
m6A methylation inhibits recruitment of the Dand5 3′UTR to the left–right determinant Bicc1
- 1Ecole Polytechnique Fédérale de Lausanne (EPFL) SV ISREC, CH-1015 Lausanne, Switzerland
- 2Department of Molecular Biology, Science III, University of Geneva, 1211 Geneva 4, Switzerland
- Corresponding author: daniel.constam{at}epfl.ch
-
Handling editor: Maria Carmo-Fonseca
Abstract
In vertebrates, left–right (LR) asymmetry is specified by asymmetric decay of Dand5 messenger RNA (mRNA) mediated by the recruitment of the BicC family RNA binding protein 1 (Bicc1). Besides regulating organ laterality, Bicc1 is required to prevent cystic dilations in renal tubules and in pancreatic and bile ducts. However, validated target mRNAs are sparse in number, and how their binding to Bicc1 is regulated remains poorly understood. Bicc1 recruitment to Dand5 transcripts requires a conserved AGACGUGAC motif in the 3′UTR. Here, we report an N6-methyladenosine (m6A) in this sequence that disrupts binding to Bicc1 K homology (KH) domains in vitro, in stark contrast to IGF2BPs and FMR1, where m6A promotes RNA recognition by KH domains. We discuss the possible implications of this finding for LR axis formation and for a related role of Bicc1 in regulating specific target mRNAs in the kidney.
Keywords
INTRODUCTION
Reversible modification of mRNAs by N6-adenosine methylation (m6A) is highly prevalent and important in health and disease to dynamically regulate the recruitment of sequence-specific proteins involved in splicing, translation, and decay (Dominissini et al. 2012; Meyer et al. 2012, 2015; Du et al. 2016). The function of m6A methylation and its regulation depend on specific proteins known as m6A writers, readers, and erasers (Shi et al. 2019). In mammals, the majority of m6A methylation is deposited by the METTL3–METTL14 methyltransferase complex (Liu et al. 2014), within a loosely defined DR(m6A)CH (D = G/A/U, R = G/A, H = A/U/C) motif, while the demethylases FTO and ALKBH5 are the main erasers needed for m6A removal. The third group called m6A readers comprises several protein families, including the YT521-B homology (YTH) domain family, the Igf2 mRNA binding proteins (IGF2BP), heterogeneous nuclear ribonucleoproteins (HNRNPs), and eukaryotic initiation factor 3 (eIF3) (Theler et al. 2014; Alarcón et al. 2015; Meyer et al. 2015; Huang et al. 2018). Different readers can sometimes functionally overlap or antagonize each other. For instance, YTHDF proteins promote mRNA degradation in a redundant manner (Lasman et al. 2020; Zaccara and Jaffrey 2020), whereas IGF2BPs inhibit mRNA decay and facilitate translation (Huang et al. 2018; Hao et al. 2020). At the tissue level, m6A can promote or diminish the growth of various types of human cancers (Liu et al. 2019; Huang et al. 2020). In addition, m6A methylation regulates essential aspects of embryonic development, such as germ cell formation, epigenetic reprogramming during maternal–zygotic transitioning, embryo implantation, and left–right (LR) asymmetry (Aguilo et al. 2015; Chen et al. 2015; Geula et al. 2015; Hsu et al. 2017; Xu et al. 2017; Kim et al. 2021; Wang et al. 2023).
In vertebrates, LR asymmetry is specified during early embryogenesis by cells of the LR organizer that extend long cilia to generate or sense, respectively, a leftward flow of extracellular fluid (Nonaka et al. 1998; Tabin and Vogan 2003; Hirokawa et al. 2006). To produce a flow that can break bilateral symmetry, it suffices if only two of these cilia acquire motility (Shinohara et al. 2012). However, during normal development, cilia motility is induced in a majority of cells at the center of the LRO by a process that requires FTO-mediated m6A demethylation to increase the mRNA stability and hence the protein levels of the transcription factor Foxj1 (Brody et al. 2000; Stubbs et al. 2008; Yu et al. 2008; Osborn et al. 2014; Kim et al. 2021). In other tissues, FTO and Foxj1 also promote cilia lengthening, although this function is dispensable in LRO cells, possibly reflecting functional overlap with other factors (Bonnafe et al. 2004; Cruz et al. 2010; Osborn et al. 2014). In the murine LRO, which is also known as the ventral node (Shinohara and Hamada 2017), flow stimulates immotile cilia at the periphery to activate the cation channel polycystin-2 (PC2) encoded by the polycystic kidney disease-2 (Pkd2) gene (Pennekamp et al. 2002; Field et al. 2011; Yoshiba et al. 2012; Mizuno et al. 2020; Djenoune et al. 2023; Katoh et al. 2023). In turn, the increased calcium flux mediated by PC2 accelerates the decay of the mRNA encoding Dand5, a secreted antagonist of the TGF-β family member Nodal, thereby allowing Nodal to signal on the left side and to specify left-sided patterning (Marques et al. 2004; Schweickert et al. 2010; Kawasumi et al. 2011; Maerker et al. 2021; Minegishi et al. 2021; Katoh et al. 2023).
Recent studies in mouse and Xenopus revealed that flow-induced repression of Dand5 mRNA on the left side of the LRO depends on a synergism of Dicer with Bicc1, and on the Ccr4–Not deadenylase complex (Maerker et al. 2021; Minegishi et al. 2021). Both pkd2 and dicer are also required for left-sided inhibition of dand5 in the zebrafish LRO, in keeping with a possible function for Bicc1 in promoting miRNA access to target mRNAs (Piazzon et al. 2012; Maerker et al. 2021; Minegishi et al. 2021). Moreover, mouse Bicc1 physically interacts with the Dand5 mRNA in the proximal 3′UTR, as initially described in Xenopus (Zhang et al. 2013). Specific Bicc1 binding to this and other target mRNAs is mediated by a tandem repeat of three ribonucleoprotein K homology (KH) domains containing the conserved sequence GXXG, where at least one X is a basic amino acid. This basic residue interacts with the phosphate backbone of single-stranded RNA to orient the free nucleobases of short 4 nucleotide (nt) motifs toward sequence-specific binding grooves on the KH domains (Piazzon et al. 2012; Zhang et al. 2014; Nicastro et al. 2015; Dowdle et al. 2019; Minegishi et al. 2021). In Bicc1, the KH1 and KH2 domains recognize the YGAC and GACR motifs, respectively, which are found in tandem in the sequence AGACGUGAC of the proximal Dand5 3′UTR (Minegishi et al. 2021). Interestingly, both AGAC and UGAC fit with the consensus sequence DRACH that is frequently methylated on the central adenosine by m6A writers (Linder et al. 2015), raising the question as to whether m6A methylation regulates Bicc1 binding.
Here, analysis of public epitranscriptomic and gene expression data showed that m6A writers and readers are expressed at the node, and that the AGACGUGAC sequence in the Dand5 3′UTR can be methylated on the second adenosine. Molecular modeling and in vitro validation by electrophoretic mobility shift assay (EMSA) and GST pull-down revealed that m6A modification of the AGACG motif suppresses Bicc1 binding to the Dand5 3′UTR. Our finding that m6A methylation of the Dand5 3′UTR inhibited Bicc1 recruitment further enlarges the repertoire of mechanisms to dynamically regulate when and how efficiently this RNA-binding protein will dampen the expression of specific targets during LR axis formation and in other tissues such as kidney, liver, and pancreas.
RESULTS
A single m6A peak in the Dand5 mRNA maps to the Bicc1 binding site in the proximal 3′UTR
Due to the absence of public data on m6A methylation of mRNAs in LR organizer cells of any model organisms, we investigated whether Dand5 mRNA is m6A methylated in an epitranscriptomic data set from mouse testes where Dand5 is highly expressed (Mendel et al. 2021). We found a single peak in the 3′UTR that overlapped the Bicc1 recognition motif (Fig. 1). The width of this peak indicates the presence of two methylated sites, including the GACG sequence that was recently identified as a binding site for the KH2 domain of Bicc1 (Minegishi et al. 2021). This observation suggests that the 3′UTR of Dand5 can be m6A methylated at a position critical for its post-transcriptional regulation by Bicc1.
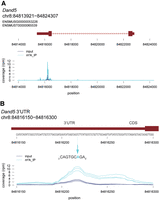
Identification of an m6A methylation site in Dand5 3′UTR. Coverage of m6A-enriched reads along Dand5 mRNA (A) and its 3′UTR (B) in mouse testes. Light and dark blue curves show the m6A reads in m6A immunoprecipitates and in corresponding input fractions, respectively. The normalized coverages (in reads per million [rpm]) from three biological replicates are plotted separately. A methylated adenosine in position 3 of the conserved AGACGUGAC motif of the Dand5 3′UTR is marked by a blue arrow. Data were retrieved from m6A-IP-seq data set generated from adult mouse testes (Mendel et al. 2021).
m6A writers and readers are coexpressed with Dand5 and Bicc1 in murine node cells
To investigate if m6A writers or readers are expressed in node cells, we mined public scRNA-seq data from mouse embryos at embryonic day (E) 7.5, when the ventral node appears (Scheibner et al. 2021). In the axial mesendoderm population, Foxj1 and Dand5 were used as markers to define peripheral node cells (Fig. 2A,B; Marques et al. 2004; Cruz et al. 2010). Among 278 axial mesendoderm cells, 268 were Foxj1+, including 57 which also expressed Dand5. In addition, the Foxj1+ Dand5+ population expressed the node markers Bicc1 and Nodal, as expected (Fig. 2C; Supplemental Fig. S1A). Interestingly, all populations clearly transcribed the widely expressed m6A writers Mettl3, Wtap, and Mettl14, and the m6A reader families of Igf2bp and YTHDF, as well as the m6A eraser FTO (Fig. 2A,C; Supplemental Fig. S1B–D). These observations suggest that components of the m6A apparatus are broadly transcribed during symmetry breaking, including in Dand5-producing node cells.
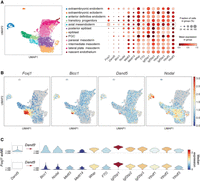
Murine node cells transcribe m6A writers, readers, and erasers. (A) UMAP plot colored by cell population of mouse embryos at embryonic day (E) 7.5 (left). The relative expression of specific genes of interest in each of these populations is depicted by the dot plot to the right. Colors, expression levels; dot size, fraction of cells expressing the gene indicated on the x-axis. (B) UMAP plot showing the relative expression levels of Foxj1, Bicc1, Dand5, and Nodal across the cell populations in A. (C) Violin plots of the expression levels of m6A writers, erasers, and readers in Foxj1+ Dand5+ (n = 57) and in Foxj1+ Dand5− (n = 211) axial mesendoderm (axME) cells. Data were retrieved from a public scRNA-seq data set (Scheibner et al. 2021). Cells containing at least one unique molecular identifier (UMI) were considered to be positive for the transcript of interest.
Structure modeling predicts that m6A methylation diminishes binding of the AGACG motif to the Bicc1 KH2 domain
m6A methylation can increase or inhibit specific binding of mRNAs to proteins (Edupuganti et al. 2017). To evaluate the impact of m6A on Bicc1 binding to the Dand5 3′UTR, we first modeled the structure of the Bicc1 KH2 domain in complex with a bound AAGACG nucleotide sequence using the AlphaFold 3 algorithm (Fig. 3; Abramson et al. 2024). Alternative RNA sequences of variable length from nonbound regions of the Dand5 3′UTR were compared in parallel to test if the algorithm can predict the sequence-specific recognition of the expected GACG recognition motif independently of flanking nucleotides (Supplemental Fig. S2A). Interestingly, AlphaFold invariably predicted the accommodation of the 5′-GACG-3′ sequence in the expected orientation within the RNA-binding cleft, along with the upstream adenosine, which is conserved among mammals and in Xenopus (Minegishi et al. 2021). Based on these models, the KH2 domain appears to recognize the GACG motif in the extended AGACG form, to which we will refer hereafter. After additional refinement by molecular dynamics (MD) simulation, we analyzed the AlphaFold model of the KH2 domain in complex with the AGACG sequence in further detail (Fig. 3A). We found that Lys152 of the GXXG motif, as well as Arg134 and Lys158, angle their side chains toward the phosphates of the AGACG RNA sequence (Supplemental Fig. S2), such that its nucleobases form up to 12 H-bonds with the sequence-specific recognition groove of KH2 (Fig. 3B; Supplemental Fig. S2C–H).
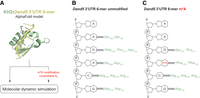
AlphaFold structure modeling of Bicc1 KH2 domain binding to the RNA sequence AGACG with and without m6A methylation. (A) Strategy to model the complex between the KH2 domain and the AGACG motif and to evaluate the impact of m6A methylation. (B,C) Diagram summarizing the predicted hydrogen bonds (H-bonds) between the KH2 domain and the nucleobases of the unmodified RNA hexamer sequence (B) and its m6A-methylated form (C) after 5 psec of MD simulation. Asterisks (*) indicates amino acids forming H-bonds via their main chain.
To model the effect of RNA methylation, we first repeated the prediction with an AGm6ACG motif. We found that all five models proposed by AlphaFold shifted the AGACG sequence by −1 position compared to the unmodified molecule (Supplemental Fig. S2B), strongly suggesting that the m6A modification perturbs binding to the KH2 domain. Therefore, we decided to further compare the nonmethylated Bicc1 KH2:AGACG model structure to the known interaction between an m6A-methylated GAC motif (AGm6ACU) and the structurally related KH2 domain of the fragile X messenger ribonucleoprotein 1 (FMR1) (Arguello et al. 2017; Edupuganti et al. 2017; Zhang et al. 2022). We found that FMR1 is predicted to bind m6A-methylated GAC in a predominantly hydrophobic groove that is wide and shallow (Supplemental Fig. S2I). In contrast, in Bicc1, this same RNA sequence fits into a narrow cleft, with the unmethylated adenosine anchored in a deep pocket characterized by a hydrophobic bottom and polar edges which would be unlikely to tolerate the steric bulk and hydrophobicity of a methyl group.
Methylation of amino groups also decreases their polarity and hence their ability to donate hydrogens to H-bonds. To test the impact on H-bonds between the AGACG sequence and Bicc1, the central adenosine was N6-methylated in silico in the minimized AlphaFold model of its complex with Bicc1 (Fig. 3A). MD simulation revealed that m6A triggers a conformational rearrangement, leading to the loss of five out of 10 H-bonds between specific amino acid side chains and interacting nucleobases, including the ones that stabilize the initial adenosine (Fig. 3C). These observations predict that m6A modification hinders binding of the AGACG motif to the Bicc1 KH2 domain rather than enhancing it.
m6A methylation of the AGACG motif from the Dand5 3′UTR abrogates specific binding to Bicc1 KH domains in vitro
To test the predicted 3D structure model, we monitored RNA binding of Bicc1 in the presence of increasing concentrations of short competitor RNAs with and without m6A using electrophoretic mobility shift assays (EMSA). In this assay, a shift in RNA mobility induced by Bicc1 binding can be visualized by preannealing the target RNA to a short complementary fluorescent probe (Minegishi et al. 2021). To preassemble these fluorescently tagged ribonucleoprotein complexes, fluorescent probe and a GST fusion of the RNA-binding Bicc1 KH domains 1–3 (GST-KH) were incubated with in vitro–transcribed nucleotides 66–110 of the Dand5 3′UTR (Fig. 4A). To estimate a dissociation constant of the resulting fluorescent RNPs, we compared their electrophoretic mobility in native polyacrylamide gels in the presence or absence of increasing amounts of nonfluorescent 45 nt competitor RNA with or without an m6A modification (Fig. 4B). The half maximal inhibitory concentration (IC50) of unmodified competitor RNA was 181 nM, confirming high affinity binding (Fig. 4C,D). As described previously, mutating the AGACG motif of the synthetic competitor RNA to AAAGG abolished binding, confirming sequence specificity (Minegishi et al. 2021). Importantly, m6A methylation of the synthetic RNA similarly abolished competitive binding (IC50 ≫ 1 µM). As a positive control, we tested if m6A-methylated competitor binds to recombinant YTH domain (residues 401–554) of the m6A reader YTHDF2 carrying a FLAG epitope (Supplemental Fig. S3A). Analysis by EMSA showed that FLAG-tagged YTH domain shifted the mobility of synthetic Dand5 3′UTR66–110 RNA only if the latter was m6A methylated, confirming the quality of m6A methylation and that it specifically favors binding to YTH while inhibiting binding to the Bicc1 KH domains (Supplemental Fig. S3B).
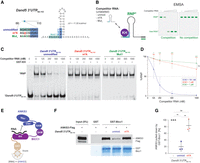
m6A methylation of the AGACGUGAC motif inhibits the binding of Bicc1 to the proximal Dand5 3′UTR. (A) In vitro–transcribed Dand5 3′UTR66–110 RNA and variant versions used as competitors in EMSA assays to assess the binding of Bicc1. The secondary structure and minimum free energy (ΔG) of the Dand5 3′UTR66–110 RNA were predicted by the RNAfold server (http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi). (B) Principle of the competitive EMSA experiment: increasing amounts of unlabeled competitors were added to preassembled complexes between recombinant GST-KH fusion protein and Dand5 3′UTR66–110 RNA marked with a fluorescent probe (green bold line) complementary to an extrinsic sequence used as a 3′ tag (black bold line). Destruction of the fluorescent ribonucleoparticle (*RNP) by the competitor RNA indicates specific interaction. (C) EMSA analysis of competitive binding of Bicc1 KH domains to synthetic transcripts of unmodified (unmod.) or m6A-methylated (m6A) Dand5 3′UTR66-110 against the fluorescent unmodified Dand5 3′UTR66–110 RNA probe. A Bicc1 binding null mutant (Mut1) (Minegishi et al. 2021) was used as a negative control to show absence of competition. (D) Graph showing the % of residual RNP relative to the indicated competitor RNA concentration, and the half maximal inhibitory concentrations (IC50) of each competitor. P-values (asterisks) above the curves refer to differences between each variant versus the unmodified RNA, whereas differences between variants ([n.s.] not significant) are marked by brackets. (E) Cartoon depicting the GST pull-down experiment presented in F and G. (F) Western blot analysis of ANKS3-FLAG from HEK293T cell extracts before (input) and after pull-down by recombinant GST-Bicc1 preincubated or not with saturating amounts of unmodified or m6A Dand5 3′UTR66–110 RNA. Coomassie blue staining of GST-Bicc1 protein retained by the beads (bottom panel) is shown below. (G) Histogram showing the amounts of pulled down ANKS3-FLAG shown in panel F experiment. Values were normalized to the control without target RNA (100%). Data are means ± SD from three independent experiments. (ns) Nonsignificant, (*) P < 0.05, (**) P < 0.01, (***) P < 0.001 (Student's t-test).
Specific binding of Bicc1 KH domains to mRNA is antagonized by a competing interaction of KH domains with the ankyrin and sterile α motif (SAM)-containing 3 protein (ANKS3) (Rothé et al. 2018, 2023a,b). Therefore, if m6A methylation blocks Dand5 3′UTR binding to Bicc1, we reasoned that it should also hinder the competition of this RNA against ANKS3. To test the impact of RNA methylation on ANKS3 binding, ANKS3-FLAG was pulled down from a HEK293T cell extract using GST-Bicc1 (Fig. 4E). As described previously, while recombinant GST-Bicc1 beads pull down ANKS3-FLAG in the absence of competitor RNA, this interaction is severely impaired if the beads are preincubated with an excess of synthetic Dand5 3′UTR66–110 RNA (Fig. 4F,G). Interestingly, m6A methylation abolished competition of the AGACG-containing RNA with ANKS3. These data further corroborate the conclusion that m6A methylation of the first GAC motif in the AGACGUGAC sequence of the Dand5 3′UTR inhibits Bicc1 binding rather than increasing it.
DISCUSSION
Specification of organ laterality in vertebrate embryos depends on the timely decay of Dand5 mRNA mediated by the proximal 3′UTR and its association with Bicc1 in LR organizer cells. Here, we found that murine LR organizer cells express several m6A writers and readers and that the conserved sequence AGACGUGAC which recruits the Dand5 3′UTR to Bicc1 can be m6A methylated on the first GAC motif. Three-dimensional structure predictions and their validation by EMSA and GST pull-down assays revealed that m6A modification at this position abolishes Bicc1 binding, likely by altering H-bond formation. Our finding that m6A methylation and Bicc1 binding of the proximal Dand5 3′UTR are mutually exclusive suggests that m6A methylation may control organ laterality in part by modulating the recognition of Dand5 mRNA by Bicc1.
Implication of m6A methylation in early development and embryo lateralization
Asymmetric expression of the Dand5 homolog Coco-1 in the LR organizer of Xenopus embryos has recently been shown to depend on the m6A eraser FTO (Kim et al. 2021). Mechanistically, FTO was shown to counteract m6A methylation of Foxj1 mRNA by the Mettl3–Mettl14 complex to thereby prevent destabilization of these transcripts by the m6A reader protein Ythdf2. In turn, increased Foxj1 protein synthesis drives the expression of genes required for motile cilia formation in LR organizer cells. Mettl3 is essential already before gastrulation for the transitioning of epiblast cells from naive to primed pluripotency (Geula et al. 2015). Consequently, Mettl3−/− embryos die before LR axis specification. Our analysis of public scRNA-seq data showed widespread expression of both m6A writers and readers during early embryogenesis, including in node cells marked by the coexpression of Foxj1 and Dand5 (Fig. 2), consistent with dynamic regulation of gene expression by m6A methylation during embryo lateralization.
m6A methylation abrogates specific binding of the Dand5 3′UTR to Bicc1 KH domains
m6A methylation can inhibit or promote binding of RNAs to specific interacting proteins, depending on the context. For instance, RRM and RGG domains of the stress granule protein G3BP1 bind newly transcribed mRNAs via nonmethylated GGACU motifs to prevent their recruitment to the destabilizing m6A reader YTHDF2 (Arguello et al. 2017; Edupuganti et al. 2017). In contrast, m6A is generally thought to enhance RNA recognition by KH domain-containing proteins, as demonstrated for FMR1 (Zhang et al. 2022) and the IGF2BP family (Huang et al. 2018). Here, pull-down by recombinant GST-Bicc1 and competitive EMSA with GST-KH domains showed that m6A methylation of the genetically validated target Dand5 on a single adenosine blocked binding to the Bicc1 KH domains in vitro. In keeping with this observation, 3D structure modeling suggested that m6A modification of the AGACG motif in the Dand5 3′UTR disrupts critical polar interactions with the KH2 domain. To our knowledge, the KH2 domain of Bicc1 is the first KH domain known to be repelled by m6A methylation of its recognition motif.
Previous analysis of the Dand5 3′UTR using the RNAalifold algorithm predicted that the UGAC motif of the Bicc1-binding AGACGUGAC sequence is part of a fairly conserved stem–loop structure (Minegishi et al. 2021). According to this model, only the AGACG motif is single-stranded by default and capable of initiating binding to the Bicc1 KH2 domain, whereas a subsequent interaction of the adjacent UGAC motif with the KH1 domain can be expected to facilitate melting of the adjacent stem (Fig. 4A). Our finding that inhibition of KH2 recruitment to AGACG by m6A abolished Bicc1 binding suggests that the energy cost to open the stem and expose the UGAC motif by the KH1 domain alone is too high. In addition, m6A methylation may inhibit RNA binding to Bicc1 by synergizing with the physiological antagonist ANKS3 (Rothé et al. 2023b) and/or by recruiting a specific m6A reader. An interesting question to address in the future is whether the expression of specific m6A readers such as IGF2BPs at the node is necessary to increase the stability and/or the translation of m6A-modified Dand5 transcripts. Moreover, Bicc1 is exclusively cytoplasmic, while the methyltransferase activity of the METTL3–METTL14 complex is confined to the nucleus (Lin et al. 2016). Given this differential compartmentalization, Bicc1 unlikely shields bound mRNAs from m6A modification, as proposed for G3BP1 (Edupuganti et al. 2017). Instead, transcripts that are methylated in the nucleus may only bind cytoplasmic Bicc1 if they are de-methylated by m6A erasers, such as FTO or ALKBH5.
Implications of Dand5 3′UTR m6A methylation for LR asymmetry
Nodal protein produced at the node is thought to diffuse to the left side mesoderm where it binds complexes of type I and II activin receptors to activate Smad2 and Smad3 transcription factors. In turn, Smad2/3 signaling mediates a feed-forward loop to induce Nodal expression also within left side mesoderm, as well as the transcription factor Pitx2 that governs asymmetric organ morphogenesis (Shiratori and Hamada 2014). In contrast, to terminate Nodal autoinduction, Smad2/3 activates the feedback inhibitors Lefty-2 and (at least in chick) Dand5 (Tavares et al. 2007). Up to eight potential Smad3-binding CAGAC motifs are also found within the 100 bp flanking the Dand5 transcription start site in mammals (Supplemental Fig. S4). Smad2/3 can influence the fate of nascent transcripts of at least some target genes by mediating cotranscriptional recruitment of the m6A methyltransferase complex (Bertero et al. 2018; Guo et al. 2025). Future studies should investigate whether Smad2/3 signaling thus protects newly synthesized Dand5 transcripts against Bicc1 binding to prevent precocious Dand5 mRNA decay prior to the onset of nodal flow. In Xenopus, Bicc1 is also recruited to an AGACA-containing sequence in the 3′UTR of the Nodal co-receptor Cripto (Zhang et al. 2014), raising the question if that interaction is similarly regulated by m6A methylation.
m6A methylation of mRNAs in the kidney
In kidneys, loss of Bicc1 leads to the accumulation of cAMP and the development of fluid-filled cysts reminiscent of polycystic kidney diseases (PKD) (Wallace 2011; Piazzon et al. 2012; Rothé et al. 2020). In line with this observation, a known function of Bicc1 is to curb the translation of adenylate cyclase 6 (Adcy6) mRNA (Piazzon et al. 2012). However, a complete list of physiological target mRNAs in whole kidneys or in other mammalian tissues is elusive. Additional candidate targets include c-Myc mRNA, which is bound and repressed by Drosophila BicC, preventing cystic dilations in malpighian tubules akin to the cysts induced by overexpressed c-Myc in mouse kidneys (Gamberi et al. 2017; Cai et al. 2018). Interestingly, the mRNAs encoding vasopressin receptor-2 upstream of cAMP and c-Myc depend on Mettl3-mediated m6A methylation for efficient translation and potentiation of cyst formation in mouse kidneys (Ramalingam et al. 2021). In addition, METTL3 promotes acute kidney injury (AKI) through m6A methylation and stabilization of the mRNA encoding the proinflammatory TGF-β–activated kinase 1 binding protein 3 (TAB3) (Wang et al. 2022). Future studies should investigate whether the access of Bicc1 to a specific pool of mRNAs is restricted by m6A methylation during the development and maintenance of renal tubules.
MATERIALS AND METHODS
Analysis of public sequencing data
The deep sequencing data set used in this study (GEO: GSE146873) has been described previously (Mendel et al. 2021). The biological replicates of input samples are RR726 (GSM4408144), RR727 (GSM4408145), and RR728 (GSM4408146). The biological replicates of m6A-IP samples are RR732 (GSM4408150), RR733 (GSM4408151), and RR734 (GSM4408152).
Analysis of public single-cell RNA sequencing data
An existing scRNA-seq data set from mesendoderm cells of mouse embryos at postnatal day E7.5 was deposited on Gene Expression Omnibus by Scheibner et al. (2021) (GEO: GSE162534). The original analysis pipelines of these authors (available at https://github.com/theislab/gastrulation_analysis) were adapted to analyze the expression of genes of interest in Foxj1+ cells. Briefly, we used their batch-corrected and annotated data set with logCPM-normalized, which includes non-batch-corrected values as raw data. Whenever a gene was represented by at least one unique molecular identifier, it was considered to be expressed in that cell.
3D modeling and MD simulation
Complex structure predictions were conducted with AlphaFold3 (https://alphafoldserver.com/). The presented structures are the top-ranked predictions with predicted Template Modeling (pTM) scores above 0.5 and interface predicted Template Modeling (ipTM) scores above 0.8, as recommended by AlphaFold. The m6A methylation of the adenosine in position 2 of the GACG motif was generated on ChimeraX (Pettersen et al. 2021). The best models were tested by MD simulation on ChimeraX. Protein–RNA complexes were simulated in water and subjected to energy minimization with steepest-descent method, equilibration and MD simulation during 5 psec using the AMBER ff14SB force field in the isobaric–isothermal ensemble, with the pressure being maintained at 1 atm and the temperature at 298 K. Final models were rendered in the PyMOL program (Schrödinger, Inc.).
Cell culture
The HEK293T (CRL-11268) cell line was purchased from ATCC and cultured in DMEM (Sigma) supplemented with 10% FBS (Sigma), 1% GlutaMAX (Thermo Fisher Scientific), and 1% gentamicin (Thermo Fisher Scientific). The pCMV6-Entry::ANKS3-FLAG plasmid (Rothé et al. 2018) was transfected using the jetPEI transfection reagent (Polyplus Transfection) according to the manufacturer's instructions.
Purification of GST fusion proteins
Fusions of GST with Bicc1 KH domains in plasmid pGEX-1λT (Piazzon et al. 2012) or with full-length Bicc1 in plasmid pGEX-6p1 (Rothé et al. 2023b) were expressed in Escherichia coli BL21 (Novagen) and purified using glutathione-sepharose 4B (GE Healthcare) as described previously (Piazzon et al. 2012). Purification was carried out in a buffer consisting of 50 mM Tris-HCl (pH 8), 200 mM NaCl, and 1 mM dithiothreitol (DTT).
In vitro transcription
The template for SP6 RNA polymerase-directed synthesis of the 3′-tagged Dand5 3′UTR66–110 was obtained by PCR amplification from the plasmid pEFBOS::Dand5-3′UTR using the 5′-ATTTAGGTGACACTATAGAAGAAAGACGTGACCTGAATGAT-3′ and 5′-CCTGAGCCTGTTGCCCAGACATTCTGCTCTCTTGACCCG-3′ primers. Note that the complementary 5′-TGTCTGGGCAACAGGCTCAGG-3′ sequence was introduced as a 3′ tag via the reverse primer. After agarose gel electrophoresis, the PCR amplicon was purified on NucleoSpin Gel and PCR Clean-up kit (Macherey-Nagel 740609). Five hundred nanograms of DNA was used for transcription with the SP6 RNA polymerase kit according to the manufacturer's conditions (Roche 10 810 274 001). After 2 h of incubation at 37°C, the transcripts were incubated for 1 additional hour in the presence of DNase I (Roche 04 716 728 001) in order to eliminate the SP6 DNA template. Finally, the transcripts were purified on Quick Spin Columns (Roche 11 274 015 001). The unmodified and m6A untagged Dand5 3′UTR66–110 competitors were synthetized by IDT (https://www.idtdna.com/page).
Fluorescent EMSA
The fluorescent DNA probes 5′-CTGAGCCTGTTGCCCAGAC-3′ and 5′-TTCTGCTCTCTTGA-3′, synthetized with a 5′-Dynomics 681 dye by Microsynth AG, were used to detect the 3′-tagged Dand5 3′UTR66–110 RNA and the untagged competitors, respectively. Before each experiment, 3′ tagged RNA and the fluorescent DNA probe were preannealed by denaturation (3 min at 98°C) and renaturation for 10 min at room temperature. One picomole of 3′-tagged RNA and 2.5 pmol of fluorescent probe were mixed for each condition. Complexes of fluorescent RNA:DNA duplex with recombinant GST-KH or with recombinant human YTHDF2401–554-FLAG (Active Motif 81103) were assembled in a final volume of 20 μL containing 10 mM Tris-HCl pH8, 100 mM KCl, 2.5 mM MgCl2, 2.5% glycerol, 1 mM DTT, and 1 µg of yeast tRNAs by incubation on ice for 30 min in the dark. For competition experiments, the recombinant GST-KH protein was used at a fixed concentration (200 nM). Increasing amounts of unlabeled competitor RNAs were then added to preassembled fluorescent complexes and incubated for another 30 min. The complexes were then resolved by electrophoresis on a 5% native polyacrylamide gel containing 45 mM Tris-Borate, 1 mM EDTA, and 2.5% glycerol. The fluorescence was detected using the Odyssey CLx Infrared Imaging System (LI-COR Biosciences).
Multiprotein complex and RNP reconstitution by GST pull-down
HEK293T cells cultured in 10 cm dishes were transfected with 2 μg of ANKS3-FLAG plasmid (OriGene RC223862). After 24 h, cells were collected and extracted in 20 mM Tris-HCl (pH 7.4), 2.5 mM MgCl2, 100 mM NaCl, 5% glycerol, 1 mM dithiothreitol (DTT), 0.05% Nonidet P-40 (NP-40), RNasin (Promega), phosphatase inhibitors (Sigma), and protease inhibitors (Roche), by passing them eight times through a syringe needle (no. 30). The cell extract was centrifuged twice at 10,000g for 5 min at 4°C and 5% set aside as a control for the “input.” In parallel, the synthetic RNA (100 pmol) was incubated with glutathione beads coated with GST-Bicc1 fusion protein (∼5–10 pmol) during 1 h at 4°C on wheel. To assess binding of preassembled RNPs or of GST-Bicc1 fusion alone to ANKS3, cleared extract corresponding to one-third of a 10 cm dish per binding assay was incubated for 2 h at 4°C with glutathione-sepharose 4B beads coated with GST alone (control), or with GST-Bicc1 fusion or RNPs. Upon washing four times for 5 min in 20 mM Tris-HCl (pH 7.4), 2 mM MgCl2, 200 mM NaCl, 1 mM DTT, and 0.1% NP-40, the retention of ANKS3-FLAG was analyzed by western blotting using the monoclonal mouse anti-FLAG M2 (Sigma F3165) and the Odyssey CLx Infrared Imaging System (LI-COR Biosciences). Binding of the GST-Bicc1 fusion was validated by Coomassie staining of eluted proteins. GST alone was included in all experiments as a specificity control but could not be retained due to its small size after the prolonged migration that was required to resolve the protein of interest at the top of the gel.
Quantification and analysis
EMSA and western blot signals were quantified using the Odyssey CLx scanner software, ensuring that all bands were below saturation. For EMSA, the levels of fluorescent RNPs were expressed as percentages normalized to their amounts in the control condition without competitor RNA. In pull-down experiments, the levels of ANKS3-FLAG that associated with GST-Bicc1 were expressed as percentages normalized to their amounts in the corresponding crude cell extracts (input) and relative to the control condition without competitor RNA. Statistics were calculated with Microsoft Excel software. Error bars represent standard deviations (SD). Student's t-test was used to calculate P-values, with P ≤ 0.05, P ≤ 0.01, or P ≤ 0.001 represented by 1, 2, or 3 asterisks, respectively.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
We would like to thank Dr. David Homolka (Geneva University) for help with figure preparations and Benjamin Seiple for reading the manuscript. This work was supported by the Swiss National Science Foundation, grant 310030 2127121/1.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080526.125.
-
Freely available online through the RNA Open Access option.
- Received April 15, 2025.
- Accepted June 21, 2025.
This article, published in RNA, is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Benjamin Rothé is the first author of this paper, “m6A methylation inhibits recruitment of the Dand5 3′UTR to the left–right determinant Bicc1.” As a staff scientist in the group of Professor Daniel Constam at EPFL (Lausanne), Benjamin studies the function of the RNA-binding protein Bicc1 and how it might be regulated by flow stimulation of primary cilia to prevent developmental laterality defects and polycystic kidney disease. Benjamin initially joined the Constam lab as a postdoc after completing his PhD in the laboratory of Dr. Christiane Branlant and Professor Bruno Charpentier (Université de Lorraine/CNRS), where he investigated the biogenesis of small nucleolar ribonucleoproteins (snoRNPs).
What are the major results described in your paper and how do they impact this branch of the field?
Mutations in the K homology (KH) domain-containing protein Bicc1 cause kidney cysts reminiscent of polycystic kidney disease (PKD), along with abnormal organ laterality linked to defective signaling by mechanosensory cilia. Over the past decade, we have uncovered a role for Bicc1 in mediating post-transcriptional regulation of specific target mRNAs. In the Dand5 3′UTR, we previously identified a Bicc1-binding motif that mediates accelerated mRNA decay downstream of flow-sensing cilia to specify organ laterality. Here, pull-down assays, competitive EMSA, and structure modeling revealed that m6A methylation of this motif inhibits Bicc1 binding. Thus, m6A may act as a rheostat to control Bicc1 access to only a fraction of Dand5 mRNA and possibly other transcripts. More broadly, our finding also raises the question of whether and how mechanotransduction by primary cilia and m6A methylation are functionally linked, an area that remains largely unexplored and may provide important insights into cellular signal transduction.
What led you to study RNA or this aspect of RNA science?
Early in my studies, I became fascinated by RNA, which struck me as the dark matter of the cell—an ancient and remarkably versatile molecule that still holds many secrets. Contributing to the endeavor of uncovering how cells utilize this intricate infrastructure feels like joining a genuine and compelling scientific quest.
During the course of these experiments, were there any surprising results or particular difficulties that altered your thinking and subsequent focus?
Our initial approach was motivated by the similarity between the Bicc1 binding site and the m6A consensus methylation motif. A surprise was that m6A methylation inhibited recognition by the KH domains of Bicc1, whereas previously characterized KH domain-containing proteins have been described as m6A readers. This result led us to reconsider how m6A modifications could act as a molecular gatekeeper to limit access of Bicc1 to only a fraction of a given transcript or, if there is more than one binding site, to perhaps reduce the number of options.
What are some of the landmark moments that provoked your interest in science or your development as a scientist?
There was no single defining moment; this is something I have always carried within me. Since childhood, I have been intrigued by fundamental questions such as “What is infinity?” or “Where does life come from?” Despite our apparent technological control, we live in a world full of mysteries. The attraction lies in the unknown—in what we do not yet understand or even see.
Are there specific individuals or groups who have influenced your philosophy or approach to science?
It is more of a scientific ideal than a direct influence on my research, but instinctively I think of the generation of physicists from the 1920s who laid the foundations of quantum mechanics. Their work marked a major turning point in the history of science. I have great respect and admiration for pioneers who dared to think differently and laid the groundwork for entirely new fields.








