′-SeCH3 or 2′-OCH3 groups behave similarly in crystallization trials, thus providing an easy, inexpensive, and fast approach to the crystallization and three-dimensional structure determination of various RNA or RNA/protein complexes.
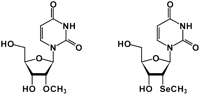
Chemical structure of 2′-methylseleno-uridine and 2′-O-methyl-uridine.
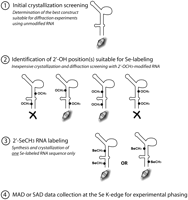
Schematic representation of the proposed strategy for selenium derivatization and phasing of RNA crystal structures.
RESULTS AND DISCUSSION
Structure determination of the HIV-1 DIS/aminoglycoside complex using selenium-labeled RNA
The approach has been developed during the structure solution of the 46-nt HIV-1 genomic RNA dimerization initiation site (DIS) bound to aminoglycoside antibiotics. This RNA/drug complex involves a 23-nt RNA homodimer, which promotes viral genome dimerization and is recognized by aminoglycosides (Ennifar et al. 2003, 2007b; Bernacchi et al. 2007; Freisz et al. 2008). In parallel to an extensive but unsuccessful search for heavy atom derivatives, we attempted to determine the structure using 5-halogen-pyrimidine derivatives. Despite multiple trials using 5-bromo- and 5-iodo-RNAs, crystals were obtained only for a 5-iodo derivative. Data sets collected for these crystals at both low-energy (to maximize the anomalous signal) and high-energy (to reduce crystal absorption and radiation damage) wavelengths showed good quality crystallographic statistics (Supplemental Table 1). However, our attempts to determine the structure using multiple isomorphous replacement (MIR), MAD or SAD techniques have failed, most likely because of strong site-specific radiation damage previously reported on 5-halogen pyrimidines (Ennifar et al. 2002). A correction for the radiolysis by a dose-dependent occupancy refinement of iodine atoms, recently implemented in the program SHARP (Schiltz et al. 2004), was not successful. The 2′-SeCH3 RNA labeling strategy was therefore investigated to determine the structure of the RNA/drug complex.
Three sequences were synthesized with selenium atoms introduced at various positions in the DIS stem: U266, U267, and U270 (Fig. 3). Out of these three sequences, only the U267-SeCH3 DIS variant produced diffraction quality crystals. Interestingly, as reported previously for a 2′-SeCH3-labeled DNA duplex (Jiang et al. 2007), these Se-derivatized crystals grew significantly faster and larger than crystals with unmodified RNA, leading to an improvement in the diffraction limit (Table 1). Poorly diffracting crystals and spherulites were obtained with the U266-SeCH3 DIS RNA, and no crystals were found for the U270-SeCH3 DIS sequence (Fig. 3). A single U267-SeCH3 crystal was used to collect highly redundant data at the Se-K edge in a three-wavelength MAD experiment (Table 1). This data set produced an experimental electron density map of exceptional quality (Fig. 4a), thus emphasizing the high quality of experimental phases obtained from Se-derivatized RNA.
Data collection and refinement statistics for unmodified, U267-OCH3 and U267-SeCH3 DIS RNAs bound to paromomycin or ribostamycin antibiotics (Freisz et al. 2008)

Sequences and crystallization results obtained for the HIV-1 genomic RNA dimerization initiation site extended duplex bound to ribostamycin aminoglycoside (Freisz et al. 2008) according to the scheme proposed on Fig. 2. On the unmodified sequence (①), green and red arrows indicate 2′-OH positions available for 2′-OCH3/SeCH3 substitution or involved in crystal packing, respectively. The distance to the closest atom is indicated in green for available positions, and the number of hydrogen bonds involved in the packing interaction is shown in red.
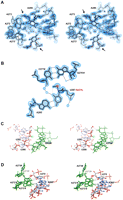
HIV-1 DIS RNA/drug complex (Freisz et al. 2008). (A) Stereo view of the experimental MAD electron density map. The map clearly shows the bulge region (A272, A273, A280) of RNA, two ribostamycin aminoglycosides (indicated with black arrows), and surrounding water molecules (red spheres). (B) View of the U267-SeCH3 A285′ base-pair in the HIV DIS structure at 1.5 Å resolution. Se atom is depicted by a pink sphere. 2Fo-Fc electron density map is represented around the model. The C277-G276′ base-pair from a symmetry-related molecule is also shown. Hydrogen bonds and the intraresidue C-H(methyl)-O bond in the modified uridine are represented with black and gray dotted lines, respectively. (C, D) Stereo views showing crystal packing interactions around U266, U267 (C), and U270 (D) residues. Symmetry-related molecules are shown in green, and nucleotides are indicated with a hash mark (#). 2′-OH groups of U266 and U270, which do not allow 2′-SeCH3 modifications, are shown with a pink circle. The 2′-OH group of U267, which can accommodate 2′-SeCH3 modification, is depicted with a green circle. The hydroxyl of U266 interacts with a 2′-OH group from a symmetry-related molecule (C), and the hydroxyl of U270 is involved in an A minor interaction with a bulged-out A272′# (D), whereas 2′-OH of U267 is not H-bonded and available for 2′-SeCH3 modification (C). Residues U266′, U267′, and U270′ from the second molecule in the asymmetric unit are not involved into crystal packing (not shown).
Subsequent examination of the 2′-SeCH3-modified residues in the structure revealed an intraresidue C-H(methyl)-O4(U) hydrogen bond that restricts rotation of the methyl group (Fig. 4b). Analysis of the DIS/ribostamycin crystal structure showed that the 2′-OH groups of U266 and U270 in one DIS strand are involved in crystal packing, whereas the 2′-hydroxyl of U267 in both strands does not provide crystal contacts (Fig. 4c,d). We therefore reasoned that the 2′-OCH3 modification can be regarded as isosteric to the 2′-SeCH3 one and, thus, could be used to screen 2′-OH positions available for the 2′-SeCH3 modification. To test this hypothesis, a 2′-OCH3 modification was introduced into U266, U267, and U270 of the DIS RNA (Fig. 3), and the crystallization and diffraction properties of these variants were compared with the crystallization behavior of the corresponding 2′-SeCH3-modified sequences. As anticipated, the 2′-OCH3-modified DIS sequences demonstrated a good correlation with the 2′-SeCH3-modified RNAs: no crystal was obtained for the U270-OCH3 DIS RNA, spherulites and poorly diffracting crystals were found for the U266-OCH3 DIS variant, and well-shaped diffracting crystals were obtained for the U267-OCH3 DIS RNA (Fig. 3; Table 1).
Validation of the strategy: structure determination of the Escherichia coli sarcin/ricin loop
We then validated our approach using the 27-nt fragment of E. coli 23 S rRNA sarcin/ricin loop (SRL) region (Correll et al. 1999). Analysis of the SRL structure (Protein Data Bank [PDB] identification no. 483D) revealed that the 2′-OH groups of U2650 and U2656 are not involved in the crystal contacts and, hence, are available for modifications, whereas the 2′-OH of U2653 is hydrogen bonded with a symmetry-related molecule (Fig. 5, ①). Four SRL sequences were therefore synthesized: an unmodified SRL (used as a control for crystallization and diffraction) and three modified sequences with the introduction of a 2′-OCH3 group into U2650, U2653, or U2656 (Fig. 5). Crystallization trials revealed that, as expected, only U2650 and U2656 accommodated the 2′-OCH3 modification and produced crystals, whereas modification of U2653 precluded crystallization (Fig. 5). Following this preliminary 2′-OCH3 screening of the SRL sequence, 2′-SeCH3-uridines were introduced at the 2650 and 2656 positions. In agreement with our previous results, both sequences produced crystals in every respect identical to unmodified or 2′-OCH3-containing SRL sequences, thus confirming the excellent correlation between 2′-OCH3 and 2′-SeCH3 modifications. The Se-derivatized SRL crystals were then used for a classical three-wavelength diffraction experiment (Table 2). Both U2650-SeCH3 and U2656-SeCH3 SRL structures were easily determined by SAD and MAD, which produced similar experimental electron density maps of exceptional quality (Figs. 6, 7), additionally illustrating the efficiency of selenium derivatization. Interestingly, diffraction data collected at atomic resolution for U2650- and U2656-OCH3 and -SeCH3 SRL confirmed the presence of the C-H(methyl)-O4(U) intraresidue bond that stabilizes the methyl group (Fig. 7).
Data collection and refinement statistics for SRL RNA (Correll et al. 1999) sequences used in this study
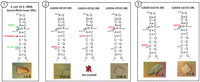
Sequences used and crystallization results obtained for the E. coli 23 S rRNA sarcin/ricin loop (Correll et al. 1999) according to the scheme proposed on Fig. 2.

Experimental MAD electron density map (contoured at 1.5 σ level) obtained with the U2656-SeCH3 SRL RNA. (A) Stereo view showing residues 2651–2669. (B) Detailed view of the A2657-G2664 mismatch. Electron density maps of similar quality were obtained by SAD phasing with U2656-SeCH3 RNA, and by MAD/SAD phasing with the U2650-SeCH3 SRL sequence.
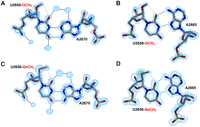
Detailed views of the labeled U2650·A2670 (A, C) and of the labeled U2656·A2665 reverse wobble (B, D) base-pairs in SRL structures. (A, B) The 2Fo-Fc electron density map at 0.97 Å and 0.96 Å resolution are represented around the refined model. (C) The experimental SAD electron density map at 1.1 Å resolution is shown around the U2650-SeCH3 A2670. (D) The experimental MAD electron density map at 1.0 Å resolution, as well as the anomalous map contoured at 40 σ level (in green), is shown for the U2656-SeCH3 SRL structure. Spheres of electron density around RNA correspond to water molecules. Hydrogen bonds and the intra-residue C-H(methyl)-O bond in modified uridines are represented with black and gray dotted lines, respectively.
Site-specific radiation damage studies of 2′-SeCH3 and comparison with 5-halogen pyrimidines
To demonstrate the greater resistance to radiation damage of 2′-SeCH3-labeled over 5-halogen-labeled RNA, a detailed comparison of site-specific radiolysis was performed using DIS or SRL RNA crystals containing 2′-SeCH3 modifications, and 5-bromo- or 5-iodo-uridines. Using heavy atom occupancies derived from the anomalous signal that was calculated from small data subsets (Olieric et al. 2007), we found that radiolysis had little effect on the C-Se bond. In contrast, 5-iodo- and 5-bromo-uridines are significantly affected by site-specific radiation damage during data collection (Fig. 8). The resistance toward radiolysis accounts for the excellent quality of SAD/MAD experimental maps obtained from 2′-SeCH3 RNA derivatives, thus reinforcing its interest for nucleic acids phasing.
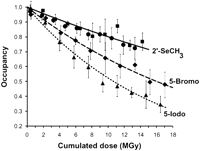
Site-specific radiation damage studies of modified RNA. Comparison of occupancies derived from anomalous difference maps for data collected on U267-Iodo DIS (triangle, dotted line), U267-Bromo DIS (diamonds, dashed line), U267-SeCH3 DIS (squares), and U2656-SeCH3 SRL (spheres) RNA crystals. The cumulative dose limit of 15 - 18 MGy was chosen as it corresponds to the upper limit of doses typically absorbed by a crystal during MAD or SAD data collection.
Conclusion
Our results demonstrate excellent correlation in the crystallization of 2′-OCH3– and 2′-SeCH3-modified RNAs and suggest the use of 2′-OCH3-modified RNA sequences for a preliminary search of 2′-OH groups available for selenium labeling (Fig. 2). The introduction of 2′-OCH3 groups can be routinely achieved by chemical synthesis. The low costs of 2′-OCH3 ribonucleotide building blocks make this approach less expensive than the synthesis of multiple variants of 2′-SeCH3-modified RNA. The greatest advantage, however, is the efficiency of screening with 2′-OCH3 nucleosides: the extensive time required for multistep syntheses of commercially unavailable Se-modified adenosine, cytosine, and guanosine nucleoside phosphoramidites is compensated by high hit-rate structure determination when Se-labels are only positioned at the most promising sites that have been identified via 2′-OCH3 nucleosides. This strategy can obviously be extended to structure determination of RNA (or DNA)/protein complexes when the classical approach with selenomethionine-modified proteins failed. Based upon what is commonly considered for the structure determination of proteins (about one selenomethionine per 20 kDa), one SeCH3-modified position (preferably in a canonical helical region) per ∼50 nt is probably a safe rule. We also anticipate that 2′-OCH3-modified RNA can be directly used for the initial screening of crystallization conditions, thereby eliminating the search for crystallization conditions using unmodified RNA. Finally, our data show that, unlike 5-halogen pyrimidines, 2′-SeCH3-modified nucleosides are strongly resistant to site-specific radiation damage and are therefore more suitable for SAD or MAD experiments, especially using small crystals and third generation synchrotrons.
MATERIAL AND METHODS
Chemical synthesis of RNA
Unmodified 23-nt DIS, 5-bromo- and 5-iodo-uridine DIS, and 2′-OCH3-modified DIS sequences were purchased from Dharmacon. All RNAs containing 2′-methylseleno uridines were synthesized according to previously published procedures (for details, see Hobartner and Micura 2004; Micura et al. 2007). To guarantee a broad dissemination of the approach presented here, we exemplify the 2′-OCH3/2′-SeCH3 strategy for uridine since the corresponding 2′-SeCH3-uridine phosphoramidite is commercially available (http://www.glenresearch.com). The syntheses of 2′-SeCH3 cytidine, 2′-SeCH3 adenine, and 2′-SeCH3 guanosine phosphoramidites have been described (Hobartner et al. 2005; Moroder et al. 2006; Puffer et al. 2008), however, these building blocks have not yet been commercialized. The approach presented here works in similar manner for adenine, cytidine, and guanosine (data not shown).
Crystallization of RNA samples, data processing, and structure refinement
The HIV-1 DIS duplexes bound to ribostamycin or paromomycin aminoglycosides were crystallized as described previously (Freisz et al. 2008). Crystals were grown for 7–10 d at 37°C with unmodified and with 5-iodo-U266 sequences, and for 1–2 d at 37°C with U267-SeCH3 DIS RNA. No DTT was added to crystallization conditions to prevent possible oxidization of the 2′-SeCH3 groups to 2′-methylselenoxide (Moroder et al. 2006). No crystals were obtained with 5-iodo-U267, 5-bromo-U266, or 5-bromo-U267 modified RNA. Crystals were frozen in liquid ethane prior data collection. Data were processed with the HKL Package (Otwinowski and Minor 1996) and D*TREK (Pflugrath 1999). The asymmetric unit (A.U.) contains two DIS molecules and two aminoglycosides. Two heavy atoms in the A.U. were located with SHELX (Sheldrick 2008), and the structure was solved using SHARP (de la Fortelle and Bricogne 1997). For radiation damage studies, 5-iodo- and 5-bromo-U267 DIS loop–loop complex bound to lividomycin aminoglycoside were crystallized as described earlier (Ennifar et al. 2006) and frozen in liquid ethane.
The 27-nt SRL hairpin was crystallized according to the method described by Correll et al. (1999). This sequence was chosen as a test case because crystallization conditions easily produce well-diffracting crystals. Crystals were grown for 1–3 d at 20°C for unmodified, 2′-OCH3-modified, and 2′-SeCH3-modified sequences. About 80% of drops produced crystals (18 drops were prepared for each SRL sequence). Glycerol was used instead of xylitol for cryoprotection, and crystals were frozen in liquid ethane for data collection. Data were processed with the XDS Package (Kabsch 1993) and D*TREK. The A.U. contains one SRL molecule. Heavy atom searches and structure determination were performed with SHELX.
Structures were refined with CNS (Brunger et al. 1998) and PHENIX (Adams et al. 2002). Because of high B factors, the methyl group was not visible on both strands in the U267-OCH3 DIS structure at 2.0 Å resolution and on one strand on the U267-SeCH3 DIS structure at 1.5 Å resolution. Coordinates have been deposited to the PDB.
Site-specific radiation damage analysis
All radiation damage experiments were carried out at the X06SA macromolecular crystallography beamline of the Swiss Light Source (SLS) at Paul Scherrer Institut, Villigen, Switzerland. Data were collected at 90 K on a Pilatus 6M detector (Dectris AG) on five U2656-SeCH3 SRL crystals, two U267-SeCH3 DIS duplex/Ribostamycin crystals (Freisz et al. 2008), four U267-Bromo DIS kissing-complex/Lividomycin crystals (Ennifar et al. 2006), and three U267-Iodo DIS kissing-complex/Lividomycin crystals (Ennifar et al. 2007a). Data for seleno and iodo derivatives were collected at 12.6647 keV and at 13.4895 keV for bromo derivatives. For all crystals, 3600° (7200 images) were collected in a row with a rotation increment of 0.5° per image and an exposure time of 0.2 sec per image. Such data collection took only 24 min using the Pilatus 6M detector. The X-ray beam was defocused in the vertical direction (150 × 80 μm) to obtain homogenous X-ray irradiation of the crystal during data collection (Schulze-Briese et al. 2005). Beam transmission was adapted to reach a dose of ∼15 MGy at the end of each data collection. The incident photon flux was monitored by using a PIN diode, and the beam position was monitored using a CVD diamond quadrant beam-position monitor. The doses were calculated with the program Raddose (Murray et al. 2005), taking into account the fluorescence correction. The 3600° were split into 20 sets (180° each), which were processed independently with XDS. Occupancies of the anomalous scatterers were derived from the anomalous difference maps calculated with the anomalous differences for each of the 20 data sets according to the method performed by Olieric et al. (2007).
Of course, we cannot exclude a possible influence on radiation damage of RNA sequence and crystallization conditions. However, in the present study, a great care was taken to compare heavy atoms (Br, I, or Se) with very similar parameters. For instance bromo-, iodo-, and seleno-labeled DIS RNA structures were compared only for the same RNA sequence, and only when crystallized in highly similar conditions, thus ruling out any influence of buffer, salts or RNA construct. In addition, results obtained for the Se-labeled DIS RNA were very comparable to those with Se-labeled SRL RNA crystallized in different conditions, and we are therefore confident in the presented results. In addition, because a potential influence of crystal environment on the site-specific radiation damage was suspected in some studies (Ramagopal et al. 2005), only atoms with low accessible surface area (ranging from 1 to 13 Å2 with a 1.4 Å probe radius) were considered in the present study. However, since other studies (Burmeister 2000; Ravelli and McSweeney 2000; Fioravanti et al. 2007) are in contradiction with a significant effect of the solvent accessibility, we are quite confident that our results can be generalized for most cases.
SUPPLEMENTAL MATERIAL
Supplemental material can be found at http://www.rnajournal.org.
ACKNOWLEDGMENTS
We thank Marc Schiltz for his support during RIP-phasing trials of the DIS structure. This work was supported by the Agence Nationale de Recherche sur le SIDA (ANRS), the French Research Agency (ANR; project PCV07-187047), and the Austrian Science Fund (FWF; project P17864).
Footnotes
-
↵6 Present address: Architecture et Réactivité de l'ARN, Université de Strasbourg, CNRS, IBMC, 15 rue René Descartes, F-67084 Strasbourg, France.
-
Reprint requests to: Eric Ennifar, Architecture et Réactivité de l'ARN, Institut de Biologie Moléculaire et Cellulaire, Université Louis Pasteur/CNRS UPR 9002, 15 Rue René Descartes, F-67084 Strasbourg, France; e-mail: e.ennifar{at}ibmc.u-strasbg.fr; fax: 33-3-88602218.
-
Article published online ahead of print. Article and publication date are at http://www.rnajournal.org/cgi/doi/10.1261/rna.1499309.
-
- Received December 5, 2008.
- Accepted January 9, 2009.
- Copyright © 2009 RNA Society








