
The modification landscape of Pseudomonas aeruginosa tRNAs
- Mariana D. Mandler1,
- Siddhardha S. Maligireddy1,
- Wilfried M. Guiblet1,
- Christina M. Fitzsimmons1,
- Kayla S. McDonald1,
- Delayna L. Warrell2 and
- Pedro J. Batista1
- 1Laboratory of Cell Biology, Center for Cancer Research, National Cancer Institute, National Institutes of Health, Bethesda, Maryland 20892, USA
- 2Laboratory of Molecular Biology, Center for Cancer Research, National Cancer Institute, National Institutes of Health, Bethesda, Maryland 20892, USA
- Corresponding author: pedro.batista{at}nih.gov
Abstract
RNA modifications have a substantial impact on tRNA function, with modifications in the anticodon loop contributing to translational fidelity and modifications in the tRNA core impacting structural stability. In bacteria, tRNA modifications are crucial for responding to stress and regulating the expression of virulence factors. Although tRNA modifications are well-characterized in a few model organisms, our knowledge of tRNA modifications in human pathogens, such as Pseudomonas aeruginosa, remains limited. Here, we leveraged two orthogonal approaches to build a reference landscape of tRNA modifications in Escherichia coli, which enabled us to identify similar modifications in P. aeruginosa. Our analysis supports a substantial degree of conservation between the two organisms, while also uncovering potential sites of tRNA modification in P. aeruginosa tRNAs that are not present in E. coli. The mutational signature at one of these sites, position 46 of tRNAGln1(UUG) is dependent on the P. aeruginosa homolog of TapT, the enzyme responsible for the 3-(3-amino-3-carboxypropyl) uridine (acp3U) modification. Identifying which modifications are present on different tRNAs will uncover the pathways impacted by the different tRNA-modifying enzymes, some of which play roles in determining virulence and pathogenicity.
Keywords
INTRODUCTION
RNA modifications have a substantial impact on gene expression, modulating the function of both coding and noncoding RNAs (Frye et al. 2018). Many RNA modifications were first identified on tRNA, which is known to have the highest frequency of modified nucleosides per RNA length, averaging eight modifications per tRNA molecule in bacteria and 13 in eukaryotes (Pan 2018; Zhang et al. 2022). Reflecting the importance of tRNA modifications in cellular function, about 1.5% of the proteins in the Escherichia coli proteome are necessary for the synthesis of the different tRNA nucleotide modifications (Björk and Hagervall 2014; de Crécy-Lagard et al. 2020). On tRNA transcripts, modifications are not uniformly distributed, and the function of a given modification depends on its location. Although modifications in the anticodon loop play an important role in translational fidelity, modifications in the tRNA D-arm and T-arm influence tRNA structural stability (Pan 2018).
In bacterial pathogens, tRNA modifications are important regulators of stress response and the expression of virulence factors (Koh and Sarin 2018). In Vibrio cholerae, the pathogen responsible for cholera, genes required for tRNA modification, including thiI, although dispensable for growth under laboratory conditions, are required for optimal bacterial growth in an animal model (Pritchard et al. 2014). The enzyme ThiI is responsible for the addition of 4-thiouridine (s4U) at positions 8, or 8 and 9, of bacteria and archaea (Dunin-Horkawicz et al. 2006; Boccaletto et al. 2022). This modification plays a unique role in prokaryote biology, as response to near-ultraviolet (UV) light is dependent on the presence of s4U in tRNAs (Ramabhadran and Jagger 1976; Thomas and Favre 1980; Thomas et al. 1981; Kramer et al. 1988). In addition, s4U has been shown to function cooperatively with other modifications to ensure that tRNAs fold properly to avoid clearance through the RNA degradosome in a tRNA quality control pathway (Kimura and Waldor 2019). In Pseudomonas aeruginosa, the tRNA-modifying enzyme MiaB is a positive regulator of the Type III secretion system and is required for induced cytotoxicity of human lung epithelial cells (Lin et al. 2022). MiaB catalyzes the final step in the synthesis of 2-methylthio-N6-isopentenyladenosine (ms2i6A), using N6-isopentenyladenosine (i6A) as a substrate (Esberg et al. 1999). The effect of MiaB on the Type III secretion system might be independent of tRNA modification levels (Lin et al. 2022). In multiple pathogens, the tRNA uridine 5-carboxymethylaminomethyl synthesis enzyme GidA plays a critical role in the Rhl-dependent quorum-sensing pathway. In a complex with MnmE, GidA catalyzes the formation of methyluridine derivates at the wobble position, critical for the decoding of NNG/A codons (Gupta et al. 2009; Moukadiri et al. 2009; Boccaletto et al. 2022). GidA homologs control the expression of virulence genes, with no impact on growth, by modulating posttranscriptional regulation of essential transcriptional activators (Durand et al. 2000; Kinscherf and Willis 2002; Sha et al. 2004; Cho and Caparon 2008). It has been proposed that mRNAs with a high proportion of codons that require methyluridine derivates at the wobble position for decoding are more sensitive to changes in the expression of the GidA enzyme (Lin et al. 2022). In P. aeruginosa, the response to oxidative stress is also dependent on tRNA modifications. Loss of the protein TtcA, responsible for the biosynthesis of 2-thiocytidine (s2C), tRNA methyltransferase TrmJ, responsible for 2′-O-methylcytidine (Cm), 2′-O-methyluridine (Um), and 2′-O-methyladenosine (Am), or TrmB, responsible for 7-methylguanosine (m7G) modification at position 46 results in hypersensitivity to oxidative stress (Jaroensuk et al. 2016; Romsang et al. 2018; Thongdee et al. 2019). The effect of TtcA, which is upregulated in response to oxidative stress, on gene expression occurs at the posttranscriptional level, and loss of this enzyme impacts virulence in a host model (Romsang et al. 2018). Loss of TrmB impacts translation of transcripts enriched with codons decoded by tRNAPhe(GAA) and tRNAAsp(GUC), including catalases that break down hydrogen peroxide, KatA and KatB. These examples highlight the role of tRNA modifications in the virulence of pathogenic bacteria (Thongdee et al. 2019).
P. aeruginosa, a Gram-negative bacterium, is a common environmental microorganism known, under specific circumstances, to become an opportunistic pathogen capable of infecting immunocompromised individuals (Lyczak et al. 2000). This microorganism, which is frequently found in multispecies infections, leverages multiple virulence strategies, and infects a wide range of hosts (Lyczak et al. 2000; Stover et al. 2000; Khare and Tavazoie 2015). Previous studies have explored RNA modifications at a global level in P. aeruginosa tRNAs by liquid chromatography–tandem mass spectrometry (LC–MS/MS) (Jaroensuk et al. 2016; Grobe et al. 2019) and identified modifications that are conserved between E. coli and P. aeruginosa, as well as modifications that are present exclusively in E. coli or P. aeruginosa (Jaroensuk et al. 2016; Grobe et al. 2019).
In this study, we aimed to map tRNA modifications in P. aeruginosa at nucleotide resolution and determine which modifications are present in each tRNA transcript. To achieve this, we used a combination of cDNA misincorporation and termination sequencing analyses to predict sites of tRNA modification with previously established methanethiosulfonate (MTS) chemistry to enrich for s4U-modified RNA (Supplemental Fig. S1A). Using E. coli as a reference, we predicted which tRNA modifications are conserved between the two organisms and assigned modification status to multiple tRNA positions in P. aeruginosa. In addition, we identified modification sites that are present in P. aeruginosa but not in E. coli, such as position 46 of tRNAGln1(UUG) and position U20B of tRNAGlu1(UUC), positions recently described to be modified with an acetylated form of N3-(3-amino-3-carboxypropyl)-uridine (acp3U) in V. cholerae (Kimura et al. 2020). Finally, we identified the P. aeruginosa homolog of TapT and demonstrated that the modification signature at position 46 of tRNAGln1(UUG) is dependent on the activity of this enzyme. It remains to be determined if acp3U at this position is further modified to acetylated acp3U (acacp3U). Altogether, the approach presented here offers the discovery of sites of tRNA modifications from understudied bacteria that are implicated in their virulence and pathogenesis.
RESULTS
To identify RNA modifications in P. aeruginosa tRNAs, we first built a reference modification landscape in the well-characterized model organism E. coli. To this end, we leveraged two orthogonal approaches: analysis of cDNA mutational signature and targeted, chemical enrichment of s4U-modified RNAs. During cDNA synthesis, the presence of modifications that interfere with the activity of the reverse transcriptase (RT) enzyme can result in the incorporation of mismatched bases in the cDNA or premature termination of cDNA synthesis. By examining these occurrences at each position on the tRNA, defined here as a mutational signature, we can effectively predict the presence of modifications (Cozen et al. 2015; Zheng et al. 2015; Gogakos et al. 2017; Richter et al. 2018; Schwartz et al. 2018; Vandivier et al. 2019; Kimura et al. 2020; Wang et al. 2021). Because of the high frequency of complex modifications found in the anticodon loop of tRNAs transcripts, cDNA products are often truncated. Consequently, read coverage for positions in proximity to the 5′ end, where s4U is found (Bommisetti and Bandarian 2022), may be restricted. To address this limitation, we used a strategy to isolate tRNAs through affinity purification targeting the s4U modification to determine levels of s4U modification (Duffy et al. 2015).
Generation of a cDNA mutational signature map for tRNA modifications in E. coli
To generate the reference landscape of tRNA modifications in E. coli, we analyzed tRNAs extracted from cells cultured in Luria–Bertani (LB) media or chemically defined media (CDM) and collected during exponential growth phase (optical density [OD]600 = 0.6). To identify both misincorporation and early termination events during cDNA synthesis, we adapted the library generation protocol described by Flynn et al. (2016). tRNAs were gel purified from a urea-polyacrylamide gel (UREA-PAGE) after ligation of an oligo adaptor to the 3′ end of total RNA. cDNA was generated with thermostable group II intron reverse transcriptase (TGIRT) (Mohr et al. 2013) and an oligonucleotide including universal molecular identifiers (UMIs), which allows for the accurate removal of PCR duplicate reads (Zarnegar et al. 2016). To ensure that truncated cDNA products are represented in the final library, the cDNA product was circularized and the final amplicon for sequencing was generated by PCR. Reads were mapped to a custom curated transcriptome, which includes annotated coding and noncoding RNAs. In this curated transcriptome, tRNAs that share 100% sequence identity were grouped together (Supplemental Table S1). Reads were mapped with Bowtie 2 (Langmead and Salzberg 2012) with the inclusion of the ‐‐local parameter to account for the addition of nontemplated nucleotides by TGIRT at the 3′ end of the cDNA (Qin et al. 2016). We limited our analysis to single mapped reads that include the CCA sequence at the 3′ end. The frequency of misincorporation and a termination score were calculated for each tRNA position, with the termination score determined using the formula (Depthi+1 − Depthi)/(Depthi+1) as previously described (Wang et al. 2021). Each analysis was performed on three biological replicates that show a high degree of correlation (Supplemental Fig. S1B). Consistent with previous studies (Cozen et al. 2015; Zheng et al. 2015; Gogakos et al. 2017; Richter et al. 2018; Schwartz et al. 2018; Vandivier et al. 2019; Kimura et al. 2020; Wang et al. 2021), we observe misincorporation, early termination, or a combination thereof (mutational signature) across all tRNAs in both culture media (Fig. 1A; Supplemental Fig. S2). In this study, we predict a tRNA position to be modified if it meets the criteria of having a read coverage equal to or >100 reads and exhibits a minimum of 5% misincorporation or termination, as previously described (Kimura et al. 2020). Applying these criteria, we identified a mutational signature for known modifications occurring on position 8 (Fig. 1A; Supplemental Figs. S2–S4A; Supplemental Table S2), the anticodon loop (Supplemental Figs. S3 and S5; Supplemental Table S4), and the variable loop (Supplemental Figs. S3 and S6; Supplemental Table S5). Additionally, we detect regions of misincorporation, early termination, or both in regions without known modifications, including the anticodon loop of certain tRNAs (Supplemental Table S2). It is worth noting that with our assay conditions, there are several types of modifications for which a consistent mutational signature is not observed (Supplemental Fig. S3; Supplemental Table S2). We observe that misincorporation at position 8 is dependent on the enzyme ThiI, confirming that the detected mutational signature is dependent on the presence of s4U modification (Supplemental Fig. S4A). Read coverage at position 8 is often low because of early termination of cDNA synthesis at the anticodon region. Therefore, we used MTSEA capture as an orthogonal method to characterize the s4U modification status of E. coli tRNAs. As previously demonstrated, MTSEA reacts with RNA extracted from wild-type E. coli, but not with RNA extracted from cells lacking the enzyme tRNA sulfurtransferase (ThiI) (Supplemental Fig. S4B,C; Mueller et al. 1998; Duffy et al. 2015). MTSEA enriched RNAs isolated from wild-type E. coli (strain BW25113) migrated at a size range corresponding to tRNA (Supplemental Fig. S4C; Favre et al. 1971; Emilsson et al. 1992). Among the tRNAs detected in this experiment, 13 and 11 were significantly enriched (log2 fold change [l2fc] > 0.6 and adjusted P-value [Padj] < 0.05) in RNA collected from cells cultured in LB and CDM, respectively (Supplemental Fig. S4D; Supplemental Table S3A,B). The MTSEA enrichment scores accurately reflected the misincorporation analysis, wherein frequency of misincorporation at position 8 correlated with MTSEA-dependent enrichment (Pearson correlation coefficient R = 0.53, and 0.55) (Supplemental Fig. 4E). These results suggest that both MTSEA capture and mutational analysis can be used to identify sites of s4U modification in tRNAs. The combination of targeted capture of s4U and analysis of mutational signature in the well-characterized model organism E. coli allowed us to build a reference map to assign RNA modifications to P. aeruginosa tRNAs.
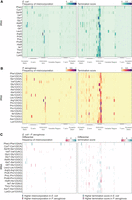
An Escherichia coli reference landscape for tRNA modifications. (A) Heatmap of average (from three independent samples) frequency of misincorporation (left) or termination score (right) at each position for E. coli tRNAs extracted from cells cultured in chemically modified media and collected at (OD)600 = 0.6. Only tRNAs with read coverage >100 at each position and with an identified isoacceptor homolog in Pseudomonas aeruginosa are represented. Different regions of the tRNA are represented at the bottom from the 5′ (left) to the 3′ end (right) of the tRNA. Darker lines indicate the loop in that region. (B) Heatmap of average (from three independent samples) frequency of misincorporation (left) or termination score (right) at each position for P. aeruginosa tRNAs extracted from cells cultured in chemically modified media and collected at (OD)600 = 0.6. Only tRNAs with read coverage >100 at each position and with an identified isoacceptor homolog in E. coli are represented. Different regions of the tRNA are represented at the bottom from the 5′ (left) to the 3′ end (right) of the tRNA. Darker lines indicate the loop in that region. The GtRNAdb names are used for P. aeruginosa tRNAs. The anticodon is shown in parentheses. (C) Heatmap of differential misincorporation frequency (left) or differential termination score (right) at each position for tRNAs with read coverage >100 at each position and with an identified isoacceptor homolog in E. coli and P. aeruginosa. Values are calculated as signal in E. coli subtracted of signal in P. aeruginosa. Sites with higher signal in E. coli are represented in blue gradient scale, whereas values with higher signal in P. aeruginosa are represented in a purple gradient scale.
Establishing a cDNA mutational signature map for tRNA modifications in P. aeruginosa
Previous studies have cataloged RNA modifications present in P. aeruginosa tRNAs by mass spectrometry (Jaroensuk et al. 2016; Grobe et al. 2019) and identified modifications that are conserved between P. aeruginosa and E. coli, as well as modifications that are found exclusively in each species. We aimed to assign globally conserved modifications between P. aeruginosa and E. coli using our data. For this comparison, tRNA transcripts were matched on the basis of both anticodon and sequence similarity (Supplemental Table S6). To assign conserved modifications to specific tRNA positions, we compared the pattern of misincorporation, early termination or both observed in tRNA sequencing libraries of both species (Fig. 1A,B; Supplemental Fig. S7A,B; Supplemental Table S7). Although the patterns of misincorporation and termination are generally similar, we identified some notable differences (Fig. 1C).
s4U modification is present in P. aeruginosa tRNAs
In P. aeruginosa, we detect a mutational signature at position 8, here defined as misincorporation of at least 5% (Figs. 1B and 2A), similar to what we observed in E. coli, which suggests conservation of s4U between the two species. However, we noted several exceptions: tRNAThrU(UGU) and tRNAAsnT(GUU) only have a signature in E. coli, whereas tRNAPhe1(GAA), tRNAGly1(UCC), tRNAArg1(ACG), and tRNAPro1(CGG) only have a signature in P. aeruginosa. As described for our E. coli experiments, coverage at this position in our data sets is less than100 reads for several tRNAs. Using MTSEA capture, we identified 13 significantly enriched tRNAs (l2fc > 0.6 and Padj < 0.05) (Fig. 2B; Supplemental Table S8). The mutational signature at position 8 of P. aeruginosa tRNAs correlates positively with enrichment after MTSEA capture (Pearson correlation coefficient R = 0.65) (Supplemental Fig. S8; Supplemental Table S9). Overall, we observe a similar pattern of s4U enrichment between E. coli and P. aeruginosa tRNAs (Fig. 2C; Supplemental Table S10), supporting the idea that this modification is conserved between the two species.

Conservation of s4U between Escherichia coli and Pseudomonas aeruginosa. (A) Heatmap of average (from three independent samples cultured in CDM) frequency of misincorporation at position 8 of isoacceptor homolog tRNAs in E. coli and P. aeruginosa. Only tRNAs with read coverage >100 at position 8 and with an identified isoacceptor homolog in both species are represented. The absence or presence of a signature based on a 5% cutoff of misincorporation is indicated on the right side of the heatmap with a circle mark. The key is listed below the heatmap. The criteria are based on the nature of the annotated modification. The GtRNAdb names are used for P. aeruginosa tRNAs. The anticodon is shown in parentheses. (B) Volcano plot representation of log2 fold change in abundance after MTSEA capture of tRNA from P. aeruginosa cells. Log2 fold change and adjusted P-value calculated with DESeq2. A dotted red line indicates the threshold used to define a tRNA as enriched (log2 fold change > 0.6). The color indicates if read coverage at position 8 is more than 100 in misincorporation frequency analysis. (C) Scatter plot representation of log2 fold change in abundance after MTSEA capture for isoacceptor homolog tRNAs from E. coli and P. aeruginosa. Log2 fold change and adjusted P-value calculated with DESeq2. Full circles indicate tRNAs that are significantly enriched in both samples.
Similarities and differences of tRNA modification between E. coli and P. aeruginosa in the anticodon region
When comparing misincorporation and early termination in the anticodon regions of E. coli and P. aeruginosa we note a significant conservation of mutational signatures for the majority of modifications. Importantly, all signatures at position 37 are conserved in both species, which includes the modifications for ms2i6A, 2-methyladenosine (m2A), 1-methylguanosine (m1G), cyclic N6-threonylcarbamoyladenosine (ct6A), and N6-methyl-N6-threonylcarbamoyladenosine (m6t6A) (Fig. 3). Interestingly, we find tRNAs in P. aeruginosa with a mutational signature that is conserved in the E. coli homologous tRNAs, for which no modifications have been annotated, such as for tRNAGly(CCC), tRNAGly(GCC), and tRNASer(GGA) (Fig. 3), which highlights the need to continue to annotate tRNA modifications, even in well-characterized organisms. As with our analysis of the s4U modification at position 8, we observe several exceptions. For example, E. coli tRNAGlnU(UUG) has a much stronger signature than the one observed in P. aeruginosa for the homologous tRNA, suggesting the presence of a different type of modification at position 37. In some cases, a drop in coverage at position 37 due to modifications that cause strong termination limits us from determining how well modifications are conserved at upstream positions, such as position 32. We also found examples in which a mutational signature is observed in both species at the same position, even though the nucleotide sequence is different: m2A at position 37 of tRNAHisR(GUG) and m1G at position 37 of tRNAArgX(CCG) in E. coli correspond to G and A in the P. aeruginosa homologous tRNAs. Finally, we also see conserved signatures at nucleotides which have different primary sequences and are not annotated as modified, such as position 32 of tRNAMetT(CAU) and tRNAMet1(CAU) (Supplemental Table S7). This observation implies that different modifications are present at these positions.
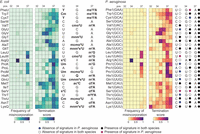
Conservation of tRNA modifications at the anticodon region. Heatmap of average (from three independent samples cultured in CDM) frequency of misincorporation and termination score between positions 32 and 38 in tRNA from Escherichia coli (left) and Pseudomonas aeruginosa (right). Only tRNAs with read coverage >100 in the anticodon loop and with an identified isoacceptor homolog in the two species are represented. Positions 32, 34, and 37, known to be modified, are annotated at the top of each heatmap. Annotated modifications for E. coli are represented to the right of the E. coli heatmap. The short name for each modification is used (Supplemental Table S11). Sequences in P. aeruginosa at positions 32, 34, and 37 are represented to the right of the P. aeruginosa heatmap. The GtRNAdb names are used for P. aeruginosa tRNAs. The anticodon is shown in parentheses. The absence or presence of a signature is indicated, based on a 5% cutoff of either misincorporation or termination for positions 32, 34, or 37. Circle marks indicate the conservation of mutational signature. The key is listed below the heatmap. The criteria are based on the nature of the annotated modifications.
Similarities and differences in tRNA variable region between E. coli and P. aeruginosa
The variable region of E. coli tRNAs features two well-defined modifications at positions 46 and 47—namely, m7G and acp3U. In E. coli, we detected a mutational signature for all sites annotated to be modified with both m7G and acp3U (Fig. 4). However, this mutational signature is absent in the majority of tRNAs annotated to be solely m7G modified, suggesting that the observed mutational signature is derived from the presence of acp3U. This is in line with previous observations that m7G alone lacks a mutational signature (Wang et al. 2021). In homologous P. aeruginosa tRNAs, we do not observe a mutational signature, suggesting that the absence of acp3U is conserved. The exceptions are tRNAArg1(CCG), tRNAAsn1(GUU), and tRNAArg1(UCU), in which the presence of a mutational signature suggests the presence of the acp3U modification in P. aeruginosa (Supplemental Table S7; Figs. 4 and 5). Notably, the tRNAArg1(UCU) nucleotide sequence differs from tRNAArgU(UCU) at position 47, where the U required for the acp3U modification is only present in P. aeruginosa. For tRNAs annotated to carry both m7G and acp3U in E. coli, we observe a conserved mutational signature for tRNAVal1(GAC), tRNAArg1(ACG), tRNAMet1(CAU), and tRNAIle1(GAU), suggesting acp3U is present in the P. aeruginosa homolog tRNAs. The absence of a mutational signature for tRNAPhe1(GAA) and tRNALys1(UUU) in P. aeruginosa implies that these tRNAs are not acp3U modified (Fig. 4). We observed a mutational signature for P. aeruginosa tRNAGln1(UUG) and tRNAfMet1(CAU), which are not annotated to be modified in E. coli. In both cases, the primary sequence is different between the two species (Fig. 4).

Conservation of acp3U and m7G Escherichia coli and Pseudomonas aeruginosa. Heatmap of average (from three independent samples cultured in CDM) frequency of misincorporation and termination score at positions 46 and 47 in tRNA from E. coli (left) and P. aeruginosa (right). Only tRNAs with read coverage >100 at position 46 and an identified isoacceptor homolog are represented. Known modifications for E. coli are represented to the right of the E. coli heatmap. The short name for each modification is used (Supplemental Table S11). Sequences in P. aeruginosa at positions 46 and 47 are represented to the right of the P. aeruginosa heatmap. The absence or presence of a signature is indicated based on a 5% cutoff of both misincorporation and termination for positions 46 or 47 by circle marks. The key is listed below the heatmap.

Modification sites at positions 46 or 47 that are lost in mutant strain of Pseudomonas aeruginosa PA14_17650. (A) LC–MS/MS quantification of acp3U from purified tRNA of wild-type P. aeruginosa versus PA14_17650 mutant for n = 2 replicates. Statistical significance was determined by Student's t-test. (B) Heatmap of average (from three independent samples cultured in CDM) frequency of misincorporation and average termination score at positions 46 and 47 in tRNA of P. aeruginosa from wild-type (left) and PA14_17650 mutant (right). Only tRNAs with read coverage >100 at position 46 are represented. The GtRNAdb names for P. aeruginosa tRNAs are listed on the left, with the anticodon shown in parentheses. The short name for the modification is shown at top right column (Supplemental Table S11). The triangle symbol labeled on the right of the heatmap indicates whether a signature is dependent PA14_17650, wherein the signature is lost in the mutant. The circle marks labeled on the right of the heatmap are color-coded and indicate conservation in Escherichia coli (key listed below the heatmap). The criteria used is based on a 5% cutoff of both misincorporation and termination for either position 46 or 47.
The variable region mutational signature is dependent on PA14_17650
To confirm that the mutational signature we observe at positions 46 and 47 in P. aeruginosa derives from the presence of acp3U, we aimed to map tRNA modifications in a mutant lacking the enzymatic activity required for this modification. In E. coli, acp3U is deposited by TapT, an enzyme containing a DTW domain (Takakura et al. 2019; Meyer et al. 2020). The P. aeruginosa protein with the highest level of sequence homology with E. coli TapT is PA14_17650, the ortholog of PAO-1 YfiP (Supplemental Table S12; Takakura et al. 2019). We obtained a mutant for PA14_17650 from a nonredundant library of P. aeruginosa strain PA14 transposon insertion mutants (Liberati et al. 2006). LC–MS/MS analysis of tRNA isolated from a PA14_17650 mutant strain confirms a role in acp3U deposition for this enzyme (Fig. 5A). Before generating sequencing libraries for the PA14_17650 mutant strain and a matched wild-type control, we tested multiple RT enzymes for sensitivity to modifications at this position. TGIRT generated the strongest signature at a site predicted to be acp3U modified (Supplemental Fig. S9). As observed in previous experiments, the correlation between mutational signatures across replicates was high for the tRNA sequencing libraries generated from the PA14_17650 mutant strain and the matched wild-type (Supplemental Fig. S10). Comparison between wild-type P. aeruginosa and the PA14_17650 mutant strain reveals a loss of misincorporation and termination signal specifically at positions 46 and 47 (Supplemental Figs. S11 and S12AB; Supplemental Table S13). This observation supports the idea that the m7G does not contribute to this mutational signature and establishes PA14_17650 as the P. aeruginosa homolog of E. coli TapT. The mutational signature on most tRNAs predicted to be acp3U modified is lost in the absence of PA14_17650, confirming these tRNAs carry either the acp3U modification or a derivate, in P. aeruginosa. The two exceptions are tRNAfMet1(CAU) and tRNAfMet2(CAU), suggesting that modifications at these positions are deposited by a different enzyme. Interestingly, the mutational signature observed at position 46 of tRNAGln1(UUG) was also lost in the PA14_17650 mutant, showing that modification at this position requires PA14_17650 activity. This position was previously found to be acacp3U modified in V. cholerae, where the primary tRNA sequence is these positions is UG, the same as P. aeruginosa (Kimura et al. 2020). However, we were not able to identify a clear homolog of AcpA from V. cholerae in P. aeruginosa by sequence alignment (Supplemental Table S12). In V. cholerae, this modification is also present at position 20 of tRNAGlu1(UUC). In P. aeruginosa we also observe a mutational signature at this position, but this signature is not dependent on PA14_17650 activity. This position could be acp3U modified by a different enzyme or have an unrelated modification. Further work is necessary to determine what is the nature of the modification present at position 20 of tRNAGlu1(UUC), position 47 of tRNAfMet1(CAU) and tRNAfMet2(CAU), and whether acp3U that is found at position 46 in tRNAGln1(UUG) can be subsequently modified to acacp3U.
DISCUSSION
RNA modifications expand the functional repertoire of each nucleoside, having a profound impact on RNA function. On tRNAs, RNA modifications play important roles in stabilizing secondary and tertiary structure and ensuring fidelity during translation (for review, see de Crécy-Lagard and Jaroch 2021). Across prokaryotic pathogens, tRNA modifications play critical roles in regulating stress responses, adaptation to the host environment, and expression of virulence factors (Koh and Sarin 2018). The repertoire of tRNA modifications in P. aeruginosa, an opportunistic pathogen, has been characterized using mass spectrometry (Jaroensuk et al. 2016; Grobe et al. 2019). However, a comprehensive understanding of the specific tRNA modifications present in each tRNA molecule is currently lacking. Determining which modifications are present on different tRNAs can help elucidate the role of enzymes involved in tRNA modification on bacterial survival and pathogenicity (Duechler et al. 2016). Here, we leveraged the well-characterized E. coli tRNA modification landscape to predict tRNA modifications in P. aeruginosa. Furthermore, we identified PA14_17650 as the gene responsible for acp3U deposition in the variable loop of P. aeruginosa tRNAs.
A cDNA mutational signature reference map for tRNA modifications in E. coli
In this study, we combined chemical capture targeting the s4U modification found close to the 5′ end of tRNAs, and cDNA misincorporation analysis to build a cDNA mutational signature map for tRNA modifications in E. coli with the objective of identifying modifications that are conserved in P. aeruginosa tRNAs. Although analysis of RT misincorporation involves less sample manipulation and provides information on a wide range of modifications, the chemical capture approach provides higher sensitivity for modifications of interest, overcoming limitations such as low read coverage at the modification site, lack of a mutational signature for a given modification, or low mutational rate. We find that the two approaches can be used as orthogonal validation for modifications of interest. Other approaches such as mass spectrometry or the use of mutant strains lacking the enzymatic activity for the modification in question are critical for evaluating and validating the activity of potential homologs, as we demonstrate in this study for acp3U. In E. coli, mutational signatures occur predominantly at the anticodon loop, and most positions with a mutational signature overlap with an annotated tRNA modification. We also detected mutational signatures in regions where a modification has not yet been annotated and might infer novel sites of modification worth further exploration. Once we generated a reference landscape of tRNA modifications in E. coli, we analyzed our P. aeruginosa tRNA sequencing data sets generated in parallel, to determine which modifications are conserved between the two species. Our data support the conservation of several modifications between E. coli and P. aeruginosa. A limitation of this approach is that several modifications do not generate a clear mutational signature. Some examples include modifications such as P. aeruginosa-specific 2′-O-methyladenosine (Am) modification at position 32 of tRNAPro1(GGG) (Jaroensuk et al. 2016), pseudouridine, 5-methyluridine, or dihydrouridine. For these modifications, this approach is not able to determine conservation between the two organisms. Chemical treatments targeting properties of specific modifications to induce a mutational signature can be leveraged to overcome this limitation (Wang et al. 2021).
Presence of s4U in P. aeruginosa tRNAS
We detected the presence of the s4U modification at position 8 of multiple tRNAs in both E. coli and P. aeruginosa using two orthogonal methods. This modification is conserved at most sites, with some exceptions; tRNAThr1(UGU) and tRNAAsn1(GUU) were not observed to have s4U in either method (Fig. 2; Supplemental Table S8). The occurrence of this modification was only observed in P. aeruginosa for a number of tRNAs, albeit at low levels, including tRNAPhe1(GAA), tRNAGly1(UCC), tRNAArg1(ACG), and tRNAPro1(CGG) (Fig. 2; Supplemental Table S7).
Anticodon loop modifications in P. aeruginosa
We identified several modifications in the anticodon loop that appear to be conserved between E. coli and P. aeruginosa (Fig. 3). Some of these modifications are synthesized by enzymes linked to virulence phenotypes, whereas others are deposited by enzymes required for survival. One example is ms2i6A at position 37. Synthesis of ms2i6A is a multistep reaction that requires the activity of MiaA, which converts A to isopentenyl adenosine (i6A), and MiaB, which converts i6A to ms2i6A. Although previous mass spectrometry studies did not detect this modification in P. aeruginosa (Jaroensuk et al. 2016; Grobe et al. 2019), Lin et al. (2022) demonstrated that loss of MiaB in the PAO1 strain leads to accumulation of i6A and a slight decrease in ms2i6A, suggesting the presence of this modification and potential alternative pathways for modifying i6A in P. aeruginosa. Alternately, ms2i6A might be further modified in P. aeruginosa. A protein with 39.92% sequence identity to Salmonella typhimurium MiaE, which converts ms2i6A to ms2io6A (Persson and Björk 1993), can be found in the P. aeruginosa genome (Supplemental Table S12; Jaroensuk et al. 2016) and might play a role in this pathway. Additionally, we observed strong conservation of the modified inosine at position 34 of tRNAArgQ(ACG), which is synthesized by TadA, coded by an essential gene in E. coli (Wolf et al. 2002). Other modifications known to play important roles in translation fidelity and protein homeostasis, such as ct6A, a modification universally found at position 37 of ANN-decoding tRNAs deposited in two steps involving multiple enzymes (Su et al. 2022), and m6t6A, deposited by TrmO (Kimura et al. 2014), are also conserved. The absence of the methyl group of m6t6A37 in tRNAThrV(GGU) reduces the efficiency of this tRNA to read its cognate codon (Qian et al. 1998). Formation of methyluridine derivates at the wobble position involves the enzymatic activity of MnmE and GidA (MnmG), and in some tRNAs, the position is further modified by MnmC. The 5-methylaminomethyl-2-thiouridine (mnm5s2U) modification, found in E. coli tRNAs tRNALys(UUU) and tRNAGltT/Glu1(UUC) is conserved in P. aeruginosa, and requires the activity of all three enzymes. Synthesis of mnm5s2U responds to growth conditions (Moukadiri et al. 2014), and cells lacking MnmC are not able to compete with a wild-type strain in growth assays (Moukadiri et al. 2014). Two particular modifications, m1G and m2A occur at position 37 and result in strong termination signatures. m1G is deposited by TrmD in E. coli, an enzyme that is essential in most bacterial species as it ensures accurate translation and prevents premature termination (Björk et al. 1989, 2001; Hagervall et al. 1993; Li and Björk 1995; Jaroensuk et al. 2019). Reduced levels of m1G impair membrane structure and sensitize cells to antibiotics (Masuda et al. 2019), suggesting a possible role for m1G in antibiotic resistance. m2A is deposited by RlmN (Benítez-Páez et al. 2012; Fitzsimmons and Fujimori 2016). The majority of m1G and m2A sites had conserved signatures between P. aeruginosa and E. coli. However, in the anticodon loop of tRNAHis1(GUG) and tRNAArg1(CCG), although we observed a mutational signature at position 37 in both species, the nucleotide identity at this position is different, indicating that different modifications are present (Fig. 3; Supplemental Table S7). In both cases, a strong termination is observed.
Variable loop modifications in P. aeruginosa
In the variable loop, there are two known modifications in E. coli, m7G deposited at position 46 by YggH and acp3U deposited at position 47 by TapT (Takakura et al. 2019; Meyer et al. 2020). Although there is no phenotype associated with the loss of YggH in E. coli, the deletion of TrmB, the enzyme responsible for m7G in P. aeruginosa, induces increased sensitivity to H2O2 (Thongdee et al. 2019). Additionally, levels of acp3U are influenced by the presence of m7G and culture conditions (Meyer et al. 2020). Although one study found loss of TapT to impact the thermostability of tRNAs, cell motility, and genome stability (Takakura et al. 2019), a different study found no impact of loss of TapT on growth under different culture conditions (Meyer et al. 2020). The lack of mutational signature in tRNAs annotated to be 5′-m7G U-3′ and the observation that RT stops are absent at position 47 in PA14_17650 mutant cells suggests that acp3U is the main contributor for the mutational signature observed at positions 46 and 47 (Meyer et al. 2020). We see conservation of the mutational signature in four tRNAs, with two other sites not showing evidence of being modified in P. aeruginosa, despite conservation of the GU sequence at positions 46 and 47. Signatures unique to P. aeruginosa are observed in tRNAArg1(CCG), tRNAfMet1(CAU), tRNAAsn1(GUU), and tRNAArg1(UCU) (Fig. 4; Supplemental Table S7). For tRNAArg1(UCU) the U required for acp3U is absent in E. coli tRNAArgU(UCU) but present in P. aeruginosa (Supplemental Table S7). Additionally, we observe a mutational signature in tRNAGln1(UUG) in a sequence context that is different from E. coli. In V. cholerae, position 46 of the homologous tRNA is acacp3U modified by the enzyme AcpA (Kimura et al. 2020). The same enzyme also modifies U20B in tRNAGlu1(UUC), a position where we also observe a mutational signature in P. aeruginosa that is absent in the homologous E. coli tRNAGltT(UUC) (Fig. 1). Although the similarities between P. aeruginosa and V. cholerae suggest acacp3U is also present in P. aeruginosa, we were not able to identify a clear homolog for AcpA, although a few enzymes with the same acetyltransferase domain are present the P. aeruginosa genome. Using the PA14_17650 mutant strain, we validated the presence of acp3U on several tRNA in P. aeruginosa (Supplemental Table S12; Fig. 5A,B), including tRNAGln1(UUG). However, we did not observe the loss of signature in the PA14_17650 mutant for tRNAGlu1(UUC), tRNAfMet1(CAU), and tRNAfMet2(CAU). There are at least two possible explanations for this observation. Either these positions are modified by a different enzyme with a DTW domain (Takakura et al. 2019) or an unrelated modification is present at this position.
Here we leveraged the wealth of information on E. coli tRNAs to explore the landscape of tRNA modifications on P. aeruginosa. With this approach, we were able to infer which tRNA modifications are conserved in the two species. Our data support a high degree of conservation between the two organisms and support the presence of tRNA modifications in P. aeruginosa that are not present in E. coli. Determining the specific modifications on each tRNA will provide insights into which pathways are impacted by different tRNA-modifying enzymes. Altogether, the approach described here can be used to study tRNA modifications in other unexplored species or across different culture conditions. Identifying RNA modifications and enzymatic pathways can help reveal targets to potentially reduce pathogenicity, virulence, and antibiotic resistance.
MATERIALS AND METHODS
Strains and growth conditions
Strains used in this work are as follows: E. coli wild-type (BW25113) and E. coli ΔthiI (JW0413-1, KEIO collection); P. aeruginosa (PA14), P. aeruginosa PA14_17650 transposon insertion strain (Liberati et al. 2006). Bacterial strains were grown in liquid media at 37°C and shaken at 235 rpm. For all experiments, bacterial strains were inoculated in LB overnight. Cultures were diluted 1:100 or in the indicated media, grown in triplicate, and collected at (OD)600 = 0.6 (log phase). CDM was prepared as described in Khare and Tavazoie (2015). For the E. coli cultures grown in CDM, cultures were diluted 1:250. P. aeruginosa was grown in CDM. Media used to culture cells in this study: LB (KD Medical; BLF-7030); CDM (1× M63 salts + 0.4 µM ferric citrate + biotin [0.1 µg/mL] + nicotinic acid [2 µg/mL] + 1× EZ supplement + 1× ACGU + 1 mM MgSO4 + Glucose [0.3%]).
RNA isolation and RNA library preparation
RNA was isolated with TRIzol reagent (ThermoFisher 15596018) (1.2 mL per 25 mL culture after pelleting cells). Samples were extracted with 240 µL of chloroform (Fisher C298-500). Precipitation of the aqueous layer was accompanied with 1 µL of 0.1 M DTT (18091050) and 15 µg of GlycoBlue reagent (ThermoFisher AM9516). Purified RNA was subject to a second round of isolation using QIAGEN RNA columns (74106) with 50 µL RNA + 3.5 volumes of RLT buffer + 10.5 volumes of 100% ethanol. RNA was washed twice, following the manufacturer's protocol, and eluted 2× in 50 µL of RNase-free water. Twenty-five micrograms of RNA was incubated with 7.5 µg of biotin-conjugated MTSEA (1 mg/mL) (90066-1) in MTSEA buffer (20 mM HEPES pH 7.3 [A611-J848-10], 10 mM EDTA [AM9261]) at 37°C for 2.5 h protected from light (600 rpm shake). After labeling with MTSEA, RNA was cleaned again on QIAGEN RNA columns. Labeling RNA with MTSEA results in a reversible covalent link by nucleophilic disulfide exchange between sulfur on s4U and MTS. The now biotinylated s4U-containing RNAs were then captured using C1 streptavidin beads (65001). Twenty-five microliters of C1 beads per sample were prepared by washing twice with 500 mL of water, followed by two washes in high salt buffer (100 mM Tris pH 7.4 [RGF-3340], 10 mM EDTA [AM9261], 1 M NaCl [AM9759], 0.05% Tween-20 [85113]). Eleven microliters of high salt buffer was added to each RNA sample and combined with beads resuspended in 11 µL of high salt buffer. RNA and beads were tumbled in the dark for 30 min at room temperature. After capture, beads containing s4U-RNA were washed three times in high salt buffer, followed by three washes in NT2 buffer (50 mM Tris pH 7.4 [RGF-3340], 1 mM MgCl2 [AM9530G], 150 mM NaCl [AM9759], 0.0005% NP-40 [11332473001]). Following washes, s4U-containing RNAs were eluted by tumbling for 15 min in the dark at room temperature in 100 mL elution buffer (100 mM DTT [18091050], 20 mM HEPES pH 7.3 [A611-J848-10], 1 mM EDTA [AM9261], 100 mM NaCl [AM9759], 0.05% Tween-20 [85113]). Elution was repeated one more time, and the volumes from both extractions were combined. RNA was cleaned using the Zymo RNA Clean and Concentrator kit (R1016), following the manufacturer's protocol and eluted twice in 7.5 µL of H2O.
Sequencing libraries were prepared from total RNA and captured s4U-containing RNA. 3′ end adaptors (Supplemental Table S14) were ligated to 100 ng of RNA in an overnight reaction at 16°C (250 nM adaptor, 1× T4 RNA ligase buffer, 1 µL of T4 RNA Ligase 1 [ssRNA Ligase], High Concentration [M0437M], 50 mM DTT [Sigma 43816-10mL], 6 µL of 50% PEG 8000 [NEB B1004A], 40 units of Ribolock RNase inhibitor [EO0382]). All samples were then gel purified on a denaturing 10% polyacrylamide gel (National Diagnostics, EC-829). Gels were cut between 60 and 200 nt and purified via crush and soak overnight tumble at 4°C (crush gel through 18 gauge needle punched bottom of 500 mL tubes into 1.7 mL Eppendorf tubes, high-speed spin for 10 min at RT, and soak crushed gel in 10 mM Tris pH 7.5 [15567-027], 500 mM NaCl [AM9759], 1 mM EDTA [AM9261], 0.1% SDS [351-032-101]). Gel buffer mix was then transferred to SpinX columns (CLS8162-96EA) to remove the remaining gel. Three volumes of 100% ethanol were added to the RNA-containing buffer for overnight precipitation. The purified RNA was then reverse transcribed using TGIRT and uniquely barcoded RT oligonucleotides per sample. Oligonucleotide design and library preparation were adapted from a method previously described (Supplemental Table S14; Zarnegar et al. 2016). RT included the RNA plus 1 µL of TGIRT enzyme (TGIRT50 from InGex), 1× buffer (20 mM Tris HCl pH 7.5 [15567-027], 5 mM MgCl2 [AM9530G], 450 mM NaCl [AM9759]), 1 µL 100 mM DTT (18091050), and 1 µL of 3 mM RT oligonucleotide, in a 19 µL reaction. This mix was incubated for 30 min at room temperature, followed by the addition of 1 µL of 25 mM dNTPs (N0446S), and incubation for 1 h at 60°C. cDNAs were then released from the RT TGIRT complex by treating with 1 µL of 5M NaOH, incubating for 3 min at 95°C, and followed by neutralization of cooled sample with 1 µL of 5M HCl. cDNAs were then pooled in groups, such as inputs or captured samples, and cleaned using the Zymo DNA Clean and Concentrator kit (D4014). Individually barcoded and cleaned pools of samples were then gel purified to remove excess oligonucleotide, following the same protocol as above, except that eluted cDNA was cleaned through the Zymo DNA Clean and Concentrator kit (D4014). Eluted pools were then subjected to circularization using a CircLigase-II kit (CL9021K) (cDNA, 0.75 µL MnCl2, 0.75 µL CircLigase-II, 1.5 µL of 10× buffer in a 15 µL reaction). After circularization, cDNA was cleaned, mixing with 30 µL of Ampure XP beads (A63881) and 75 µL of isopropanol, incubated 10 min at room temperature, applied to magnet, washed two times 30 sec each with freshly made 80% ethanol. Beads with cDNA were air dried for 5 min and cDNA was then eluted from beads in 14 µL of H2O. Directly following cleanup, cDNA was amplified with a first set of oligos in a PCR reaction on a qPCR machine to detect amplification in real time to capture linear amplification (0.75 µL of 20 µM P3 tall and P6 tall oligo mix, 15 µL of 2× NEBNext [M0541L], 14 µL cDNA, and 0.25 µL 25× diluted Sybr green [S7563] in a 30 µL reaction). PCR parameters were 40 sec at 98°C, cycling for 20 sec at 98°C, 30 sec at 65°C , 45 sec at 72°C , and a final extension for 5 min at 72°C. The number of cycles was determined by real-time amplification. Samples were then cleaned again using Ampure XP beads (A63881) (54 µL of beads, 4 µL of isopropanol, and eluted in 10 µL of H2O). Eluted cDNA was amplified for three more cycles using a second set of oligonucleotides for Illumina sequencing in an NEBNext Master Mix reaction. Following the second PCR, samples were cleaned one last time on the Ampure XP beads (36 µL of beads, 2.7 µL of isopropanol, and eluted in 12 µL of H2O). Final libraries were analyzed by DNA high-sensitivity tape station.
Targeted RT sequencing
Adapted from a previously described method, we performed targeted RT sequencing (Link et al. 2022). Briefly, RNA was isolated from TRIzol Reagent (ThermoFisher 15596018) with chloroform according to the manufacturer's protocol, except that RNA was isolated from the aqueous layer using QIAGEN columns, washed 2× in 500 µL of RPE wash buffer, dried, and eluted with 100 µL of H2O. Reverse transcription was performed with 500 ng RNA each using TGIRT (obtained from the Lambowitz laboratory, University of Texas at Austin) (Mohr et al. 2013), SuperScript III (ThermoFisher 18080044), SuperScript IV (ThermoFisher 18090050), or Induro (NEB M0681L) RTs. Each RT was performed using an RT oligo that was specifically designed to target the 3′ end of tRNAArg1(ACG) (Supplemental Table S14). TGIRT RT was performed just as described in the library preparation, SuperScript III RT was performed, following the manufacturer's protocol (500 ng RNA, 1 µL of 3 µM RT oligo, 4 µL of 5× FS buffer and up to 16 µL total with H2O). Samples were annealed for 5 min at 65°C, for 1 min on ice, and then supplemented with 1 µL 10 mM dNTPs, 1 µL of 0.1 M DTT, 1 µL RNasin (Promega N2611), and 1 µL of SuperScript III enzyme, and then incubated for 1 h at 55°C , followed 15 min at by 70°C. Similarly, SuperScript IV RT was performed, following the manufacturer's protocol. First step (500 ng RNA, 1 µL of 3 µM oligo, 1 µL of 10 mM dNTP mix, and up to 13 µL of H2O) incubated for 5 min at 65°C, 1 min on ice, followed by addition of 4 µL of 5× SSIV buffer, 1 µL of 0.1M DTT, 1 µL of RNase inhibitor (ThermoFisher EO0381), and 1 µL of SSIV enzyme, and incubation for 10 min at 55°C, followed by heat inactivation for 10 min at 80°C. Finally, Induro RT was also performed according to the manufacturer's protocol, first incubating for 5 min at 65°C, and then 1 min on ice (500 ng RNA, 1 µL of 3 µM oligo, 1 µL of 10 mM dNTP mix up to 10 µL of H2O), followed by the addition of 4 µL of 5× Induro RT reaction buffer, 0.2 µL of RNase inhibitor, 1 µL of Induro enzyme, and 4.8 µL of H2O, incubating for 1 h at 60°C, and heat inactivated for 1 min at 95°C. Following all RT reactions, samples were PCR amplified using HF Phusion enzyme (NEB M0530L) (2 µL of cDNA, 10 µL of 5× HF buffer, 2.5 µL of 10 µM forward oligo, 2.5 µL of 10 µM reverse RT oligo, 1 µL of 10 mM dNTP, 0.5 µL of Phusion, and 31.5 µL of H2O). PCR parameters were 1 min at 95°C, cycling for 15 sec at 95°C, 15 sec at 61°C, 30 sec at 72°C , for 33 cycles, and a final extension for 7 min at 72°C. Following the PCR, samples were cleaned with the Zymo DNA Clean and Concentrator kit (D4014), following the manufacturer's protocol and eluted 2× in 11 µL of H2O each. Samples were quantified and then submitted for Sanger sequencing using the forward sequencing oligo (oligos listed in Supplemental Table S14).
tRNA purifications and mass spectrometry
tRNAs were gel purified from total RNA on 10% UREA-PAGE. Briefly, 10–20 µg of RNA-containing 2× RNA loading dye (ThermoFisher R0641) were heated for 1 min at 95°C before loading onto a prerun 10% UREA-PAGE gel, in comparison to low molecular mass ssRNA (NEB N0364S). Gels were cut on a blue light box corresponding to the size of tRNAs (70–100 nt) and purified via crush and soak overnight tumble at 4°C (crush gel through 18 gauge needle punched the bottom of 500 mL tubes into 1.7 mL Eppendorf tubes, high-speed spin for 10 min at room temperature, and soak crushed gel in 10 mM Tris pH 7.5 [15567-027], 500 mM NaCl [AM9759], 1 mM EDTA [AM9261], 0.1% SDS [351-032-101]). Gel buffer mix was then transferred to SpinX columns (Sigma CLS8162-96EA) to remove the remaining gel, and 1 µL of GlycoBlue coprecipitant (ThermoFisher AM9516). Three volumes of 100% ethanol were added to the RNA-containing buffer for overnight precipitation. Ethanol precipitations were then centrifuged at high speed for 45 min at 4°C, washed 1× with 75% ethanol, air dried for 5 min, and resuspended in 50 µL of H2O. Purified tRNAs were then recleaned using the Zymo RNA Clean and Concentrator kit (R1016) according to the manufacturer's protocol, except we used three volumes of 100% ethanol in the first step. Samples were eluted 2× 11 µL of H2O and quantified by DeNovix spectrophotometer (DS-11+) before preparing for mass spectrometry.
Total tRNA or decapped RNA was subsequently digested to nucleosides in a 30 µL reaction containing 24 µL RNA, 1 µL of 2 U/µL Nuclease P1 (Sigma-Aldrich N8630), 3 µL 1 M ammonium acetate, pH 5.2, and 100 fmol of 13C-adenosine (Cambridge Isotope CLM-3678-0.05) as an internal standard. Reactions were digested for 3 h at 45°C. After 3 h, 3 µL of 1 M ammonium bicarbonate and 1 µL of 0.002 U/µL of snake venom phosphodiesterase from Crotalus adamanteus (Sigma-Aldrich P3243) were added to the reaction and incubation continued for 2 h at 37°C. After 2 h, 1 µL of 1 U/µL bacterial alkaline phosphatase (ThermoFisher 18011015) was added to the reaction and the reaction continued for 1 h at 37°C. Digested nucleosides were passed through a 3 kDa MWCO spin filter (Amicon UFC500396) and the filter was rinsed with 3 × 200 µL nuclease-free water before being dried under vacuum. Dried samples were dissolved in 45 µL buffer A (0.1% formic acid). Nucleosides were separated on a Luna Omega C18 reverse-phase column (1.6 µm PS, 100 Å, 30 mm, ID 2.1 mm, Phenomenex) with a flow rate of 0.200 µL/min under the following conditions: buffer A = 0.1% formic acid in high-performance liquid chromatography (HPLC)-grade water; buffer B = 0.1% formic acid in acetonitrile. The gradient for the liquid chromatography is as follows: gradient 0.2% buffer B from 0 to 2.6 min; gradient 2% buffer B from 2.6 to 5.0 min; gradient 8.5% B from 5.01 to 6.75 min; gradient 40% B from 6.76 to 7.5 min; 50% buffer B from 7.51 to 10 min; 0% buffer B from 10.01 to 14.0 min. Following chromatography, nucleosides were resolved on a SciEX 500 QTOF in positive electrospray ionization mode using a multiple-reaction-monitoring (MRM) MS method over the range of 100–500 Daltons (spray voltage = 5500 V; curtain gas = 35; CAD gas = 7; temp = 500°C; declustering potential = 25 V; collision energy = 10 V; accumulation time = 0.25 sec). The nucleosides were quantified using the nucleoside-to-base ion mass transitions. Chemical standards for the modified acp3U were obtained from Smolecule (S562863). Statistical significance was determined by Student's t-test.
Modifying enzyme homology search between P. aeruginosa and E. coli
Enzymes responsible for tRNA modifications in E. coli were identified from a search of the literature (Kimura et al. 2020; de Crécy-Lagard and Jaroch 2021). Accession numbers, genomic loci, and corresponding peptide sequences were retrieved from NCBI's gene lookup tool, specifically referencing E. coli K-12 MG1655 (NCBI:txid511145). Peptide sequences were used to carry out a BLASTP search using the Pseudomonas Genome Database's tool looking for proteins found in the P. aeruginosa strain PA14 (UCBPP-PA14 2006) (Winsor et al. 2016). If a match was found, the top hit was listed along with the total number of hits. Although our approach used protein sequence alignment, inferring direct evolutionary relationships can be challenging, even if sequences have diverged only moderately, because of the underlying models used (Rajapaksa et al. 2023). Therefore, functional validation may be required for some of the tRNA-modifying enzymes in P. aeruginosa predicted to be E. coli homologs. The predicted homologous proteins can be found listed in Supplemental Table S12.
RNA sequencing analysis
RNA sequencing was performed using NextSeq 550 or 2000 Illumina sequencers. Sequence reads were separated by barcode and duplicated reads were removed. Adapter sequences were trimmed. Reads were mapped with Bowtie2 to a custom transcriptome using a local alignment tool. Read counts were compiled for each sample and analyzed using DESeq2 to identify the enrichment of reads from s4U-captured RNAs as compared to inputs (Supplemental Tables S3 and S8). tRNAs were considered enriched if l2fc > 0.06, Padj <0.05, and >100 read coverage in the input sample.
To generate custom transcriptomes for both E. coli and P. aeruginosa, fasta files for annotated cDNA and ncRNA were downloaded from Ensembl for genome assembly ASM584v2 (GCA_000005845) and ASM1462v1 (GCA_000014625.1), respectively. For tRNA genes, duplicated sequences were removed, so that only unique sequences are present in the final transcriptome. CCA was added to the 3′ end on tRNAs in the P. aeruginosa transcriptome. Codon identity for each P. aeruginosa tRNA gene was obtained from the tRNA database (Jühling et al. 2009). Annotated modification information was obtained from Kimura et al. (2020) and Boccaletto et al. (2022). For all misincorporation and early termination analyses, only positions with 100 reads or higher were considered. E. coli and P. aeruginosa tRNA were matched based on anticodon and by sequence similarity, determined with the Emboss Needle tool (https://www.ebi.ac.uk/Tools/psa/emboss_needle/).
Dot blot and acrylamide gel analysis
Five milligrams of RNA was incubated with 2.5 mg biotin-conjugated MTSEA (1 mg/mL) (Biotium 90066-1) in MTSEA buffer (20 mM HEPES pH 7.3 [Quality Biological A611-J848-10], 10 mM EDTA [Invitrogen AM9261] for 2.5 h at 37°C [600 rpm shake]). Samples were extracted with 100 µL of chloroform (Fisher C298-500) and the aqueous phase was processed on a Zymo column (Zymo R1018). Three-hundred nanograms of each sample was loaded on a Hybond-N+ membrane (Amersham RPN119B). The membrane was cross-linked with UV (254 nm) for 120,000 mJ, twice. Membrane was blocked in Odyssey blocking buffer (LiCor 927-40003) for 10 min. IRDye 800CW Streptavidin (LiCor P/N 926-32230) was added to the blocking buffer at 1:1000 dilution and incubated for 45 min. Membrane was washed with phosphate buffer saline (PBS) (pH 7.2) (KD Medical) Tween 20 (0.1%) (Sigma P9416) two times and once with PBS (KD Medical) and imaged on ODYSSEY CLx (LiCor). A solution of 0.04% Methylene blue (Sigma M4159) in 0.5 M sodium acetate (NaOAc) (Invitrogen AM9740) was used to stain nucleic acids for loading control. The membrane was washed with DI water and imaged on a ChemiDoc MP Imaging system (BioRad).
Denaturing polyacrylamide gels were made using the SequaGel UreaGel 29:1 Denaturing Gel System (EC-829). Nondenaturing gels were made with acrylamide (30% Acrylamide/Bis Solution, 29:1 [1610156]), 1× TBE (BP13334), 0.1% ammonium persulfate, and 0.1% TEMED (1610800). Gels were run with size standards for RNA or DNA (low range ssRNA ladder [N0364S]; low molecular mass DNA ladder [N3233L]). Gels were visualized by staining with Sybr Gold nucleic acid gel stain (S11494).
DATA DEPOSITION
The raw sequencing data reported in this paper have been deposited in the Gene Expression Omnibus (GEO) at GSE236676. The code and scripts used to analyze the data and generate the figures are available at https://github.com/BatistaLab/Prokaryotic_tRNA.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
We thank Susan Gottesman, Richard Maraia, Efsun Arda, and members of Batista laboratory for critical reading of the manuscript, as well as Anupama Khare for providing the bacteria strain, reagents, and protocols. We also thank Alan Lambowitz at University of Texas at Austin, for providing us with the TGIRT enzyme. The authors thank the Center for Cancer Research (CCR) Genomics Core in Bethesda, Maryland for help with high-throughput sequencing. This work used the computational resources of the National Insitutes of Health (NIH) HPC Biowulf cluster (http://hpc.nih.gov). We appreciate the editorial assistance of George Leiman. Research was supported by the Intramural Research Program at the National Cancer Institute (NCI) of the NIH. The views expressed in this article are those of the authors and may not reflect the official policy or position of the NIH. Mention of trade names, commercial products, or organizations does not imply endorsement by the U.S. Government.
Footnotes
-
Handling editor: Eric Phizicky
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080004.124.
- Received February 26, 2024.
- Accepted April 9, 2024.
This is a work of the US Government
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Mariana D. Mandler is the first author of this paper, “The modification landscape of Pseudomonas aeruginosa tRNAs.” Mariana is a biologist in the Batista laboratory at the National Cancer Institute (NCI).
What are the major results described in your paper and how do they impact this branch of the field?
In this paper, we use orthogonal techniques, analysis of mutational signature during cDNA synthesis and chemical capture, to identify RNA modifications in tRNA of an understudied pathogenic bacteria, Pseudomonas aeruginosa. Although there are limitations to the techniques we use, wherein certain types of RNA modifications do not create mutational signatures, we were able to predict tRNA modification status in P. aeruginosa by comparison to E. coli. Furthermore, we validated the presence of acp3U and identified the enzyme responsible for the deposition of this modification in the variable loop. Many of the modifications explored here play roles in the virulence and pathogenesis of bacteria.
What led you to study RNA or this aspect of RNA science?
RNA modifications play major roles in regulating RNA metabolism and thus cellular function. More specifically, tRNA are known to be covered in modifications: averaging eight per tRNA in prokaryotes and 13 per tRNA in eukaryotes. The initial motivation of this study was to determine variation in levels and types of modification in tRNA in response to environmental conditions. As performed here, the methods can be extended to measure variation between species or could also be used to explore variation in different conditions within the same species.
During the course of these experiments, were there any surprising results or particular difficulties that altered your thinking and subsequent focus?
The project initially began by studying the variation of a particular RNA modification, 4-thiouridine, in E. coli during the log versus stationary phase and in different media. One condition we were interested in was the impact of bacterial communities on tRNA modifications. With help from Dr. Khare, we started to use P. aeruginosa and spent media as a model. During this work, we realized that not much was known about the location of tRNA modifications in P. aeruginosa, and we decided to use both methods to explore tRNA modifications. In P. aeruginosa, we were interested to learn the similarities and the differences in RNA modifications as compared to E. coli and other pathogenic bacteria, such as Vibrio cholerae.
What are some of the landmark moments that provoked your interest in science or your development as a scientist?
I have always been a curious person. In high school and college, I was interested in science related to human behavior, psychology, and neuroscience. After college, my father was diagnosed with a brain tumor. Soon after, I decided to participate in the NIH postbaccalaureate program in a radiation oncology laboratory. This was my first real research biology laboratory experience outside of school. I absolutely fell in love and decided graduate school and research was the next step for me. It was during graduate school that I learned more about RNA. I was fascinated by how much RNA could do, that not only does RNA carry the message, it also contains the information necessary for making, transporting, and translating the message and can regulate which message needs to be made at any given time in a cell. I decided that focusing my studies on RNA could answer so many questions regarding biology. Since then, I have continued my research in the field of RNA and remain fascinated every day.
If you were able to give one piece of advice to your younger self, what would that be?
Do not ever compare yourself to others. Instead, take what you can to learn from the aspects that you like and see different in others. Focus on the question, why it is interesting and how can we answer it, rather than try to “make it” as a scientist. “Making it” sounds great, but those that do usually follow this approach themselves. Also, do not let the pressure of competition and career success deter you from loving the work that you do every day.
Are there specific individuals or groups who have influenced your philosophy or approach to science?
I have learned an enormous amount from each one of my advisors and mentors along my journey. I will say that the RNA community especially drew me in as I realized how fascinated each member of this community also seems to be regarding the immeasurable capabilities of RNA. Dr. Deborah Citrin at NCI was my post-baccalaureate mentor, and without having the opportunity to experience research and exploration as I did then, I would not be where I am today. Several of my professors during graduate school, including Dr. Anita Corbett, challenged and encouraged me through my studies. My graduate school advisor Dr. Yue Feng at Emory University taught me how to think critically about each question and how to determine the best methods to approach each question. I learned from my postdoc advisor Dr. Shalini Oberdoerffer at NCI how to focus on the game-changing questions and to never let a negative result deter you from moving forward, instead, allow it to encourage you to change your mindset and to follow the science. In my current position, working with Dr. Pedro Batista, I have room to focus on the details in the fabric to answer questions that are crucial to our understanding of biology. Each of my advisors along the way taught me bits and pieces of these same ideologies, and I appreciate all that I have learned from them and the space they have given me to explore.
What are your subsequent near- or long-term career plans?
I am currently a biologist in the Batista laboratory. I absolutely love the position I am in, as it fits well with my personality. Without having to worry about the next steps in my career, I am able to focus on my own research, while also helping out with other projects. I get to train fellows, make sure the laboratory runs smoothly, and continue tackling questions regarding that fascinating molecule RNA.








