
Novel RNA molecular bioengineering technology efficiently produces functional miRNA agents
- Department of Biochemistry and Molecular Medicine, University of California-Davis, School of Medicine, Sacramento, California 95817, USA
- Corresponding author: aimyu{at}ucdavis.edu
-
Handling editor: Javier Caceres
Abstract
Genome-derived microRNAs (miRNAs or miRs) govern posttranscriptional gene regulation and play important roles in various cellular processes and disease progression. While chemo-engineered miRNA mimics or biosimilars made in vitro are widely available and used, miRNA agents produced in vivo are emerging to closely recapitulate natural miRNA species for research. Our recent work has demonstrated the success of high-yield, in vivo production of recombinant miRNAs by using human tRNA (htRNA) fused precursor miRNA (pre-miR) carriers. In this study, we aim to compare the production of bioengineered RNA (BioRNA) molecules with glycyl versus leucyl htRNA fused hsa-pre-miR-34a carriers, namely, BioRNAGly and BioRNALeu, respectively, and perform the initial functional assessment. We designed, cloned, overexpressed, and purified a total of 48 new BioRNA/miRNAs, and overall expression levels, final yields, and purities were revealed to be comparable between BioRNAGly and BioRNALeu molecules. Meanwhile, the two versions of BioRNA/miRNAs showed similar activities to inhibit non-small cell lung cancer cell viability. Interestingly, functional analyses using model BioRNA/miR-7-5p demonstrated that BioRNAGly/miR-7-5p exhibited greater efficiency to regulate a known target gene expression (EGFR) than BioRNALeu/miR-7-5p, consistent with miR-7-5p levels released in cells. Moreover, BioRNAGly/miR-7-5p showed comparable or slightly greater activities to modulate MRP1 and VDAC1 expression, compared with miRCURY LNA miR-7-5p mimic. Computational modeling illustrated overall comparable 3D structures for exemplary BioRNA/miRNAs with noticeable differences in htRNA species and payload miRNAs. These findings support the utility of hybrid htRNA/hsa-pre-miR-34a as reliable carriers for RNA molecular bioengineering, and the resultant BioRNAs serve as functional biologic RNAs for research and development.
Keywords
INTRODUCTION
MicroRNAs (miRNAs or miRs) are a superfamily of functional noncoding RNAs (ncRNAs) that are present in almost all species (Ambros 2003, 2004). Upon entering the RNA-induced silencing complex (RISC) and binding with target transcripts, miRNAs exert an RNA interference (RNAi) mechanism to achieve posttranscriptional gene regulation to modulate many critical cellular processes. The potency and versatility of miRNA to target multiple transcripts provide an opportunity for a single molecule to modulate common pathways that may be dysregulated in diseases (Yu et al. 2020; Traber and Yu 2023). Beyond initial discovery, researchers have developed novel, adaptable RNAi-based tools and biotechnologies for basic genetic and biomedical research as well as RNAi-based therapeutics (Yu et al. 2020; Traber and Yu 2023). In fact, there are a number of RNAi-based therapies being approved by the US Food and Drug Administration and many others in clinical or preclinical development for the treatment of various human diseases, including lung cancer (Mollaei et al. 2019; Yu et al. 2020; Yu and Tu 2022; Traber and Yu 2023; Gogate et al. 2024).
Lung cancer remains the leading cause of cancer-related deaths in the United States and worldwide with 85% of cases classified as non-small cell lung cancer (NSCLC) (Thai et al. 2021; Siegel et al. 2023). The low survival rate of NSCLC patients warrants the development of more effective pharmacotherapies (Grodzka et al. 2023; Siegel et al. 2023). However, growing interest in RNAi-based interventions and lung cancer-associated miRNAs positions NSCLC as a prime candidate for miRNA replacement therapy (Xue et al. 2017; Petrek and Yu 2019; Grodzka et al. 2023). In particular, miRNA replacement therapy reintroduces tumor-suppressive miRNAs depleted in disease cells to harness endogenous RNAi machinery for the treatment of human disease, representing a new viable strategy (Bader et al. 2010; Mollaei et al. 2019; Traber and Yu 2023). However, most conventional miRNA interventions make use of in vitro synthesized miRNA mimics or biosimilars bearing extensive chemical modifications whose physiochemical and biological properties may not truly recapitulate their endogenous counterparts (Ho and Yu 2016; Yu et al. 2019,2020; Traber and Yu 2023). While chemical modifications may improve metabolic stability and binding affinity, such as the modifications on the ribose subunit of individual ribonucleotides as demonstrated in the approved RNAi medications and the latest locked nucleic acids (LNAs) under active development, the inclusion and introduction of nonnatural chemical modifications might increase the risk of severe immune responses (Yu et al. 2019, 2020; Traber and Yu 2023).
To address these concerns, our laboratory has developed a novel in vivo fermentation-based, RNA molecular bioengineering platform technology by using a human transfer RNA (htRNA) fused hsa-pre-miR-34a carrier, which offers true, biologic RNAi agents (termed BioRNAs, previously BERAs) (Fig. 1A) with high yield and purity that might more closely represent the structural and chemical properties of endogenous miRNAs (Chen et al. 2015; Ho et al. 2018; Li et al. 2021). Furthermore, our BioRNAs exhibit biocompatibility and efficacy to suppress tumor growth in vivo and regulate target gene expression in vitro (Chen et al. 2015, 2023; Ho et al. 2018; Jilek et al. 2019; Tu et al. 2019; Li et al. 2021; Petrek et al. 2021). Through our previous work, the hybrid htRNA/hsa-pre-miR-34a carriers are imperative for in vivo microbial fermentation production and efficient release of payload RNAi molecules to exert their biologic functions in mammalian cells (Chen et al. 2015; Ho et al. 2018; Tu et al. 2020; Yi et al. 2020; Li et al. 2021).
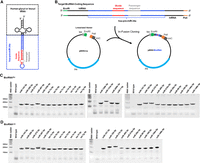
Heterologous overexpression of a total of 48 new BioRNAs with human glycyl or leucyl tRNA fused hsa-pre-miR-34a carrier (BioRNAGly or BioRNALeu). (A) The 2D graphic of BioRNAGly or BioRNALeu with a payload miRNA whose guide and passenger sequences are highlighted. (B) Sequence design of target BioRNA inserts for cloning into a pBSKrna vector linearized by EcoRI and PstI restriction enzymes to offer the BioRNA expression plasmid (pBSK/BioRNA), which is comprised of a lipoprotein (lpp) promoter, terminator from the Escherichia coli ribosomal RNA rrnC operon (rrnC), and the gene encoding β-lactamase (bla) for antibiotic resistance and selection. (C, D) Urea–PAGE analyses depicting the successful overexpression of 26 BioRNAGly (C) and 22 BioRNALeu (D) versions of recombinant miRNAs, as manifested by the appearance of a strong new band at the expected size when compared to total RNA from the wild-type HST08 E. coli; 300 ng RNA per lane. Urea–PAGE analyses depicting the successful overexpression of BioRNALeu/miR-126b-3p, -133a-3p, -200b-3p, and -7-5p have been reported recently (Li et al. 2021).
From an initial screening, leucyl htRNA/hsa-pre-miR-34a-based BioRNAs (BioRNALeu) consistently showed a high level of heterologous overexpression in bacteria and were selectively processed to target miRNAs in human cancer cells (Li et al. 2021). A more comprehensive screening suggested that glycyl htRNA/hsa-pre-miR-34a-based BioRNA (BioRNAGly) exhibited a low cytotoxicity while showing a higher yield than other htRNAs (MJ Tu, N Batra, and AM Yu, unpubl.). Therefore, in the present study, we aim to compare the utility of htRNAGly- versus htRNALeu-fused hsa-pre-miR-34a carriers in producing corresponding BioRNAGly and BioRNALeu molecules with the same panel of payload miRNAs and in regulating target gene expression with a model miRNA, miR-7-5p (BioRNAGly/miR-7-5p versus BioRNALeu/miR-7-5p). Our data demonstrated that all 48 new BioRNAGly/miRNAs and BioRNALeu/miRNAs were successfully produced, and overall yields, purities, and antiproliferative activities were comparable between BioRNAGly- and BioRNALeu-based miRNAs. Furthermore, we validated the release of payload miR-7-5p and its efficiency to regulate the expression of several well-known target genes in two human NSCLC cell lines. Through computational modeling, we found the overall 3D structures of BioRNA/miRNAs and their subcomponents to be comparable amid notable differences in htRNAs and payload miRNAs. These findings support the robustness and reliability of hybrid tRNA/pre-miRNA carriers for molecular bioengineering of functional RNAi molecules that represent a novel class of true, biological RNAs for research and development.
RESULTS
In vivo fermentation production of novel biologic RNA agents and purification to a high degree of homogeneity
To compare the utilities of htRNAGly- and htRNALeu-fused hsa-pre-miR-34a carriers (Fig. 1A), we first identified a panel of recombinant miRNAs (Table 1) for in vivo fermentation production. Our BioRNA panel consisted of 22 novel BioRNALeu/miRNAs and 26 BioRNAGly/miRNAs, the latter including four counterparts to previously published BioRNALeu/miRNAs, specifically BioRNALeu/miR126b-3p, /miR133a-3p, /miR-200b-3p, and /miR-7-5p (Li et al. 2021). BioRNAGly/miRNAs and BioRNALeu/miRNAs (Fig. 1B; Supplemental Fig. S1; Supplemental Tables S1 and S2) were cloned into pBSKrna vector to offer target pBSK/BioRNA plasmids subsequently verified by DNA sequencing. Individual plasmids were transformed into HST08 E. coli competent cells, and overexpression of target BioRNA/miRNAs was confirmed through urea–PAGE analyses of total RNAs isolated from E. coli (Fig. 1C,D). We confirmed the heterologous overexpression of all 26 BioRNAGly/miRNAs (Fig. 1C) and 22 BioRNALeu/miRNAs (Fig. 1D) through the appearance of strong new bands at expected size, offering a 100% success rate and each accounting for >40% of total RNA based on the density of RNA band (Fig. 1C,D).
Yields, purities, and endotoxin activity levels of individual BioRNA/miRNAs using human leucyl or glycyl tRNA fused hsa-pre-miR-34a carrier, and isolated by anion exchange FPLC methods
Each BioRNA was thus subjected to anion exchange fast protein liquid chromatography (FPLC) purification. Fractions of eluted BioRNA monitored by UV detection (Fig. 2A) were collected and verified through urea–PAGE analysis as a single RNA band (Fig. 2B). The fractions were pooled, desalted, and concentrated to generate BioRNA product whose homogeneity was semi-quantitatively and quantitatively determined by urea–PAGE analysis (Fig. 2D,E) and high-performance liquid chromatography (HPLC)-UV analysis (Fig. 2C), respectively. Their endotoxin levels (Table 1) were further measured with an LAL assay kit. All FPLC-purified BioRNAs were highly homogenous (>97%) and most exhibited low endotoxin activity (≤5 EU/µg RNA) (Table 1). The majority of pure BioRNA products accounted for more than 30% of the total RNA, yielding 5–10 mg of pure BioRNA from 250 mL bacterial culture. Overall, the production yields, purity, and endotoxin levels of recombinant miRNAs using BioRNAGly and BioRNALeu carriers were not statistically significant (Table 1), supporting their applications to molecular bioengineering of target RNAs.
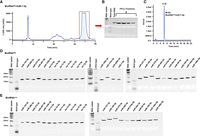
Purification of recombinant miRNA molecules. (A) Representative FPLC-UV trace during the purification of the model BioRNAGly/miR-7-5p with a highlighted peak for target fraction collection. (B) Urea–PAGE analysis of BioRNAGly/miR-7-5p fractions. (C) HPLC determination of the purity (99.6%) of final BioRNAGly/miR-7-5p product. (D, E) Urea–PAGE analyses depicting FPLC-purified BioRNAGly/miRNAs and BioRNALeu/miRNAs, respectively (50 ng RNA/lane). Purification of BioRNALeu/miR-7-5p, -126b-3p, -133a-3p, and -200b-3p has been published recently (Li et al. 2021).
Antiproliferative activities of novel BioRNAs against human NSCLC cells
To examine possible functional differences between BioRNAGly- and BioRNALeu-based miRNAs, we first used the CellTiter-Glo biochemical assay to determine their activities to inhibit NSCLC A549 and H1975 cell viability. Several recently published (Li et al. 2021) BioRNAs (Table 1) were included for comparison. A549 (Fig. 3A) and H1975 (Fig. 3B) cell viabilities were reduced to various degrees by 15 nM BioRNAGly/ and BioRNALeu/miRNAs. Overall, the antiproliferative effects were comparable between the paired BioRNAGly/miRNAs and BioRNALeu/miRNAs (e.g., miR-206-3p, miR-125b-5p, and miR-7-5p) and demonstrated putative cell line-specific differences. Similar results were obtained in pancreatic cancer AsPC-1 cells (Supplemental Fig. S2). Interestingly, A549 cells seemed more sensitive to a few BioRNALeu/miRNAs (e.g., miR-370-3p and -146a-5p) than corresponding BioRNAGly/miRNAs (Fig. 3A), while H1975 cells seemed slightly more sensitive to particular BioRNAGly/miRNAs (e.g., miR-495-3p and -146b-5p) than corresponding BioRNALeu/miRNAs with some exceptions (e.g., miR-133a-3p and -100-5p) (Fig. 3B). BioRNALeu/miR-22-3p exhibited a strong antiproliferation activity with putative cell line-specific differences, similar as those reported recently (Chen et al. 2023). Notably, miR-7-5p demonstrated comparable antiproliferative activities between BioRNAGly- and BioRNALeu-based carriers as well as against both NSCLC A549 and H1975 cells (Fig. 3A,B). These results support the activities of both BioRNAGly- and BioRNALeu-based miRNAs to inhibit NSCLC cell viability with putative dependence on cell lines and the embedded payload miRNAs.
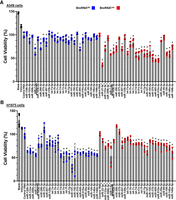
Screen for antiproliferative effects of 48 novel, highly purified BioRNAs against human NSCLC cells. Cell viability of A549 (A) and H1975 (B) cells was reduced to various degrees by BioRNAGly (blue) and BioRNALeu (red) based miRNAs (15 nM) as compared with control BioRNA or vehicle as well as untreated cells. Cell viability values were determined by CellTiter-Glo assay at 72 h post-transfection and normalized to respective control RNAs (100%). Values are mean ± SD (N = 4 biological replicates per group). #Several BioRNAs published recently were included for comparison. Overall, the effects on NSCLC cell viability were comparable between the paired BioRNAGly- and BioRNALeu-based miRNAs with putative cell line-specific differences. BioRNA/miR-7-5p used for further studies are underlined. (*) P < 0.05, as compared to respective control RNA (one-way ANOVA with Bonferroni post hoc tests).
BioRNAGly/miR-7-5p is selectively processed to target miR-7-5p in human NSCLC cells
We thus selected miR-7-5p as a model to assess the release of payload miRNA from BioRNA based on the comparable effectiveness of BioRNAGly/ and BioRNALeu/miR-7-5p to inhibit cell viability as well as the well-established anticancer function of miR-7-5p in NSCLC cells (Xiong et al. 2011; Li et al. 2019; Guo et al. 2020). We used the selective stem–loop RT-qPCR method to determine the levels of mature miR-7-5p in NSCLC cells following transfection with 15 nM of BioRNAGly/ and BioRNALeu/miR-7-5p and controls for 48 h. The data showed that BioRNA/miR-7-5p led to higher levels of miR-7-5p in NSCLC cells (Fig. 4A,B). Surprisingly, BioRNAGly/miR-7-5p resulted in a significant higher mature miR-7-5p level than BioRNALeu/miR-7-5p. In addition, BioRNAGly/miR-7-5p led to about 10-fold higher change in miR-7-5p levels in H1975 cells (Fig. 4A) than A549 cells (Fig. 4B). In contrast, the levels of miR-7-5p were similar between A549 and H1975 cells transfected with BioRNALeu/miR-7-5p or chemo-engineered mimic, and levels of miR-7-5p increased in BioRNALeu/miR-7-5p-transfected cells did not reach statistical significance when compared to each control. These results demonstrate the selective release of payload miRNA from BioRNA/miRNA in human NSCLC cells.
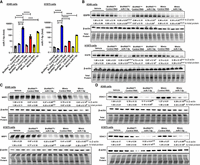
Efficacy of model recombinant miR-7-5p to regulate target gene expression in human NSCLC cells. (A) BioRNAGly- and BioRNALeu-carried miR-7-5p are processed to target miR-7-5p in A549 and H1975 cells. Cells were transfected with 15 nM BioRNA/miR-7-5p, control BioRNA, or Lipofectamine 3000 (vehicle) for 48 h, and miR-7-5p levels were determined by selective stem–loop RT qPCR assay. Commercial miRCURY LNA miR-7-5p mimic (Mimic miR-7-5p) and control RNA (Mimic Control) from Qiagen were used for comparison. (B) Impact of BioRNA/miR-7-5p on a well-known miR-7-5p target, epidermal growth factor receptor (EGFR) in A549 and H1975 cells after 72-h transfection, as determined by western blot analyses. Surprisingly, BioRNAGly/miR-7-5p more effectively reduced EGFR protein levels than BioRNALeu/miR-7-5p besides control RNA. Therefore, the effects of BioRNAGly/miR-7-5p versus the mimic on (C) multidrug resistance-associated protein 1 (MPR1) and (D) voltage-dependent anion channel protein 1 (VDAC1) protein levels were further defined. miR-7-5p and targeted protein levels were normalized to corresponding U6 and β-actin or total protein, respectively, and vehicle control groups were set as 1.0. All values are mean ± SD (N = 3 biological replicates per group). (****) P < 0.0001; (***) P < 0.001; aP < 0.05, as compared to respective control RNA; bP < 0.05, compared to vehicle control; cP < 0.05, compared to Mimic miR-7-5p; dP < 0.05, compared to BioRNALeu/miR-7-5p (one-way ANOVA with Bonferroni post hoc tests).
BioRNAGly/miR-7-5p effectively regulates target gene expression in human NSCLC cells
We next performed western blot analyses to investigate the effectiveness of BioRNA/miR-7-5p to regulate the expression of a well-known target, namely, EGFR, in NSCLC cells after a 72-h transfection. Our data showed that BioRNAGly/miR-7-5p suppressed EGFR protein levels by 76% and 68% in A549 and H1975 cells, respectively (Fig. 4B), while the same dose of BioRNALeu/miR-7-5p did not alter EGFR protein outcomes. Additionally, the extent of EGFR down-regulation by the same dose of BioRNAGly/miR-7-5p and commercial miRCURY LNA miR-7-5p mimic was comparable in both A549 and H1975 cells.
Based on these results, coupled with the findings on the increases in miR-7-5p levels, we elected to further evaluate the function of BioRNAGly/miR-7-5p to control the expression of two other targets, multidrug resistance protein 1 (MRP1) and voltage-dependent anion channel 1 (VDAC1). Western blot analyses revealed that BioRNAGly/miR-7-5p effectively reduced the protein levels of both MRP1 (73% and 75%) (Fig. 4C) and VDAC1 (82% and 72%) (Fig. 4D) in NSCLC cells (A549 and H1975, respectively). Additionally, the extents of MRP1 and VDAC1 down-regulation were comparable or slightly less in cells transfected with miR-7-5p mimics, as compared with BioRNAGly/miR-7-5p. These results confirm the effectiveness of BioRNA/miRNA to control target gene expression. Taken together, while both BioRNAGly/ and BioRNALeu/miR-7-5p were processed to mature miR-7-5p in NSCLC cells, much higher levels of miR-7-5p were noted in cells transfected with BioRNAGly/miR-7-5p than BioRNALeu/miR-7-5p, and thus BioRNAGly/miR-7-5p demonstrated a consistent efficacy in the control of target gene expression that was comparable as or slightly greater than the same dose of chemo-engineered LNA miR-7-5p mimics.
Computational modeling of BioRNA 3D structures
To explore the possible effects of htRNAGly and htRNALeu on hsa-pre-miR-34a and overt BioRNA structures, we used the RNAComposer to determine the 3D structures of several BioRNAs that were further visualized and analyzed with ChimeraX (Fig. 5). Computational modeling revealed that htRNAGly and htRNALeu maintained their canonical L-shaped 3D structures (Fig. 5A; Porat et al. 2021) when their anticodon sequences were replaced with hsa-pre-miR-34a (Fig. 5A). Similarly, the 3D structure of hsa-pre-miR-34a remained unchanged within the recombinant BioRNA (Fig. 5A).
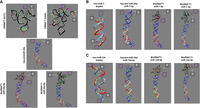
Comparison of computationally predicted 3D structures of BioRNAs. (A) Predicted 3D structures of human leucyl (UAA) and glycyl (GCC) tRNAs, hsa-pre-miR-34a, and the hybrid carrier RNAs (BioRNAGly/miR-34a-5p and BioRNALeu/miR-34a-5p). (B) Predicted structures of the hsa-miR-7 duplex, hsa-pre-miR-34a carrier with payload miR-7 (hsa-pre-miR-34a/miR-7-5p), BioRNAGly/miR-7-5p, and BioRNALeu/miR-7-5p. (C) 3D structures of the hsa-miR-124 duplex, hsa-pre-miR-34a carrier substituted by miR-124 duplex (hsa-pre-miR-34a/miR-124-3p), BioRNAGly/miR-124-3p, and BioRNALeu/miR-124-3p. Colored subcomponents of BioRNAs consist of ribbon structures: htRNA (black), hsa-pre-miR-34a (blue), miRNA guide (antisense; red) and passenger (sense; light blue or light gray) sequence, five-prime (5′; magenta), and three-prime (3′; white). Nucleotides consist of color-filled nucleobases: adenine (red), thymine (blue), guanine (green), and cytosine (yellow). Modeling results demonstrate a retained overall structure of BioRNAs when substituted with target miRNAs. All RNA 3D structures were generated by using RNAComposer after inputting the secondary structures predicted by RNAFold, based on the primary sequences shown in Supplemental Tables S1 and S4. The resulting structures were adapted using UCSF ChimeraX next-generation molecular visualization program.
As we were curious if the functional difference observed between BioRNAGly/ and BioRNALeu/miR-7-5p (Fig. 4A,B) were reflected by any structural elements, no meaningful steric differences were noted in the hsa-pre-miR-34a/miR-7-5p (Fig. 5B) besides changes in overall length of BioRNA and noticeable differences in respective htRNA subcomponents. Similar results were seen in paired BioRNAGly/, BioRNALeu/miR-124-3p, and their subcomponent derivatives (Fig. 5C). Overall, computational modeling indicated that structures of hsa-pre-miR-34a carrier and miRNA duplexes were unaltered in the paired BioRNAGly/miRNAs and BioRNALeu/miRNAs while noticeable differences depend on their respective htRNA species. In addition, exchanging the hsa-miR-34a duplex induced little change in chimeric hsa-pre-miR-34a/miR-7-5p and hsa-pre-miR-34a/miR-124-3p structures, as well as the whole BioRNAs.
DISCUSSION
The discovery and application of RNAi has ushered in a new era of adaptable biological and biomedical research with the development of novel routes for therapeutic intervention of human diseases (Bader et al. 2010; Xue et al. 2017; Mollaei et al. 2019; Traber and Yu 2023). With the understanding of loss or down-regulation of tumor suppressive miRNAs in lung and other types of cancers, novel tools may be developed to restore or reintroduce such miRNAs to achieve the control of disease progression, namely, miRNA replacement therapy. Nevertheless, RNAi research and therapy are dominated by using chemically or enzymatically synthesized RNA analogs (Ho and Yu 2016; Yu et al. 2020; Traber and Yu 2023). Alternatively, our present study, built upon our recent efforts in bioengineering recombinant RNA agents, demonstrates the reliability of both htRNAGly- and htRNALeu-fused hsa-pre-miR-34a carriers to achieve consistent, high-yield, in vivo fermentation production of recombinant RNAs, BioRNAGly/miRNAs, and BioRNALeu/miRNAs, respectively, which are effective to inhibit human NSCLC cell viability with both putative cell line- (Foster et al. 1998; Pao et al. 2005; Neuperger et al. 2022) and embedded payload miRNA-dependence (Petrek and Yu 2019).
Through functional analyses using model BioRNA/miR-7-5p, our results indicate that BioRNA/miRNAs are intracellularly processed to release the miRNA payload and effectively regulate the expression of well-known targets, namely, EGFR (Kefas et al. 2008), MRP1 (Liu et al. 2015b), and VDAC1 (Chaudhuri et al. 2016). As BioRNAGly/miR-7-5p exhibits a greater degree of efficacy to regulate EGFR expression than the same dose of BioRNALeu/miR-7-5p, further assessment demonstrates that BioRNAGly/miR-7-5p is equally or slightly more efficacious to regulate target gene expression when compared to the miR-7-5p mimic with popular LNA structures (Kaur et al. 2010; Lundin et al. 2013), associated with higher levels of mature miR-7-5p in BioRNAGly/miR-7-5p-transfected cells. These results suggest the possible impact of htRNA on the release of payload miRNA that may subsequently affect the efficacy to modulate target gene expression. Furthermore, the antiproliferative activities of individual BioRNA/miRNAs may be cell line-dependent in concert with the results from our unbiased viability screen. Therefore, it should be noted that possible differences in processing and target gene regulation between BioRNAGly and BioRNALeu may be sequence dependent and require further examination of additional targeted genes as well as paired BioRNA/miRNAs.
Although the limiting factor of BioRNALeu/miR-7-5p processing is not fully known, one may suspect that the structural or steric interactions induced by the htRNALeu sequence on the chimeric structure may affect identification and/or cytoplasmic processing by the microprocessor complex of the miRNA biogenesis pathway (Bergeron et al. 2010). The inclusion of the hsa-pre-miR-34a sequence is intended to guide BioRNA into the miRNA biogenesis pathway to interact with the microprocessor and release the mature and functional miRNAs, which may or may not be Dicer-dependent (Ho et al. 2018; Traber and Yu 2023). However, a lower extent of miRNA payload release from BioRNALeu/miR-7-5p than BioRNAGly/miR-7-5p, together with their antiproliferation and target gene regulation activities, suggest possible effects of the tRNA scaffold, which warrants further validation prior to extrapolating the observed effects to the full BioRNA/miRNA panel discussed in this study. Furthermore, although efforts were made to optimize the experimental conditions with multiple cell lines showing high transfection efficiency, caution should be noted that transfection efficiencies might vary among different BioRNA/miRNAs and mimics, as well as various cell lines, which, if any, would also contribute to the observed differences in particular effects.
While exploration of RNA structure is currently limited by the lack of available and confirmed RNA 3D structures (Bonilla and Kieft 2022; Liu et al. 2022; Ma et al. 2022), the integration of computational 3D predictive modeling allows for initial assessments of RNA structures. Through predictive modeling, our results demonstrate little change between both hsa-pre-miR-34a/miRNA chimeras and overall BioRNA structures with putative dependence on payload RNAs and respective htRNA species. Still, previous work has demonstrated both the effective processing and target gene regulation of BioRNALeu/miR-34a-5p and /miR-124-3p (Deng et al. 2021; Li et al. 2021) and BioRNAGly/miR-124-3p (MJ Tu, N Batra, GM Traber, et al., unpubl.), though to a lesser degree than BioRNAGly/miR-7-5p found in this study. Yet, all predicted 3D structures assessed in this study remain highly comparable to BioRNAGly/ and BioRNALeu/miR-7-5p. Together, these results suggest the sequence- and cell line-dependence in structural and functional analyses of our BioRNA/miRNAs which warrants a more comprehensive study to compare between heterologous BioRNA/miRNA panels. Nevertheless, provided that computational modeling remains only the initial work into determining the 3D structure of BioRNA/miRNA, it should be noted that experimental, structural studies may provide further insight into true differences in BioRNA/miRNA structure and functional efficiency.
In addition, to the best of our knowledge, these results demonstrate the first evidence of hsa-miR-7-5p to regulate both MRP1 and VDAC1 in human NSCLC cells, alongside EGFR (Webster et al. 2009). Regulation by hsa-miR-7-5p in NSCLC complements similar findings in previous reports of MRP1 regulation in breast cancer (Pogribny et al. 2010; Hong et al. 2019), hepatocellular carcinoma (HCC) (Hu et al. 2018), and small cell lung cancer (Liu et al. 2015a,b) cells. Similar regulation complements previous cancer study findings of VDAC1 regulation by hsa-miR-7-5p in cervical cancer (Zhang et al. 2019), HCC (Wang et al. 2016), neuroblastoma (Chaudhuri et al. 2016), and rhabdomyosarcoma (Yang et al. 2020) cells. The regulation of both MRP1 and VDAC1 in NSCLC alongside EGFR introduces new therapeutic routes by using biologic miR-7-5p to sensitize drug-resistant NSCLC cells, which await future exploration.
Conclusions
In conclusion, this study demonstrates the reliability of our novel RNA molecular bioengineering platform technology to allow in vivo fermentation production of target BioRNA/miRNA agents by using glycyl and leucyl htRNA/hsa-pre-miR-34a stable carriers. BioRNAs are produced with high yield, high degree of homogeneity, and 100% success rate to closely represent the properties of natural miRNA molecules. Overall antiproliferative activities between paired BioRNALeu/ and BioRNAGly/miRNA are comparable with putative cell line-specific differences. Functional studies with model BioRNA/miR-7-5p validate the intracellular release of miRNA payload and confirm the effectiveness of BioRNA/miRNA to regulate the expression of targeted genes in NSCLC cells. Further, computational modeling suggests a highly similar structure for the BioRNAs with noticeable differences in htRNA scaffolds. Lastly, our study also confirms the regulation of MRP1 and VDAC1 in NSCLC cells by hsa-miR-7-5p. Overall, these findings support the robustness of tRNA/pre-miRNA carrier-based RNA biotechnology and the applications of novel BioRNA molecules to basic research as well as the development of therapeutic RNAs for NSCLC.
MATERIALS AND METHODS
Chemicals and materials
Lipofectamine 3000 (Cat# L3000001), RPMI medium 1640 (Cat# 11875119), DMEM medium (Cat# 11965092), 0.05% trypsin-EDTA (Cat# 25300054), phosphate-buffered saline (PBS; Cat# 10010023), fetal bovine serum (Cat# 26140079), opti-MEM (Cat# 31985070), RIPA buffer (Cat# 89900), and bicinchoninic acid (BCA) Protein Assay Kit (Cat# 23227) were purchased from Thermo Fisher Scientific. Saturated phenol (pH 4.5; Cat# 97064-716) was purchased from VWR. Pure ethanol (Cat# E7023) and protease inhibitor cocktail (Cat# P8340) were purchased from Sigma-Aldrich. CellTiter-Glo 2.0 Cell Viability Assay Kit (Cat# G9241) was purchased from Promega. miRCURY LNA hsa-miR-7-5p mimic (Cat# 339174 YM00472714-AGA) and negative control (Cat# 339174 YM00479902-AGA) were purchased from Qiagen. Direct-zol RNA miniPrep Kit (Cat# R2061) was purchased from Zymo Research. All primers (Supplemental Table S3) were synthesized and purchased from Integrated DNA Technologies. iTaq Universal SYBR Green Supermix (Cat# 1725121), TGX Stain-Free FastCast Acrylamide Kit (7.5%; Cat# 1610181, and 10%; Cat# 1610183), Clarity Western Enhanced Chemiluminescence Substrates (Cat# 1705061), blotting-grade blocker (Cat# 1706404), and polyvinylidene difluoride (PVDF) membranes (Cat# 1620177) were purchased from Bio-Rad. All other chemicals and organic solvents of analytical grade were purchased from Thermo Fisher Scientific, VWR, or Sigma-Aldrich. All laboratory supplies and reagents used in our protocols are DNase and RNase free.
Cloning of BioRNA expression plasmids
Novel BioRNAs with payload miRNAs (Supplemental Table S1) as well as control BioRNAs, htRNAGly/sephadex-aptamer (BioRNAGly Control RNA) and htRNALeu/sephadex-aptamer (BioRNALeu Control RNA) were designed as reported (Li et al. 2021). In short, human tRNA (Genomic tRNA Database [GtRNAdb]; Chan and Lowe 2016) and miRNA (miRbase; Griffiths-Jones et al. 2008; Kozomara et al. 2019) sequences were obtained to clone BioRNA expression plasmids (Fig. 1B) by following two basic strategies to amplify target inserts (Supplemental Fig. S1). Inserts for BioRNALeu were obtained directly through PCR amplification using primers with 16-nt complementary base pair overlaps (IDT) (Supplemental Table S2). The inserts were cloned into the pBSKrna vector, linearized by endonucleases EcoRI-HF (Cat# R3101) and PstI-HF (Cat# R3140; New England Biolabs) with an In-Fusion Cloning Kit (Cat# 638949; Takara Bio), following the manufacturer's instructions. Inserts for BioRNAGly were produced through PCR amplification using the respective BioRNALeu/miRNA plasmids as a template and htRNAGly specific primers (Supplemental Table S2), and similarly cloned into a linearized pBSKrna vector. The ligation products (5 μL) were individually transformed into HST08 E. coli Stellar Competent Cells (Cat# 636766; 30 μL) (Takara Bio) and inoculated in Luria-Bertani (LB) media (400 μL) without selection on a shaker for 60 min (37°C, 225 rpm). Inoculated LB media (Cat# DF0446-17-3; Fisher Scientific) was plated on LB-ampicillin (100 μg/mL) agar plate and incubated overnight at 37°C. Three positive colonies were chosen and transferred separately to 15 mL of LB-ampicillin (100 μg/mL) medium and incubated overnight on a shaker (37°C, 225 rpm). Plasmids were individually extracted from 2 mL of the media via QIAprep Spin Miniprep Kit (Qiagen) and subjected to DNA sequencing (Genewiz from Azenta Life Sciences). Sequence-verified BioRNA expression plasmids, namely, pBSK/BioRNA, were preserved at −80°C for future use.
Small- and large-scale overexpression of target BioRNAs in E. coli
Small- and large-scale BioRNA production was conducted as previously detailed (Li et al. 2021), to confirm target BioRNA overexpression and further purification, respectively. In brief, E. coli transformed with pBSKrna/BioRNA plasmids were cultured in 2×YT media (Cat# BP9743-500; Fisher Scientific) for small- (15 mL) or large-scale (250 mL) production and incubated on a shaker (225 rpm and 37°C for 16 h). Total RNAs were isolated from centrifuged (10,000g and 4°C for 2 min) bacteria pellets through Tris–HCL (magnesium acetate [10 mM]–Tris–HCl)-saturated phenol (pH 4.7) extraction. The aqueous phase supernatant of the extraction was mixed with NaCl (5 M) (10% of supernatant volume), centrifuged (10,000g and 4°C for 10 min), and RNA contained supernatant was precipitated with ethanol (ethanol/sample solution = 2/1, v/v; −80°C for 60 min). Precipitated RNA was centrifuged (10,000g and 4°C for 10 min), RNA pellets were dried, and resuspended in autoclaved diethylpyrocarbonate (DEPC)-treated H2O for small- (500 μL) and large-scale (3 mL) production. For large-scale, resuspended RNAs were centrifuged (16,000g and 4°C for 15 min) and filtered through a 0.22 μm sterile syringe filter. Total RNAs were quantified with a Tecan SPARK running SPARKCONTROL, Nucleic Acid Quantification software (v2.3). Urea–PAGE analyses were performed to verify individual BioRNA levels in total RNAs (300 ng in 10 μL) (Fig. 1C,D) using an 8% polyacrylamide–urea (8 M) denaturing gel (urea–PAGE) in fresh 0.5× tris–acetate–ethylenediamine tetra acetic acid (TAE) buffer (50 min at 120 V). Urea–PAGE gels were incubated with ethidium bromide (0.5 μg/mL) for 5 min on a rocker and visualized under UV detector using the ChemiDoc MP Imaging System (Bio-Rad).
Purification and quality control of target BioRNAs
BioRNAs were purified by anion exchange FPLC as previously detailed (Li et al. 2021). In brief, total RNAs (5 mg/injection) were separated on an ENrichTM Q 10 × 100 column (Cat# 7800003) by using an NGC Quest 10 Plus Chromatography FPLC system (Bio-Rad). PVDF durapore membrane (0.22 μm) filtered and sonicated (20 min) mobile phases consisting of buffer A (10 mM NaH2PO4) and buffer B (10 mM NaH2PO4, 1 M NaCl) were used for elution, each adjusted pH to 7.0 with NaOH. The FPLC gradient schematic (flow rate 2 mL/min) was as follows: 100% buffer A 0–5 min; 55% buffer B 5–10 min; 55%–75% buffer B 10–40 min followed by washing with 100% buffer B and re-equilibrating with 100% buffer A before a subsequent injection. The FPLC injection loop was washed with autoclaved DEPC-treated H2O (6 mL) in between purification of individual BioRNAs. FPLC traces were monitored using a UV/Vis detector (260 nm) (Fig. 2A), and peak areas were used to evaluate the relative levels of BioRNA within the total RNA, which were consistent with the relative levels of BioRNA estimated from the initial urea–PAGE analyses. Fractions of target BioRNA peak were collected and verified by urea–PAGE analysis (Fig. 2B). Pure fractions were pooled, precipitated (ethanol/sample solution = 2/1, v/v), resolubilized with sterile DEPC treated H2O, and desalted and concentrated (2 mL sterile autoclaved DEPC treated H2O, 3×) using Amicon ultra-0.5 mL centrifugal filters (30 kDa; Cat# UFC203024; Sigma-Aldrich) following manufacturer's instructions. Purified BioRNAs were quantified using a Spark plate reader. Urea–PAGE was carried out to further verify individual final BioRNAs (50 ng in 10 μL) (Fig. 2D,E).
The quality of each BioRNA (Table 1) was determined by quantification of purity with HPLC and measurement of endotoxin level, as previously detailed (Li et al. 2021). In brief, individual BioRNA purities were quantitated using an XBridge OST C18 column (2.1 × 50 mm, 2.5 μm particle size, maintained at 60°C; Cat# 186003953; Waters) on a Shimadzu LC-20AD HPLC system. Individual BioRNAs were diluted in 100 μL of DEPC (10 ng/μL). BioRNA (5 μL) was injected and eluted (flow rate 0.2 mL/min) with buffer C (8.6 mM TEA, 100 mM Hexafluoro-2-propanol [HFIP] in HPLC grade H2O) and buffer D (8.6 mM TEA, 100 mM HFIP in HPLC grade methanol) as follows: 16% buffer D 0–1 min, and 16%–22% buffer D at 1–21 min followed by system washing with 22% buffer D at 21–22 min and re-equilibration with 16% buffer D. RNAs were detected using a photodiode array detector (260 nm) (Fig. 2C), and purity was determined by dividing the area under the BioRNA peak by all peaks displayed on the HPLC chromatogram. Endotoxin levels were measured with the Pyrogent-5000 kinetic LAL assay (Cat# N383; Lonza), following the manufacturer's instructions. BioRNA (100 ng/μL) and quantification standards were solubilized and diluted in endotoxin-free H2O, as instructed. The assay was performed over 40 min to assess turbidity (340 nm) as a readout of endotoxin levels using a SpectraMax iD5 plate reader (Molecular Devices). Provided endotoxin standards were used to generate a standard curve to determine endotoxin activity. BioRNAs with high purity (>98% by HPLC) and low endotoxin activity (<10 EU/μg RNA) were used for this study.
Cell culture
Human NSCLC lung carcinoma A549 (Cat# CRM-CCL-185) and H1975 (Cat# CRL-5908), and pancreatic carcinoma AsPC-1 (Cat# CRL-1682) cell lines were purchased from American Type Culture Collection. Cells were maintained in RPMI medium 1640 supplemented with 10% fetal bovine serum (Thermo Fisher Scientific) at 37°C in a humidified atmosphere with 5% CO2 and tested for mycoplasma contamination on a bimonthly basis.
CellTiter-Glo cell viability assay
A549, H1975, and AsPC-1 cells were individually seeded in flatbottom 96-well plates at 5000, 8000, and 6000 cells/well, respectively, as calculated with the Countless II FL system (Life Technologies). After RNA (15 nM) transfection using Lipofectamine 3000 (Vehicle) for 72 h with four biological replicates (N = 4/group), and cell viability values were determined in proxy by luminescence using the CellTiter-Glo 2.0 Cell Viability Assay Kit per manufacturer's instructions (Promega). Luminescence per well was determined by a SpectraMax iD5 plate reader. The viability of cells transfected with respective control RNAs was defined as 100%. Each experiment/study was repeated at least twice, and similar results were obtained.
RNA isolation and reverse transcription, quantitative real-time PCR analysis
NSCLC cells were seeded in six-well plates (500,000 cells/well), incubated overnight, and transfected with 15 nM of BioRNAGly/miR-7-5p, BioRNALeu/miR-7-5p, BioRNA Control RNA, mimic hsa-miR-7-5p, mimic negative control, or Vehicle for 48 h with three biological replicates (N = 3/group). Total RNA was extracted using Direct-zol RNA isolation kit (Zymo Research), quantified with a Spark microplate reader, and total RNA (500 ng) was used for cDNA synthesis with random hexamers or miR-7-5p stem–loop primer (Supplemental Table S3) by reverse transcription (RT) using NxGen M-MuLV reverse transcriptase (Cat# 30222-1; Lucigen). Quantitative real-time PCR (qPCR) analyses were carried out on a CFX96 Touch Real-Time PCR System (Bio-Rad) using gene-specific primers (Supplemental Table S3) and iTaq Universal SYBR Green Supermix (Bio-Rad) according to the manufacturer's protocols. Each individual biological replicate was assessed with two technical replicates. Levels of mature miR-7-5p were normalized to U6 snRNA in corresponding samples and determined using the formula 2−ΔΔCT. Each experiment/study was repeated at least twice, and similar results were obtained.
Protein isolation and western blot analysis
Cells were seeded into six-well plates (500,000 cell/well) and transfected with 30 nM BioRNAGly/miR-7-5p, BioRNALeu/miR-7-5p, BioRNA Control RNA, mimic hsa-miR-7-5p, mimic negative control (Supplemental Table S3), or Vehicle for 72 h with three biological replicates (N = 3/group). Cells were lysed by using Pierce RIPA buffer supplemented with complete protease inhibitors for 30 min on ice. Centrifuged supernatant (12,000g and 4°C for 10 min) was transferred to a 1.7 mL microcentrifuge tube and protein concentrations were determined with a BCA Protein Assay Kit. Whole-cell proteins (20 μg/lane) were separated via gel electrophoresis on a 7.5% (MRP1 and VDAC1) or 10% (EGFR) TGX Stain-Free SDS-PAGE gel. Protein gels were electrophoretically transferred onto methanol-activated polyvinylidene fluoride (PVDF) membranes using the Trans-Blot Turbo Transfer System (Bio-Rad). Membranes were imaged for total protein with a ChemiDoc MP Imaging System, blocked with 5% blotting-grade blocker, and incubated overnight at 4°C with primary antibodies against target proteins EGFR (1:1000; Cat# 4267S; Cell Signaling Technology), MRP1/ABCC1 (1:1000; Cat# 14685S; Cell Signaling Technology), VDAC1 (1:1,000; Cat# ab15895; Abcam), or β-actin (1:1,000; Cat# A5441; Sigma-Aldrich). Membranes were incubated (2 h, 25°C) with anti-rabbit (1:10,000; Cat# 111-035-003; Jackson ImmunoResearch Inc.) or anti-mouse IgG HRP-linked (1:3,000; Cat# 7076S; Cell Signaling Technology) secondary antibody. Clarity Western Enhanced Chemiluminescence Substrates were mixed (1:1) and applied to develop and image the membrane using a ChemiDoc MP Imaging System. The intensity values of the protein bands were determined by the Image Lab software (Bio-Rad) and normalized to corresponding β-actin and total protein for comparison. Each experiment/study was repeated at least twice, and similar results were obtained.
Computation modeling of BioRNA 3D structures
Predictive modeling of novel BioRNA and BioRNA derivatives was performed using sequences (Supplemental Table S4) attained from the GtRNAdb, our previous publication (Ho and Yu 2016), and miRbase.org (Griffiths-Jones et al. 2008; Kozomara et al. 2019). Tertiary structure was predicted using RNAComposer: Automated RNA Structure 3D Modeling Server (Popenda et al. 2012; Antczak et al. 2017) with incorporated secondary structure prediction method using RNAFold WebServer (Institute for Theoretical Chemistry, University of Vienna). The resulting prediction was manipulated using ChimeraX Next-Generation Molecular Visualization software (Resource for Biocomputing, Visualization, and Informatics, University of California, San Francisco). The sequence selection application was used to color code each BioRNA derivative. BioRNA and BioRNA derivatives include: htRNA (black), hsa-premiR-34a (blue), miRNA guide sequence (antisense; red), passenger sequences (light blue or light gray), five-prime (5′; magenta), three-prime (3′; white). Nucleotide code includes adenine (red), thymine (blue), guanine (green), and cytosine (yellow).
Statistics
Values are mean ± standard deviation (SD) and all data were analyzed with one-way ANOVA with Bonferroni post-tests (Prism, GraphPad Software). Difference between analyzed groups was considered as statistically significant when the probability value (P-value) was less than an α level of 0.05 (P < 0.05).
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
This study is supported by the National Cancer Institute (R01CA225958 and R01CA253230) and the National Institute of General Medical Sciences (R35GM140835), the National Institutes of Health (NIH). G.M.T. was supported by a Pharmacology Training Program Grant (T32GM099608 and T32GM144303) from the National Institutes of General Medical Sciences, NIH. The authors appreciate the access to the Molecular Pharmacology Shared Resource funded by the UC Davis Comprehensive Cancer Center Support Grant awarded by the National Cancer Institute (P30CA093373), NIH.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079904.123.
-
Freely available online through the RNA Open Access option.
- Received November 28, 2023.
- Accepted February 19, 2024.
This article, published in RNA, is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Gavin M. Traber is the first author of this paper, “Novel RNA molecular bioengineering technology efficiently produces functional miRNA agents.” Gavin is a graduate student in the laboratory of Ai-Ming Yu at the University of California, Davis School of Medicine Department of Biochemistry and Molecular Medicine. The major focus of his research is on RNAi anti-cancer pharmacology and experimental therapeutics to translate noncoding RNAs into new forms of therapy, investigate the mechanistic actions of microRNAs in the regulation of cancer cellular processes, and progress their novel RNA bioengineering technologies and syntheses of recombinant RNAs for basic research and drug development.
What are the major results described in your paper and how do they impact this branch of the field?
Our study demonstrates the reliability and robustness of our novel RNA molecular bioengineering platform technology to allow in vivo fermentation production of target BioRNA/miRNA agents by using human glycyl or leucyl tRNA fused hsa-pre-miR-34a stable carriers. Our BioRNAs are produced with a high yield, high degree of homogeneity, and 100% success rate to closely represent the properties of natural miRNA molecules. Comparisons between glycyl and leucyl stable carriers were overall equivalent with highly similar predicted 3D structures for the BioRNAs with noticeable differences in htRNA scaffolds and putative cell line-specific differences in antiproliferative activities. Use of the model BioRNA/miR-7-5p validates the intracellular release of miRNA payload and confirms the effectiveness of BioRNA/miRNA to regulate the expression of targeted genes in human NSCLC cells. Lastly, we validated the use of our BioRNA technology as a functional and comparable alternative to commercially available miRNA mimics for basic research as well as potential for therapy.
What led you to study RNA or this aspect of RNA science?
During my undergraduate studies at Harding University in Searcy, Arkansas, I was introduced to the impacts and biological functions of both coding and noncoding RNAs by my genetics professor, Dr. Rebekah Rampey. There I spent time studying the growing classes of noncoding RNAs and learning from giants in the RNA-based regulatory field such as Drs. Jennifer Doudna, David Bartel, Mark A. Kay, and Craig Mello.
During the course of these experiments, were there any surprising results or particular difficulties that altered your thinking and subsequent focus?
Initially, based on preliminary data in the lab suggesting that human glycyl tRNA fused hsa-pre-miR-34a exhibited a lower cytotoxicity while showing higher yield in producing target BioRNAs than other htRNAs, we thought the glycyl tRNA fused hsa-pre-miR-34a would outperform the leucyl tRNA version. Instead, our evidence suggests that both htRNA/pre-miR-34a carriers demonstrated roughly equal production efficiencies. Further, both versions of BioRNAs demonstrated comparable effects on cell viability with evidence of cell line dependence. Therefore, we expected similar outcomes from our functional studies of model BioRNA/miR-7-5p. However, we instead found BioRNAGly/miR-7-5p to outperform BioRNALeu/miR-7-5p in both the release of payload miRNA and their efficacy to modulate target gene expression. Although the limiting factor of BioRNALeu/miR-7-5p was not determined, further exploration into their structure by computational predictive modeling resulted in highly similar overall structures with only noticeable differences in the structure of their respective htRNA subcomponents.
What are some of the landmark moments that provoked your interest in science or your development as a scientist?
My passion for science began in the 5th grade in my hometown of La Crescenta, California while learning about cellular structure and substructure, as well as how these basic units of life come together, organize, reproduce, and function to sustain life. At the same time, I was captivated by the work of naturalist Sir David Attenborough, through his novels and documentaries as well as Mr. William (Bill) Nye, who took the intricacies within many scientific concepts and processes and made them approachable for young and aspiring scientists at the time, such as myself. This was a pivotal time for me, as it oriented my drive into my undergraduate studies where exposure to basic scientific research and study design propelled me to explore both academic- and industrial-based research as a technician at Stanford University School of Medicine, and with what was formally Google Accelerated Science, respectively, prior to beginning my graduate studies.
If you were able to give one piece of advice to your younger self, what would that be?
Looking back, I would urge my younger self to ask more questions earlier on and independent of the chance or likelihood of being wrong, as this is where I now believe the true scientist lives. I would further say to live more closely to the saying of another professor of mine from Harding University, Dr. Nathan Mills, “There is nothing wrong with a little open and honest investigation,” as well as the words of astrophysicist Dr. Neil deGrasse Tyson, “Whether or not you can ever become great at something, you can always become better at it.”
Are there specific individuals or groups who have influenced your philosophy or approach to science?
Specifically, I owe my scientific curiosity as well as philosophy and approach to science to my undergraduate professors at Harding University Dept. of Biology, naturalist Sir David Attenborough, Mr. William (Bill) Nye, astrophysicist Dr. Neil deGrasse Tyson, my former mentors Drs. Daniel Hornburg and Sara Ahadi, and chiefly my primary investigator and mentor for my graduate training, Dr. Ai-Ming Yu.
What are your subsequent near- or long-term career plans?
My near-term career plans are to complete my dissertation and graduate from the University of California, Davis in June of 2024. Further, my long-term plans are to pursue a career as a research scientist in the San Francisco-San Jose Bay Area of California, and to use my skills to contribute to a better understanding of the molecular roots behind disease at an institution focused on the development and translation of basic science and research to helping the lives of patients, and ultimately pass along my knowledge and experiences to coming generations of future scientists.








