
Diversity and modularity of tyrosine-accepting tRNA-like structures
- 1Department of Biochemistry and Molecular Genetics, University of Colorado Anschutz Medical Campus, Aurora, Colorado 80045, USA
- 2RNA Bioscience Initiative, University of Colorado Anschutz Medical Campus, Aurora, Colorado 80045, USA
- Corresponding author: jkieft{at}nysbc.org
-
Handling editor: Adrian Ferre-D'Amare
Abstract
Certain positive-sense single-stranded RNA viruses contain elements at their 3′ termini that structurally mimic tRNAs. These tRNA-like structures (TLSs) are classified based on which amino acid is covalently added to the 3′ end by host aminoacyl-tRNA synthetase. Recently, a cryoEM reconstruction of a representative tyrosine-accepting tRNA-like structure (TLSTyr) from brome mosaic virus (BMV) revealed a unique mode of recognition of the viral anticodon-mimicking domain by tyrosyl-tRNA synthetase. Some viruses in the hordeivirus genus of Virgaviridae are also selectively aminoacylated with tyrosine, yet these TLS RNAs have a different architecture in the 5′ domain that comprises the atypical anticodon loop mimic. Herein, we present bioinformatic and biochemical data supporting a distinct secondary structure for the 5′ domain of the hordeivirus TLSTyr compared to those in Bromoviridae. Despite forming a different secondary structure, the 5′ domain is necessary to achieve robust in vitro aminoacylation. Furthermore, a chimeric RNA containing the 5′ domain from the BMV TLSTyr and the 3′ domain from a hordeivirus TLSTyr are aminoacylated, illustrating modularity in these structured RNA elements. We propose that the structurally distinct 5′ domain of the hordeivirus TLSTyrs performs the same role in mimicking the anticodon loop as its counterpart in the BMV TLSTyr. Finally, these structurally and phylogenetically divergent types of TLSTyr provide insight into the evolutionary connections between all classes of viral tRNA-like structures.
Keywords
INTRODUCTION
Viruses have evolved many strategies to promote translation and replication of their genome while evading host cell antiviral surveillance pathways. In many cases, specifically structured RNA elements in the viral genome's noncoding regions play critical roles during infection, such as promoting viral protein synthesis and preventing viral RNA from degradation (Dreher 2010; Moon et al. 2012; Jaafar and Kieft 2019). Examples of multifunctional viral RNA elements are the tRNA-like structures (TLS) in many positive-sense, single-stranded, plant-infecting viruses (Hall 1979; Dreher 2010), and a few insect-infecting viruses (Gordon et al. 1995; Sherlock et al. 2021). These RNA elements are located at the termini of the 3′ UTR within the viral RNA and have been implicated in enhancement of translation, protection from 3′ to 5′ degradation, viral replication, and viral packaging (Joshi et al. 1985; Dreher and Hall 1988; Rao et al. 1989; Mans et al. 1992; Osman et al. 2000; Matsuda and Dreher 2004; Dreher 2009; Rao and Kao 2015).
As their name suggests, TLSs contain features that structurally mimic canonical tRNA, enabling interactions with certain host factors that normally process cellular tRNAs (Litvak et al. 1973; Rietveld et al. 1984; Goodwin and Dreher 1998; Hammond et al. 2009; Colussi et al. 2014; Bonilla et al. 2021). Notably, viruses that contain an authentic TLS have a canonical tRNA “CCA” tail appended to the 3′ end of a pseudoknot that mimics the acceptor stem, and they undergo aminoacylation by a host aminoacyl tRNA synthetase (aaRS) (Rietveld et al. 1984; Pleij et al. 1985; Dreher and Hall 1988; Bonilla et al. 2021; Langeberg et al. 2021; Sherlock et al. 2021).
TLSs are classified by the amino acid they are charged with, which correlates with sequence and structural features unique to each of the three classes. Valine-accepting tRNA-like structures (TLSVal) are found in the Tymoviridae, Virgaviridae, and Alphatetraviridae families and most closely resemble canonical tRNA with elements mimicking the D, T, and anticodon stems of tRNAVal (Giegé et al. 1978; Dreher et al. 1992; Colussi et al. 2014; Hartwick et al. 2018; Sherlock et al. 2021). Histidine-accepting tRNA-like structures (TLSHis) are found in Virgaviridae and Tymoviridae and retain some canonical tRNA features (D-loop/T-loop mimic) but are larger than tRNAs and contain certain secondary structure elements that are dissimilar to tRNA, including a long anticodon stem mimic with a large asymmetric internal loop (Rietveld et al. 1984; Joshi et al. 1985; Langeberg et al. 2021). Tyrosine-accepting tRNA-like structures (TLSTyr) are found in Virgaviridae and Bromoviridae, contain even fewer features of canonical tRNA, and are much larger in size than tRNAs (Loesch-Fries and Hall 1982; Dreher and Hall 1988; Felden et al. 1994; Bonilla et al. 2021).
Recently, structures of the TLSTyr from brome mosaic virus (BMV) in both the unbound (RNA-only) and bound (RNA–aaRS complex) forms were determined using single-particle cryo-electron microscopy (cryoEM) (Bonilla et al. 2021). Models of the BMV TLS resulting from these cryoEM reconstructions revealed the overall architecture of the seven helical domains of the RNA as well as an unexpected binding mode with the host tyrosyl-tRNA synthetase (TyrRS) involving the anticodon recognition domain (Fig. 1). Specifically, in the unbound form, the BMV TLS anticodon-mimicking domain is mobile, sampling a variety of positions and is thus not preorganized to interact with the TyrRS (Fig. 1C; Bonilla et al. 2021). However, in the BMV TLS–TyrRS complex, this domain is stably placed and oriented such that it is nearly parallel with the acceptor stem, contrasting with the perpendicular orientation resulting from the L-shape of tRNA (Fig. 1D).

Domain architecture of tyrosine-accepting TLS RNAs. (A, B) Cartoon diagrams of the secondary structure of the tyrosine-accepting tRNA-like structures (TLSTyr) from the (A) anulavirus, bromovirus, cucumovirus, and oleavirus genera of Bromoviridae and (B) hordeivirus genus of Virgaviridae. Helical elements are lettered according to previous literature (Ahlquist et al. 1981; Solovyev et al. 1996; Savenkov et al. 1998; Bonilla et al. 2021). The respective 5′ domains, consisting of the (A) E and B3 stem–loops or (B) R stem–loop, are boxed in gray. The secondary structure of the R domain of the hordeivirus TLSTyr was not experimentally probed before this study; the putative structure is shown. (C, D) Structural model of the BMV TLS in the C unbound (RNA-only; PDB ID: 7SAM) and (D) bound (RNA-tyrosine synthetase complex; PDB ID: 7SCQ) states with each domain labeled. The synthetase is not shown for clarity.
As mentioned above, TLSTyr is also found in viruses in the Virgaviridae family, and these TLSTyr representatives contain key differences compared to the BMV TLS. Specifically, all three members of the hordeivirus genus within Virgaviridae—barley stripe mosaic virus (BSMV), poa semilatent virus (PSLV), and lychnis ringspot virus (LRSV)—contain TLSTyrs in their 3′ UTRs (Solovyev et al. 1996). The most notable difference between the hordeivirus TLSTyrs and the BMV TLS (and related structures in other members of Bromoviridae) is that the former do not contain the E and B3 stem–loops (Fig. 1A). This is puzzling as these elements function as the BMV TLS's anticodon-mimicking domain (Felden et al. 1994; Bonilla et al. 2021). Instead, the 5′ domain for hordeivirus TLSs was proposed to comprise a large stem–loop structure with an asymmetric internal loop, which we label “R” (Fig. 1B) to remain consistent with previous nomenclature (Kozlov et al. 1984; Solovyev et al. 1996; Savenkov et al. 1998). Despite these differences, the aminoacylation of the BSMV genome with tyrosine at its 3′ terminus was demonstrated over four decades ago (Agranovsky et al. 1981, 1982; Loesch-Fries and Hall 1982). However, key features of these divergent TLSTyr, such as the minimal tyrosylation-competent element and therefore the necessary domains within the 3′ UTR that comprise the TLS, have not been experimentally determined. Also, while secondary structure models for the 3′ UTR, including the putative TLS domain, have been proposed for all three members of the hordeivirus genus (Kozlov et al. 1984; Agranovsky et al. 1992; Solovyev et al. 1996; Savenkov et al. 1998), these have not been probed experimentally.
The discrepancy as to the nature of a putative anticodon-mimicking domain in the TLSTyr representatives in hordeivirus as well as other subtle differences compared to TLSTyrs in Bromoviridae, motivated us to experimentally interrogate the structural features of these RNAs. Chemical probing data combined with functional assays enabled us to compare domains of hordeivirus TLSTyrs with those of BMV in the context of its known structure. We have assigned the role of anticodon mimicry to the 5′ domain of hordeivirus TLSTyr, which is required for tyrosylation and much larger than its counterpart in BMV. We then identified the most highly conserved features and areas of divergence among all TLSTyr examples to illuminate the most critical features for aminoacylation function across this diverse TLS class.
RESULTS AND DISCUSSION
Experimental exploration of hordeivirus TLSTyrs secondary structures
We first experimentally interrogated the proposed hordeivirus TLSTyrs secondary structures compared to Bromoviridae TLSTyrs. Selective 2′ hydroxyl acylation by primer extension (SHAPE) probing was used to evaluate the previously proposed structures by mapping the reactivity of the chemical modifier for each RNA onto its secondary structure model. Overall, the chemical probing data for all three hordeivirus TLSs (Fig. 2A; Supplemental Fig. S1) are consistent with previously proposed secondary structure predictions based solely on sequence information (Kozlov et al. 1984; Agranovsky et al. 1992; Solovyev et al. 1996). Notably, the 5′ portion of all three hordeivirus TLSTyr folds into a long stem–loop containing an asymmetric internal loop and a single bulged nucleotide; hereafter we refer to the 5′ region as the R domain. The remaining portion of the hordeivirus TLSsTyr folds in a manner that is largely consistent with Bromoviridae TLSTyr RNAs, with the base-paired regions in A, B1, B2, and C all displaying low reactivity to the chemical modifier. Notably, the pyrimidine-rich linker between B2 and the closing of the acceptor stem pseudoknot (A) shows low reactivity in all three hordeivirus TLSTyrs. The pyrimidine-rich linker region in the BMV TLSTyr also demonstrates low chemical probing reactivity, likely due to a stable conformation against the B1 stem and the loop of the E stem as seen in the cryoEM structure (Bonilla et al. 2021).

Structure and function of hordeivirus TLSTyr. (A) Chemical probing of the TLS representative from BSMV RNA1 using the SHAPE reagent NMIA. Reactivity was background subtracted and normalized according to the reactivity of loop regions in hairpin structures (not shown) flanking the TLS structure on both the 5′ and 3′ ends. See Supplemental Table S1 for complete sequence details and Supplemental Figure S1 for chemical probing of additional hordeivirus TLSTyr RNAs. (B) Schematic of in vitro aminoacylation assay using in vitro transcribed RNA, recombinantly expressed and purified TyrRS, and 3H-labeled tyrosine. (C) Relative activity of TyrRS on various TLSTyr RNAs as measured by the covalent addition of radiolabeled 3H-tyrosine at their 3′ termini after a 30-min incubation. 3H incorporation was normalized to the wild-type (WT) BMV TLSTyr construct, which had been previously tested (Bonilla et al. 2021). The truncated constructs Δ(E + B3) or ΔR are missing their respective 5′ domains, as indicated by the dashed box in Figure 1, and begin 4 nt before the B1 stem. The PSLV full 3′-UTR RNA includes the entire sequence after the stop codon. The BSMV TLS RNAs with mutations to the B2 and R stem–loops (B2mut and Rmut) contain UUCG tetraloops in place of the native loop sequence (see panel A for the WT BSMV TLSTyr sequence).
Defining the minimal functional unit for the hordeivirus TLSTyr
We next determined which portions of hordeivirus 3′ UTR comprise the minimal unit competent for tyrosylation, as previous studies used the entirety of the 3′ UTR as a substrate (Agranovsky et al. 1982, 1992). We used an in vitro assay with recombinantly expressed and purified TyrRS previously shown to be active on the BMV TLSTyr in a dose-dependent manner, and inactive on a TLSVal (Bonilla et al. 2021). In this assay, aminoacylation with 3H-tyrosine leads to 3H-labeled RNA (Fig. 2B), which is quantified and normalized to the WT full-length BMV TLSTyr. Validating the assay, there was no measurable tyrosylation above the no RNA (negative) control for a BMV TLSTyr that is truncated to lack the E and B3 stem–loops (Fig. 2B), consistent with previous data showing the BMV TLSTyr is sensitive to mutations in the B3 loop (Bonilla et al. 2021).
We then tested three putative hordeivirus RNAs, each beginning 5 nt before the R domain. All were tyrosylated to a similar level to the BMV TLSTyr. Truncations to all three hordeivirus TLSTyrs that removed the R domain decreased tyrosylation to levels similar to the negative control. Thus, the R domain is critical for aminoacylation of the hordeivirus TLSTyrs, perhaps analogous to the BMV TLSTyr 5′ domain comprising the E and B3 stem–loops, referred to hereafter as “E + B3.” Furthermore, an RNA containing the entire 3′ UTR of PSLV is tyrosylated to a similar level as the PSLV TLSTyr that begins just before the R domain (Fig. 2C). For PSLV, the other RNA elements in the 3′ UTR between the stop codon and R domain are interspersed poly(A) stretches and a set of three pseudoknots that comprise the “upstream pseudoknot domain” (UPD) (Supplemental Fig. S2A). These results demonstrate that the R domain is a required functional component for the hordeivirus TLSTyrs, but the other elements in the 3′ UTR (discussed in greater detail below) are not necessary for aminoacylation.
To further test the specific involvement of the hordeivirus TLSTyr R stem in tyrosylation, we mutated its apical loop to a UUCG tetraloop. This mutation substantially decreased tyrosylation, further illustrating the importance of this element. In contrast, mutation of the hordeivirus TLSTyr B2 apical loop had a less deleterious effect on tyrosylation (Fig. 2C). These results are consistent with analogous mutations to the BMV TLS B3 and B2 apical loops, respectively (Bonilla et al. 2021).
Anticodon mimicry by the 5′ domain of hordeivirus TLSTyr
After confirming the secondary structure of the R domain (Fig. 2A) and determining that this region of the hordeivirus TLSTyr is a necessary component for aminoacylation (Fig. 2C), we next investigated whether the R domain participates in anticodon mimicry analogously to the B3 domain of BMV TLS (Felden et al. 1994; Bonilla et al. 2021). In the R domain, the putative anticodon-mimicking loop for BSMV (i.e., the apical loop) is an “UAAC” tetraloop, which differs slightly from the BMV B3 apical loop (UACA). Neither match the canonical tRNATyr GUA anticodon embedded in a 7-nt loop. Nonetheless, mutation of either the BSMV R domain or BMV TLSTyr B3 apical loops to UUCG tetraloops resulted in the loss of aminoacylation (Fig. 2C; Bonilla et al. 2021). Additionally, the R domain of hordeivirus TLSTyrs contains a much longer helical region (18 total base pairs) than the combined length of the E + B3 domains of BMV (12 total base pairs).
Despite these differences, we hypothesized that the R domain of the hordeivirus TLSs serves the same functional role as the E + B3 domain of BMV, including interactions with the anticodon-recognition domain of TyrRS. If so, they could be functionally interchangeable. To test this, we created chimeric BMV and BSMV TLSTyr RNAs by appending the E + B3 domain of BMV to the BSMV TLS (Fig. 3A), and the R domain of BSMV to the BMV TLS (Fig. 3B). We then tested whether these chimeric constructs were competent substrates for tyrosylation in vitro.
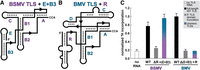
Aminoacylation of chimeric TLSTyrs. (A, B) Cartoon diagrams of the secondary structure of the TLSTyr from (A) BSMV with the E and B3 stem–loops of BMV appended to the 5′ end in lieu of the R domain and (B) BMV with the R stem–loop of BSMV appended to the 5′ end in lieu of the E + B3 domain. (C) Relative activity of TyrRS on WT, truncated (ΔR or Δ[E + B3], gray), and chimeric (+[E + B3] or + R, blue-purple) TLSTyr RNAs as measured by 3H-tyrosine incorporation. See legend to Figure 2 for additional details.
Again, the 5′ truncated BSMV TLSTyr without the R domain has no detectable aminoacylation activity above the no RNA negative control (Fig. 2C). However, the addition of the E + B3 domain to the BSMV hordeivirus TLSTyr in place of the R domain fully restores tyrosylatability (Fig. 3C). Thus, loss-of-function due to removal of the R domain is restored by adding the known anticodon-mimicking domain of the BMV TLS. Despite differences in size, primary sequence, and architecture, the R and E + B3 domains perform homologous functions. This could be accomplished by forming a similar structural architecture with the apical loop of the R domain contacting the anticodon recognition domain of the TyrRS enzyme similarly to the apical B3 loop. We speculate that the internal loop nucleotides in the R domain could allow it to be structurally flexible to contact the anticodon recognition domain of TyrRS, perhaps in a parallel conformation as observed for the BMV TLSTyr (Fig. 1D; Bonilla et al. 2021).
Interestingly, the other chimeric construct with the R domain of the BSMV appended to the 3′ portion of the BMV TLSTyr (Fig. 3B) did not restore aminoacylation activity (Fig. 3C). Chemical probing data show that this chimeric RNA is likely not globally misfolded (Supplemental Fig. S3). Rather, we hypothesize that the much larger R domain, which contains five additional base pairs compared to E and B3 combined, is somehow incompatible with the overall structure of the BMV TLSTyr. Some possible explanations are that the R domain is unable to adopt the correct conformation in the context of a TLSTyr containing both the C and D stem–loops. Or, in the BMV TLSTyr, there is necessary contact between E + B3 and B2 or the linker preceding the acceptor stem pseudoknot that cannot be formed by the R stem–loop due to its larger size or specific nucleotide identities. However, in the absence of a high-resolution structure of a hordeivirus TLSTyr RNA, it is difficult to determine whether steric clashes, the absence of critical tertiary contacts, or other factors lead to the inability of the chimeric BMV TLSTyr + BSMV R construct to be aminoacylated.
Unifying and distinguishing features between all viral tyrosine TLSs
To evaluate the conservation and variations across all known TLSTyrs, we created sequence and secondary structure alignments using 32 unique TLSTyr sequences from 13 distinct viruses in Bromoviridae, and the eight unique TLSTyr sequences from the three members of hordeivirus (Supplemental Files 1 and 2). We used R2R (Weinberg and Breaker 2011) to depict the consensus sequence and secondary structure models (Fig. 4A,B).

Conservation and phylogenetic distribution of viral tyrosine-accepting tRNA-like structures. (A) Consensus sequence and secondary structure model for the 32 unique TLSTyr sequences from 13 distinct viruses in Bromoviridae. (B) Consensus sequence and secondary structure model for the eight unique TLSTyr sequences from three distinct viruses in Virgaviridae. The stem–loop labeled as C/D was referred to as the C stem in previous literature; however, it may more closely resemble the optional D stem found in type 1 TLSTyr. (C) Phylogenetic distribution of all types of TLSs in the Virgaviridae and Bromoviridae families.
All TLSTyrs contain a highly conserved 3′ end; the identity of the discriminator base—the nucleotide immediately preceding the terminal CCA—is perfectly conserved as an adenosine, and the terminal base pair of the acceptor stem-mimicking pseudoknot is also perfectly conserved as a C–G. This high degree of conservation is perhaps unsurprising, given that the adenosine discriminator nucleotide and the C–G pair at the base of the acceptor stem are the most critical identity elements for recognition of canonical tRNATyr by TyrRS (Fechter et al. 2000, 2001; Tsunoda et al. 2007). While the R and E + B3 domains diverge between the two subclasses of TLSTyr, both contain tetranucleotide apical loops that typically start with a U followed by three A or C nucleotides. Again, this sequence differs from the tRNATyr anticodon sequence, but in tRNATyr the anticodon is a weaker identity element for TyrRS recognition compared to the acceptor stem and discriminator nucleotide (Fechter et al. 2001). Specifically, most mutations to the tRNATyr anticodon result in <10-fold loss in aminoacylation efficiency while mutations to the acceptor stem and discriminator nucleotide result in a >1000-fold decrease or complete loss in aminoacylation (Fechter et al. 2001). Although the identity of the nucleotides in the anticodon-mimicking loop (B3 for BMV; R for BSMV) differ from canonical tRNATyr, mutations to the loop sequence for TLSTyr decrease aminoacylation efficiency (Fig. 2C; (Bonilla et al. 2021).
An area of variability among TLSTyr is in the region located between the B1 stem and the A pseudoknot. In the BMV TLSTyr, this region comprises the C and D stem–loops, which are not universally conserved. The C stem–loop is always present in Bromoviridae TLSTyrs and contains elements critical for recognition by the RNA-dependent RNA polymerase (RdRP) to achieve viral replication (Rao et al. 1989; Chapman and Kao 1999; Rao and Kao 2015). The cryoEM structure of the BMV TLSTyr revealed that the C stem–loop is close in space to the A pseudoknot (Bonilla et al. 2021), consistent with its importance for replication as transcription begins at the 3′ end. The D stem–loop is not always present for Bromoviridae TLSTyrs (Ahlquist et al. 1981; Bonilla et al. 2021); when present, its function is unknown.
The hordeivirus TLSTyrs have a single stem–loop in this region, which we have labeled C throughout in accordance with previous literature, and are proposed to be missing a D stem–loop (Savenkov et al. 1998). However, the hordeivirus TLSTyrs C stem–loop differs significantly from the C stem–loop of Bromoviridae TLSs. First, the length of the helix in hordeiviruses (6 bp with a pentanucleotide apical loop) is dissimilar to the Bromoviridae TLSTyr C stem (10 bp with a large, asymmetric internal loop). Second, the proximity of the C stem to the 3′ end to perform its role in replication might not be relevant for the hordeivirus TLSTyr. As members of the Virgaviridae family, these viruses have a different RdRP lineage compared to Bromoviridae. The hordeivirus TLSTyrs might not rely on this additional C stem–loop, but instead contain recognition requirements for replication more consistent with the other Virgaviridae TLS classes (Singh and Dreher 1997; Deiman et al. 1998; Osman et al. 2000). We propose that the stem–loop in this region in hordeivirus TLSs might have more similarity to the BMV D stem–loop and have labeled it “C/D” (Fig. 4B) to note this ambiguity.
TLSTyrs differ in the structures directly upstream
An intriguing connection between the viruses within hordeivirus and other genera within Virgaviridae (Fig. 4C) is the overall architecture of the entire 3′ UTR upstream of the TLS domain. The TLSHis from tobacco mosaic virus (TMV) is also preceded by a UPD consisting of three consecutive pseudoknots connected by linker and loop nucleotides (Van Belkum et al. 1985; Zeenko et al. 2002). For TMV, a portion of the UPD—two of the pseudoknots and the intervening loop nucleotides—has been implicated as necessary for enhancing translation and is the binding site for elongation factor eEF1A (Zeenko et al. 2002). The translation enhancement effect of the UPD was increased synergistically with the 5′ cap structure and elements in the 5′ UTR (Leathers et al. 1993). Both BSMV and PSLV contain the same UPD architecture as TMV (Supplemental Fig. S2), including the highly conserved loop nucleotides and three pseudoknots (Van Belkum et al. 1985; Zeenko et al. 2002). Chemical probing data support the predicted secondary structure of this region for both BSMV and PSLV, and the presence of the UPD does not appear to affect the folding of the TLS (Supplemental Fig. S4) or be necessary for aminoacylation (Fig. 2C).
An additional layer of complexity in the hordeivirus 3′ UTRs are poly(A) sequences (Supplemental Fig. S2). Both BSMV (contains a UPD) and LRSV (no UPD) contain a poly(A) stretch of variable length (4–40 nt) directly after the stop codon (Agranovsky et al. 1982, 1992; Jackson et al. 1989; Solovyev et al. 1996), which is not required for replication (Zhou and Jackson 1996). In contrast, PSLV (contains a UPD) has several short (4–5 nt), interspersed poly(A) stretches between the stop codon and UPD (Agranovsky et al. 1992). It has been proposed (Dreher and Miller 2006) that 3′-UTR elements in other viruses within Virgaviridae might contribute to 5′–3′ end communication similarly to eukaryotic mRNAs—a role that poly(A) sequences in hordeivirus genomes could provide through poly(A) binding protein. However, the relative contributions of the TLS, UPD, and poly(A) features to viral translation enhancement have not been investigated in hordeivirus to date, and much remains to be unraveled in the function of these multifunctional, highly structured, noncoding RNA elements.
Concluding remarks
The data presented herein define the functional element necessary to achieve aminoacylation for hordeivirus tyrosine-accepting TLSs. We have shown that certain portions of TLSTyrs are similar among all representatives, with the highest degree of conservation at the extreme 3′ end. Other portions of the TLSTyr are much more variable and some features may be more relevant to functions other than aminoacylation. The 5′ domains of the two TLSTyr types appear dissimilar in terms of secondary structure, but our mutational analyses suggest a common function, which also demonstrates the modularity of RNA structure and function.
While the evolutionary trajectory of all TLS-containing viruses is difficult to definitively determine, it has been proposed that these RNAs originated in the Virgaviridae family, since all three known TLS classes are represented in this one viral family (Fig. 1A; Dreher 2010). While the amino acid specificity of the hordeivirus TLS class for tyrosine matches that of Bromoviridae, other features of the hordeivirus genus align more closely with other members of Virgaviridae, including the same UPD in some members of both hordeivirus and tobamovirus. The hordeivirus TLS R domain, which we propose functions as an anticodon mimic, has some intriguing similarities to the anticodon stem mimic of TLSHis: both have similar overall lengths, have an asymmetric internal loop, and the trinucleotide anticodon mimic contained in the apical loop is divergent from the 7-nt canonical tRNA anticodon loop. It is tempting to speculate that the original tyrosine-accepting TLSs could have emerged in hordeivirus, having diverged at some point from the histidine class in tobamovirus. These TLSTyr could have subsequently appeared in Bromoviridae, possibly through recombination. Regardless of the evolutionary trajectory, the presence of different TLS classes with other RNA elements in the 3′ UTR represents an intriguing set of RNA structures contributing to numerous functions in the viral lifecycle. Despite recent advances in understanding the structural tRNA mimicry of these TLS domains, the individual roles and potential synergistic effects of these viral RNA structures in translation, replication, and packaging remain largely mysterious.
MATERIALS AND METHODS
RNA consensus sequence and secondary structure model
An existing alignment (Rfam ID: RF01075) of two hordeivirus TLSs (BSMV RNA1 and PSLV RNA1) was obtained from the Rfam database version 13.0 (Kalvari et al. 2018). This preliminary alignment was extended on the 5′ end to include the R domain. This domain was previously excluded as it had not been confirmed as necessary for function. Sequences of additional examples of this class (BSMV RNA2 and RNA3, PSLV RNA2, LRSV RNA1 and RNA2) were retrieved from the National Center for Biotechnology Information (NCBI) viral nucleotide database (https://www.ncbi.nlm.nih.gov/labs/virus/vssi/#/), then manually added and aligned. A previously published alignment of Bromoviridae TLSs (Bonilla et al. 2021) was amended to only include one unique example per viral RNA (previously all 512 unique sequences were reported, which included many examples from different strains of the same virus) to be consistent with the alignment of hordeivirus TLSs, as described above. Additionally, the sequences from the tomato aspermy virus, which is not tyrosylated (Joshi and Haenni 1986) and therefore does not functionally belong to this class despite sequence and secondary structure similarity, were removed from the Bromoviridae TLS alignment. The consensus sequence and secondary structure models for the hordeivirus and Bromoviridae TLSTyr classes were separately calculated and visualized, using the two above alignments (hordeivirus: Supplemental File 1; Bromoviridae: Supplemental File 2) as input files, using R2R (Weinberg and Breaker 2011) and labeled in Adobe Illustrator.
RNA preparation
DNA templates were ordered as gBlock double-stranded DNA fragments (IDT) amplified by PCR in 100 μL volumes (see Supplemental Table S1 for all DNA templates and primer sequences used in this study). Reverse primers used to amplify the constructs for in vitro aminoacylation assays contained two 5′-terminal 2′OMe-modified bases to achieve a higher yield with the correct 3′ end of the construct. dsDNA amplification was confirmed by 1% agarose gel electrophoresis. Transcriptions were performed in 200 μL volume using 50 μL of PCR product as template dsDNA, 30 mM Tris pH 8.0, 60 mM MgCl2, 8 mM each NTP, 10 mM DTT, 0.1% spermidine, 0.1% Triton X-100, and T7 RNA polymerase. Reactions were incubated at 37°C overnight, then purified using denaturing 10% polyacrylamide gel electrophoresis (PAGE). Bands were visualized by UV shadowing, excised, sliced into small pieces, and soaked in ∼300 μL of diethylpyrocarbonate (DEPC)-treated milli-Q filtered water (Millipore) containing 20 mM sodium acetate (pH 5.3) at 4°C overnight to elute the RNA. Supernatant-containing RNA was subjected to ethanol precipitation, then resuspended in DEPC-treated water, diluted to the appropriate concentration, and stored at −20°C.
Chemical probing of RNAs in vitro
Structure probing experiments using the chemical modifier N-methyl isatoic anhydride (NMIA) were performed as described previously (Cordero et al. 2014; Langeberg et al. 2021; Sherlock et al. 2021). Briefly, RNA was refolded by heating to 90°C for 5 min, cooled to 23°C, then incubated with 10 mM MgCl2 for 20 min. Subsequently, the refolded RNA was modified by incubating with NMIA for 15 min at 23°C. NMIA modification reaction conditions and final concentrations: 120 nM RNA, 6 mg/mL NMIA or dimethyl sulfoxide (DMSO; negative control), 50 mM HEPES-KOH (pH 8.0), 10 mM MgCl2, 3 nM 5–6 FAM-labeled RT primer (see Supplemental Table S1 for sequence). The modification was quenched by the addition of NaCl to 500 mM, Na-MES buffer (pH 6.0) to 50 mM, and oligo (dT) magnetic beads [Invitrogen Poly(A) Purist MAG Kit], which hybridize to the poly(A) stretch contained in the RT DNA primer.
Chemically modified RNAs were recovered using oligo dT magnetic beads, washed with 70% ethanol, then resuspended in water. Reverse transcription was performed using the SuperScript III enzyme (Invitrogen). RNA ladders were produced by four separate reverse transcription reactions using ddNTPs. Reverse transcription reactions were incubated at 50°C for 45 min, then the RNA was degraded by adding NaOH (final concentration 200 mM), heating to 90°C for 5 min, then quenching with an acidic solution (final concentration: 250 mM sodium acetate pH 5.2, 250 mM HCl, 500 mM NaCl). The remaining DNA products were purified using the magnetic stand then washed with 70% ethanol. A solution containing HiDi formamide solution (Thermo Fisher) and spiked with GeneScan 500 ROX dye Size Standard (Thermo Fisher) was added to elute DNA products from the magnetic beads.
5′-FAM-labeled reverse-strand DNA products were analyzed by capillary electrophoresis using an Applied Biosystems 3500 XL instrument. Fragment size analysis, alignment, background subtraction, and normalization (based on reactivity in flanking stem–loop regions) were performed using the HiTRACE RiboKit (https://ribokit.github.io/HiTRACE/) (Kladwang et al. 2011; Yoon et al. 2011; Kim et al. 2013; Lee et al. 2015) in MatLab (MathWorks), and figures were rendered using RiboPaint (https://ribokit.github.io/RiboPaint/) in MatLab, then labeled in Adobe Illustrator. SHAPE reactivity was superimposed on the previously proposed secondary structure models (Van Belkum et al. 1985; Solovyev et al. 1996).
Aminoacylation assays
The TyrRS enzyme from a model host of BMV, Phaseolus vulgaris (common bean), used in the current study was recombinantly expressed and purified previously; details can be found in the Materials and Methods section of Bonilla et al. (2021). Aminoacylation constructs were refolded by heating to 90°C for 5 min, cooling to 23°C, then incubated with 10 mM MgCl2 for 20 min.
Aminoacylation reactions were set up by mixing 2 μL of 1 μM RNA or water, 1 μL of 10× aminoacylation buffer (10×: 300 mM HEPES-KOH [pH 7.5], 20 mM ATP, 300 mM KCl, 50 mM MgCl2, 50 mM DTT), 1 μL 3H-labeled l-tyrosine (60 Ci/mmol), 1 μL of 3 μM TyrRS enzyme (10×: 2 μM) and 5 μL of water (final reaction volume = 10 μL). Aminoacylation reactions, each performed in triplicate, were incubated at 30°C for 30 min, diluted by adding 200 μL of wash buffer (1×: 20 mM Bis-Tris [pH 6.5], 10 mM NaCl, 1 mM MgCl2, with trace xylene cyanol for visualization), then immediately loaded onto a vacuum filter Minifold I 96 well dot-blot apparatus (Whatman). Using the dot-blot system, the reaction solution was filtered through a layer of Tuffryn membrane (Pall Life Sciences), Hybond positively charged membrane (GE Healthcare), and gel dryer filter paper (Bio-Rad). Before the filter blotting apparatus assembly, each layer was equilibrated in 1× wash buffer without xylene cyanol. The RNA binds to the positively charged Hybond membrane. After application of the reaction solution, each blot was immediately washed three times with 300 μL of wash buffer containing trace xylene cyanol. The filters were subsequently dried and the blots from the Hybond membrane were excised and measured for 3H incorporation by liquid scintillation counter (PerkinElmer Tri-Carb 2910 TR). Data were analyzed and plotted using Microsoft Excel, then labeled in Adobe Illustrator.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
We thank Andrea MacFadden for assistance in purifying the TyrRS enzyme. We thank members of the Kieft laboratory for helpful discussions. This work was supported by the National Institutes of Health/National Institute of General Medical Sciences (NIH/NIGMS) grant R35GM118070 to J.S.K. M.E.S. was a Jane Coffin Childs Postdoctoral Fellow.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079768.123.
- Received July 8, 2023.
- Accepted December 12, 2023.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Madeline Sherlock is the first author of this paper, “Diversity and modularity of tyrosine-accepting tRNA-like structures.” Madeline is a Jane Coffin Childs postdoctoral fellow in the laboratory of Jeffrey Kieft, which was formerly located at the University of Colorado Anschutz Medical Campus and recently moved to the New York Structural Biology Center. Her research focus is on structure–function relationships of viral RNAs and how they influence translation.
What are the major results described in your paper and how do they impact this branch of the field?
This paper describes the structure and phylogenetic conservation of RNAs found at the 3′ end of certain viral genomes that mimic cellular tRNAs and become aminoacylated by tyrosine. We found that there are two distinct types of RNAs within this class of tRNA-like structures, which are found in distinct viral families. Some portions of the tRNA-like structure are highly similar between the two types, while others—in particular the domain that mimics the anticodon stem–loop—adopt different secondary structures but perform the same function to achieve tyrosylation.
What led you to study RNA or this aspect of RNA science?
This work builds on multiple previous publications from the Kieft lab studying the structure, function, and phylogenetic distribution of all classes of tRNA-like structures to understand how tRNA mimicry is achieved at the molecular level. I'm fascinated by these multifunctional viral RNAs and how they can interact with a range of host and viral proteins. I wanted to determine how this type of RNA folds and interacts with the tyrosine synthetase to understand how different structures can accomplish the same function.
What are some of the landmark moments that provoked your interest in science or your development as a scientist?
A huge moment for me, both personally and professionally, was receiving the Scaringe Graduate Student Award from the RNA Society in Berkeley in 2018. I presented during the opening session in front of a massive audience which was both terrifying and thrilling. For the rest of that week, I met so many new people who came up to me after seeing my talk, and I truly felt I had been embraced by the entire community. I haven't missed an RNA Society meeting since my second year of graduate school—it's my favorite conference and the field is incredibly supportive.
Are there specific individuals or groups who have influenced your philosophy or approach to science?
I've had the pleasure of working with amazing mentors over my career. My undergraduate research with Phil Bevilacqua sparked my passion for RNA research and shaped how I think about RNA folding. My graduate training with Ron Breaker helped me realize the power of using bioinformatics to evaluate genetic context and conservation to gain insight into RNA function, which is evident in this publication. During my postdoc with Jeff Kieft, I started thinking more like a structural biologist, learned all sorts of experimental techniques including cryoEM, and gained considerable confidence and independence in my research.
What are your subsequent near- or long-term career plans?
I am currently interviewing for positions to begin my independent career, so stay tuned for the future Sherlock lab.








