
Structural idiosyncrasies of glycyl T-box riboswitches among pathogenic bacteria
- Nikoleta Giarimoglou1,
- Adamantia Kouvela1,
- Jinwei Zhang2,
- Vassiliki Stamatopoulou1 and
- Constantinos Stathopoulos1
- 1Department of Biochemistry, School of Medicine, University of Patras, 26504 Patras, Greece
- 2Laboratory of Molecular Biology, National Institute of Diabetes and Digestive and Kidney Diseases, NIH, Bethesda, Maryland 20892, USA
- Corresponding author: cstath{at}med.upatras.gr
-
Handling editor: Adrian Ferré-D'Amaré
Abstract
T-box riboswitches are widespread bacterial regulatory noncoding RNAs that directly interact with tRNAs and switch conformations to regulate the transcription or translation of genes related to amino acid metabolism. Recent studies in Bacilli have revealed the core mechanisms of T-boxes that enable multivalent, specific recognition of both the identity and aminoacylation status of the tRNA substrates. However, in-depth knowledge on a vast number of T-boxes in other bacterial species remains scarce, although a remarkable structural diversity, particularly among pathogens, is apparent. In the present study, analysis of T-boxes that control the transcription of glycyl–tRNA synthetases from four prominent human pathogens revealed significant structural idiosyncrasies. Nonetheless, these diverse T-boxes maintain functional T-box:tRNAGly interactions both in vitro and in vivo. Probing analysis not only validated recent structural observations, but also expanded our knowledge on the substantial diversities among T-boxes and suggest interesting distinctions from the canonical Bacilli T-boxes. Surprisingly, some glycyl T-boxes seem to redirect the T-box trajectory in the absence of recognizable K-turns or contain Stem II modules that are generally absent in glycyl T-boxes. These results consolidate the notion of a lineage-specific diversification and elaboration of the T-box mechanism and corroborate the potential of T-boxes as promising species-specific RNA targets for next-generation antibacterial compounds.
Keywords
INTRODUCTION
T-box riboswitches are important conditional RNA regulators of transcription or translation in Gram-positive bacteria (Grundy and Henkin 1993; Henkin 2014; Sherwood et al. 2015). They constitute elaborate structures as part of 5′ UTR of mRNAs which encode for aminoacyl-tRNA synthetases, amino acid transporters, and genes involved in amino acid metabolism (Henkin 2008; Gutiérrez-Preciado et al. 2009). In contrast to the broader class of riboswitches that bind relatively small ligands, T-boxes bind cognate tRNAs which are metabolically related to the genes under control. Binding of the tRNA through specific interactions ensures a structural switch which enables the T-box to sense whether the tRNA is charged or uncharged with its cognate amino acid (Grundy and Henkin 1993; Grundy et al. 2002; Yousef et al. 2005; Green et al. 2010; Zhang and Ferré-D'Amaré 2014; Kreuzer and Henkin 2018). For many years, the regulatory mechanism of T-box:tRNA interactions, was portraited through secondary structures of varying lengths, stems, and single-stranded regions, some of which are phylogenetically conserved (Suddala and Zhang 2019a). Although T-boxes are widespread among Gram-positive bacteria and more than 20,000 putative sequences have been annotated, they have been extensively studied only in a few Bacilli species.
A typical T-box consists of an elongated Stem I domain, ∼100 nt long that contains a conserved AG bulge and an apical loop in the distal region (Fig. 1). Together, the AG bulge and the distal loop interact to create a flat platform termed interdigitated double T-loop motif (IDTM), which stacks against the tRNA elbow (Zhang and Ferré-D'Amaré 2013, 2015; Suddala and Zhang 2019b). The ability of Stem I to act as a tRNA molecular ruler is due to a small 2–3 nt long pyrimidine-rich flexible bulge, positioned between the AG bulge and the specifier loop (Zhang and Ferré-D'Amaré 2016; Suddala and Zhang 2019b). The apical loop is followed by the highly conserved specifier loop, in which the specifier sequence—a codon-like triplet that interacts with the tRNA anticodon—is positioned in a discrete location (Zhang and Ferré-D'Amaré 2015). This interaction is fundamental for the specific selection of the cognate tRNA against noncognates, and further modulates tRNA-binding affinity (Fig. 1; Caserta et al. 2015). A kink-turn motif (also known as a GA motif) is present underneath the specifier loop (Kreuzer and Henkin 2018). Although not necessary for the tRNA recognition by Stem I, K-turn can contribute to the correct orientation of the 3′ antiterminator domain, in order to facilitate the sensing of the tRNA aminoacylation status (Fig. 1; Green et al. 2010; Huang and Lilley 2013; Zhang and Ferré-D'Amaré 2013; Lilley 2014; Battaglia et al. 2019; Suddala and Zhang 2019b). The 5′ Stem I domain and the 3′ T-box antiterminator/antisequestrator domain are joined via a linker of variable length, depending on the presence or absence of the Stem II and Stem IIA/B pseudoknot (Zhang and Ferré-D'Amaré 2015). Located immediately after the linker and upstream of the antiterminator is the Stem III element. Although this structural element has variable length and sequence, it is present in all identified T-boxes so far. Stem III has two conserved purine-rich flanks on either side, necessary for tRNA binding, that follow the pattern 5′-RRRxG-Stem III-AA-3′, where x stands for any nucleotide. Stem III, together with its adjoining purines and the downstream antiterminator/antisequestrator domain, create the T-box discriminator, which preferentially binds uncharged tRNAs while discriminating against charged tRNAs (Li et al. 2019). The antiterminator of transcriptional T-boxes consists of two short helices, A1 and A2, joined by a highly conserved 7 nt bulge of the sequence 5′-UGGNACC-3′. The 4 nt 3′-NGGU-5′ sequence is complementary to the tRNA ligand's universal 3′-NCCA end. The ACC trinucleotide in the 3′ edge is also highly conserved, whereas the fourth nucleotide at the center of the T-box bulge covaries with the discriminator base of the cognate tRNA to form a 4 bp intermolecular helix between the T-box and the tRNA (Zhang and Ferré-D'Amaré 2015).
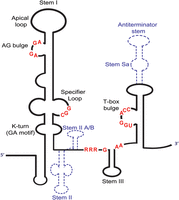
Illustration of a typical glycyl T-box riboswitch. The highly conserved nucleotides are indicated with red color. The stems that are not always present on the T-boxes are illustrated with blue dashed lines.
From previous detailed information derived from studies in Bacilli, it is evident that T-boxes from different species exhibit substantial sequence and structural diversification from the canonical “Bacillus rule” (Vitreschak et al. 2008; Wels et al. 2008; Gutiérrez-Preciado et al. 2009; Saad et al. 2013; Apostolidi et al. 2015; Wencker et al. 2021). For example, the Staphylococcus aureus glyS T-box contains an appended domain in the terminator/antiterminator region termed stem Sa. Stem Sa is important for tRNAGly isoacceptor discrimination and also for binding of mainstream antibiotics, features that make it an ideal target for new antibacterials (Apostolidi et al. 2015; Stamatopoulou et al. 2017; Giarimoglou et al. 2022a, b). Similarly, in the S. aureus Met-T-box, three additional stem–loops were identified in its extended linker region spanning 226 nt, the function of which is currently unknown (Wencker et al. 2021). Moreover, a peculiar T-box in Clostridium acetobutylicum responds to two different but metabolically related tRNAs for Asn and Glu through a larger specifier loop (Saad et al. 2012). These findings suggest that distinct T-box RNA structural features have evolved and been preserved to confer fitness to diverse bacteria species to adapt to their specific and variable environmental niches (Vitreschak et al. 2008).
In the present study, we report the secondary structures of transcriptional T-boxes that control the expression of glycyl–tRNA synthetase from four prominent human pathogens, Clostridium tetani, Listeria monocytogenes, Streptococcus pneumoniae, and Streptococcus pyogenes. Chemical probing and in vivo readthrough assays revealed important structural differences among the T-boxes which, interestingly, do not interfere with the ability of the T-boxes to interact both in vitro and in vivo with the tRNAGlyGCC ligand. Surprisingly, the K-turn motif, which has been considered indispensable for a functional T-box, is apparently absent from both streptococcal T-boxes, while the S. pneumoniae glyQS T-box contains an unusually short linker. Interestingly, the S. pyogenes glyQS T-box contains a Stem II domain that is typically absent from most other glycine-specific T-boxes. Collectively, the results highlight the structural diversity and functional competency of T-box riboswitches from different species, which in turn inform their divergent metabolic adaptation mechanisms and suggest novel RNA targets for the development of species-specific antibacterial compounds.
RESULTS
Deviations among glycyl T-boxes from pathogens in nonconserved domains
Glycyl T-boxes from C. tetani, L. monocytogenes, S. pneumoniae, and S. pyogenes were selected for structural and functional analysis. Multiple sequence alignment analysis including the “typical” Geobacillus kaustophilus and Bacillus subtilis T-boxes showed that all six T-boxes contain the major conserved structural and sequence features. These include a classical Stem I with the conserved triplet of the specifier loop, the AG bulge, and the Apical loop. The 5′ side of the specifier loop preserves the conserved motif 5′ AGUA 3′, except for the S. pyogenes T-box in which this motif is 5′ AGGA 3′. Additionally, the moderately conserved purines that are usually located upstream of this motif are generally present (Fig. 2). A pyrimidine-rich region with low conservation can be discerned between the specifier loop and the AG bulge. Typically, in this region a small 2–3 nt long pyrimidine-rich bulge exists that functions as a flexible hinge which allows Stem I to adapt to the shape or flexibility of the tRNA (Zhang and Ferré-D'Amaré 2016). The AG bulge is also present in all four T-boxes, exhibiting a high degree of conservation of the motif 5′ AGCGA 3′, which differs only in the C. tetani T-box (5′ AGAGAG 3′). This motif is highly conserved with few exceptions for the cytosine, which in the case of C. tetani glyS T-box is substituted by an adenosine. The Apical loop appears conserved in all T-boxes except for the S. pyogenes glyQS T-box which contains the 5′ GGUG 3′ motif. Interestingly, the Apical loops of the L. monocytogenes, S. pneumoniae, and G. kaustophilus glyQS T-boxes (80–88 nt in the alignment) (Fig. 2) are very similar in sequence. The region following the Apical loop exhibits low conservation in all cases. The specifier loop region generally contains a conserved 5′ GAA 3′ sequence which forms an S-turn motif. This sequence signature is curiously absent from the streptococcal glyQ T-boxes (positions 110–112 nt) (Fig. 2). In line with the previous idiosyncrasies is also the C. tetani specifier codon, which instead of GGC is a GGA. Right next to the specifier codon, a purine is present in all cases (“A” in the Bacilli glyQ T-boxes and “G” in the rest of them; position 118 nt in the alignment) (Fig. 2). This purine reinforces the 3 bp intermolecular helix between the specifier codon and tRNA anticodon, together with a purine that is located on the opposite side of the 3 bp intermolecular helix. This kind of interaction is analogous to the codon–anticodon duplex in the ribosome. At the proximal end of Stem I, a K-turn has been reported for the typical T-boxes and is suggested to be important for the orientation of the tRNA ligand as well as Stem II, when present. Interestingly, the nucleotides that support this K-turn formation appear to be missing from the streptococcal glyQS T-boxes.
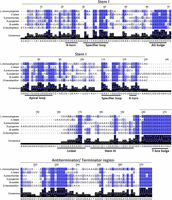
Multiple sequence alignment of the glycyl T-boxes from L. monocytogenes, C. tetani, S. pneumoniae, S. pyogenes, B. subtilis, and G. kaustophilus. The alignment was performed using Clustal Omega, and Jalview 2.11.1.7 was used for illustration purposes. For the alignment, the highly conserved nucleotides (80%–100%) are indicated by dark blue color, the moderately conserved nucleotides (50%–80%) are indicated with blue, while the less conserved nucleotides (≤50%) are indicated with light blue. Alignment numbering does not correspond to the numbering of the respective glycyl T-boxes.
Surprisingly, the S. pyogenes glyQS T-box contains a long stem region after Stem I, which could account for a Stem II domain, a feature that has not been previously described in glyQ T-boxes. Stem II, together with the Stem II A/B pseudoknot, is absent for the remaining T-boxes under study, as is true for the typical Bacilli glyQS T-boxes. Instead, they contain an interdomain linker with variable length, which is unusually short in the S. pneumoniae glyQS T-box. The Stem III domain is present but with lower conservation. This is expected, as only the termini of the Stem III contribute to the discriminator function, which permits the length and sequence of the stem–loop to vary without losing the T-box function. Stem III in the C. tetani glyS T-box appears shorter compared to the two streptococcal T-boxes. Finally, the terminator/antiterminator stems of typical lengths are found to flank the highly conserved T-box bulge 5′ UGGAACC 3′ (nucleotides 205–211 in the alignment) (Fig. 2), followed by a highly conserved downstream region located in the 3′ end of the antiterminator stem. Altogether, the comparison of the corresponding T-boxes verified the conservation of the important structural features, but also structural peculiarities that could affect their ability to sense their tRNA ligands and regulate transcription.
L. monocytogenes and C. tetani contain typical Bacilli-like glycyl T-boxes
Primer extension analysis using specific primers revealed that the secondary structure of the glyQS T-box from L. monocytogenes is similar to those from Bacilli which are considered typical structures (Fig. 3; Supplemental Table S1). The Stem I region of the L. monocytogenes glyQS T-box contains all the conserved structural features and nucleotides, with a typical K-turn (GA motif). The analysis showed that K-turn is extensively modified by both dimethyl-sulfate (DMS) and kethoxal (KE) at positions G18, A22, and A23 at the 5′ side and G107 and G108 at the 3′ side (Fig. 3). The conventional K-turn consists of a 3 nt bulge flanked on its 3′ side by base pairs G•A and A•G (Noncanonical Stem, N-C Stem), and occasionally one or two non-Watson–Crick pairs before regular base-pairing continues, and on its 5′ side by a segment of normal base-pairing (Canonical Stem, C Stem) (Lilley 2014). In the L. monocytogenes T-box, the K-turn consists of the 3 nt bulge formed by G18, U19, and U20. The 3′ side of the bulge is formed by the G21•A109 and A22•G108 and the non-Watson–Crick G24•U106 base pairs (Fig. 3B). The specifier loop also contains the conserved nucleotides described previously and is 7 nt long. Modifications by DMS and KE were observed at positions A31, G32, and A34 and G96, A98, the codon-like triplet G99, G100, C101, and the conserved G102 (Fig. 3). A C-loop preceding the specifier loop can be discerned in positions C43, C45, and A87, and also contains a bulged A87 responsible for a subtle bend in the Stem I that positions the codon-like triplet and the IDTM in the optimal orientation for tRNA interaction (Grigg et al. 2013; Li et al. 2019).
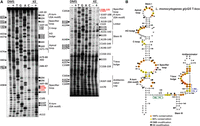
Chemical probing analysis of glyQS T-box from L. monocytogenes. (A) Α 6% PAGE/8 M urea denaturing gel showing primer extension analysis of modified with DMS or KE (+) and unmodified (−) L. monocytogenes glyQS T-box transcript (20 pmol). The triangles on the left correspond to DMS modifications, whereas the hexagons on the right identify KE modifications, according to the sequencing tracks (lanes T, G, A, and C). (B) The predicted secondary structure of L. monocytogenes glyQS T-box. LiMo_PE_3 primer (in green) was used for Stem I primer extension analysis and LiMo_PE_4 (in blue) for the analysis of the antiterminator domain. Highly conserved (100%) and moderately conserved (66%) nucleotides are indicated with orange and yellow circles, respectively. The specifier codon is indicated with red letters. The trinucleotide at the T-box bulge that interacts with the 3′ CCA of the tRNA is indicated with red circles.
The probing analysis verified the existence of an AG bulge which contains a 10 nt loop compared to the 8 nt loop that is found in the Bacilli glyQS T-boxes, as judged by the modification pattern of A53, G54, G56, A57, and A82. An interesting difference is the existence of five consecutive U (48–52) right before the conserved nucleotides.
The Apical loop is 12 nt long and includes the conserved G68, U70, G71, and A74 and the moderately conserved G69 and G73 nucleotides; however, only G64, G69, and A74 appeared accessible for modification. This observation verifies the reported long-range interactions between the Apical loop and the AG bulge (Grigg et al. 2013).
A 14 nt long single-stranded linker region that connects Stem I with the antiterminator region is purine-rich, and among them, A119, A120, A124, G126, and G128 appeared modified. In addition, contains the Stem III domain which is important for the formation of the discriminator conformation. It consists of a 6 bp stem and a loop of 4 nt, among which G135 was modified and followed by the conserved A145, A146, and the nonconserved A147. The probing analysis also showed that the antiterminator domain does not deviate from the canonical pattern that is observed in a typical T-box and verified that it contains all the conserved nucleotides that have been reported, including the conserved T-box bulge 5′-UGGAACC-3′ sequence. This region also contains the G-U wobble pair (G160-U175) at the top of the T-box bulge, which has been shown to be important for the development of the steric filter that examines the aminoacylation status of the associated tRNA. Additionally, at the top of the antiterminator domain, a 6 nt loop is formed, in which C165, A167, A168, and C169 were found modified, confirming the existence of a solvent exposed, flexible loop in this position. Overall, the glyQS T-box from L. monocytogenes contains all the structural features and nucleotides that are found in a typical T-box riboswitch.
The same analysis was performed for the glyS T-box from C. tetani to verify the predicted secondary structure (Fig. 4; Supplemental Table S1). All the conserved structural elements and nucleotide sequences that are present in a typical T-box riboswitch are observed in the Stem I region of the C. tetani glyS T-box. A typical K-turn (or GA motif) exists and the positions G18, A19, G21, A22, G105, and A106 of the K-turn bulge were modified (Fig. 4A). However, although both T-boxes appear very similar in sequence between them and compared to the Bacilli counterparts, few but important differences exist. The first difference that was observed is the absence of a C-loop, replaced by UxU mismatches. The second difference is in the Stem III domain, which is shorter than those found in the Bacilli glyQS T-boxes. It contains a stem comprised of 3 bp and a loop that is formed by 4 nt (Fig. 4B). The modification of G129 by KE validates the formation of a loop at this position. Collectively, these results suggest that the L. monocytogenes glyQS and the C. tetani glyS T-boxes although very similar between them and compared to the Bacilli T-boxes exhibit differences which serve the sensing mechanism and could be exploited further for targeting by antibacterial compounds.
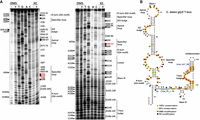
Chemical probing analysis of glyS T-box from C. tetani. (A) Α 6% PAGE/8 M urea denaturing gel showing primer extension analysis of modified with DMS or KE (+) and unmodified (−) C. tetani glyS T-box transcript (20 pmol). The triangles on the left correspond to DMS modifications, whereas the hexagons on the right identify KE modifications, according to the sequencing tracks (lanes T, G, A, and C). (B) The predicted secondary structure of C. tetani glyS T-box. Clote_PE_3 primer (in green) was used for Stem I primer extension analysis and Clote_GLT_RS (in blue) for the analysis of almost the whole T-box region. Highly conserved (100%) and moderately conserved (66%) nucleotides are indicated with orange and yellow circles, respectively. The specifier codon is indicated with red letters. The trinucleotide at the T-box bulge that interacts with the 3′ CCA of the tRNA is indicated with red circles.
S. pneumoniae and S. pyogenes glyQS T-boxes deviate significantly from the typical T-box structural context
Primer extension analysis of the streptococcal T-boxes revealed that although they contain the important conserved structural features, some unusual deviations from a typical T-box riboswitch occur. Specifically, at the base of Stem I of the S. pneumoniae glyQS T-box, a typical GA motif could not be detected, denoting the absence of a typical K-turn. The only modified nucleotide that was observed in this region is G22, which is presented in a short bulge in the secondary structure (Fig. 5). This observation is in line with a previous in silico analysis of 200 putative T-boxes from different microorganisms showing that a recognizable GA motif was rare among streptococci (Winkler et al. 2001). The specifier loop at its 5′ side is short and contains five conserved nucleotides, among which only A28 and A29 appear modified by DMS. The main difference with the typical T-box is that the highly conserved 5′ AGUA 3′ sequence is replaced by 5′ AGUC 3′. The specifier loop at its 3′ side is 10 nt long and lacks the conserved 5′ GAA 3′ sequence, and most of the nucleotides in this loop appeared modified, among which are G95, G96 that belong to the specifier codon and G98 (Fig. 5). Right above the specifier loop, a symmetrical internal loop is formed, distinct from the C-loop that is usually found in this position. The formation of the internal loop is evidenced by its solvent exposure and frequent modifications (C40, A82, A83, and A84) (Fig. 5B). This loop could possibly contribute to the necessary bend required for tRNA elbow interaction with the T-box AG bulge and Apical loop, with both features exhibiting the expected length, sequence, and conservation. Notably, the single-stranded interdomain linker that connects Stem I with the antiterminator is surprisingly short in the S. pneumoniae glyQS T-box, only 5 nt long, and exhibits very low susceptibility to modifications (only G108). The shorter linker may be compensated by the elongated stem below the specifier loop. The Stem III domain appears typical Bacilli-like, while the Stem III 3′ flanking region contains three additional purines, of which A134 and A135 appeared modified by DMS (Fig. 5B). Unlike the rather nontypical linker and Stem III flanking regions, a typical antiterminator domain follows that contains the highly conserved T-box bulge. The G-U wobble pair that normally exists at the top of the T-box bulge is substituted by a G-C pair at positions G149-C161. It was shown previously that a G-C pair can also serve as a steric filter to sense the aminoacylation status of the cognate tRNA but is less frequently observed than the G-U wobble pair (Li et al. 2019). At the top of the antiterminator there is a loop consisting of 3 nt (Fig. 5B).
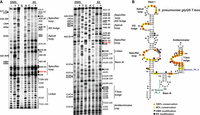
Chemical probing analysis of glyQS T-box from S. pneumoniae. (A) Α 6% PAGE/8 M urea denaturing gel showing primer extension analysis of modified with DMS or KE (+) and unmodified (−) S. pneumoniae glyQS T-box transcript (20 pmol). The triangles on the left correspond to DMS modifications, whereas the hexagons on the right identify KE modifications, according to the sequencing tracks (lanes T, G, A, and C). (B) Predicted secondary structure of S. pneumoniae glyQS T-box. Spneum_PE_3 primer (in green) was used for Stem I primer extension analysis and Spneum_PE_5 (in blue) for the analysis of the whole T-box region. Highly conserved (100%) and moderately conserved (66%) nucleotides are indicated with orange and yellow circles, respectively. The specifier codon is indicated with red letters. The trinucleotide at the T-box bulge that interacts with the 3′ CCA of the tRNA is indicated with red circles.
Analysis of the secondary structure of the glyQS T-box from S. pyogenes revealed that this T-box also exhibits structural differences compared to a typical glyQ T-box. In the Stem I region of this T-box, the typical GA motif that denotes a K-turn is missing. However, a pyrimidine-rich internal loop replaces the K-turn, may fold into a “C-loop” motif (Lescoute et al. 2005) as seen in the G. kaustophilus glyQ Stem I, and could potentially account for the necessary backbone bend that facilitates interaction with the cognate tRNA (Fig. 6B). The specifier loop follows the typical pattern, except for the highly conserved 5′ AGUA 3′ sequence that is substituted by the 5′ AGGA 3′. The Stem I structure contains two small bulges in which only A34 at the first bulge appears modified by DMS, that most likely are separated by a G35-C83 pair. However, as C83 appears modified by DMS (Fig. 6A), it is possible that this base pair is not stable enough, and instead of two bulges, a loop could be formed in the tertiary structure. The UA dinucleotide bulge could serve a similar structural role as the AU dinucleotide in Oceanobacillus iheyensis glyQ Stem I. In either case, this region seems to bend the helical axis of the RNA to better track the L shape of the tRNA substrate. The AG bulge and the Apical loop of the S. pyogenes glyQS T-box present a typical nucleotide sequence (Fig. 6). Surprisingly, this T-box contains a highly unusual Stem II domain that is 36 nt long with a small bulge, an internal loop, and a distal loop at the bottom. Stem II domains are generally absent in glycyl T-boxes, and this is the first reported T-box that contains such a strikingly distinct feature. The Stem II region is not susceptible to modifications by either DMS or KE, as only A124 that is positioned in the distal loop appeared modified by DMS. This suggests the formation of Stem II and the protection of the nucleotides from modification. An additional confirmation of the Stem II formation is a major stop of the reverse transcription reaction which is observed right after C140 (Fig. 6). This nucleotide participates in the first base pair of the stem, and the folding of this region explains the substantial drop-off of the reverse transcriptase at this location. However, although all Stem II domains that have been characterized so far contain an S-turn motif, in the case of S. pyogenes glyQS T-box, there is no immediately recognizable S-turn. The presence of an unusual Stem II domain also accounts for a shorter interdomain linker, which is 7 nt long and is composed only of purines (six adenosines and only one guanosine), which are not accessible for modification by either DMS or KE. Curiously, there is no immediately recognizable Stem IIA/B pseudoknot here, which may suggest a Stem II structure and function that is distinct from the typical Stem II domain (Battaglia et al. 2019; Suddala and Zhang 2019a). An unconventional Stem III with a larger loop that consists of 4 bp and a loop of 10 nt is also detected, which, however, preserves the conserved flanking purines at both sides of Stem III (Fig. 6). Finally, a typical antiterminator domain with the highly conserved T-box bulge and a G-C instead of the G-U wobble pair is found (Fig. 6).
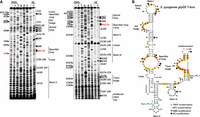
Chemical probing analysis of glyQS T-box from S. pyogenes. (A) Α 6% PAGE/8 M urea denaturing gel showing primer extension analysis of modified with DMS or KE (+) and unmodified (−) S. pyogenes glyQS T-box transcript (20 pmol). The triangles on the left correspond to DMS modifications, whereas the hexagons on the right identify KE modifications, according to the sequencing tracks (lanes T, G, A, and C). (B) The predicted secondary structure of S. pyogenes glyQS T-box. Spyo_PE_3 primer (in green) was used for Stem I primer extension analysis and Spyo_PE_4 (in blue) for the analysis of the whole T-box region. Highly conserved (100%) and moderately conserved (66%) nucleotides are indicated with orange and yellow circles, respectively. The specifier codon is indicated with red letters. The trinucleotide at the T-box bulge that interacts with the 3′ CCA of the tRNA is indicated with red circles.
The Stem I domain of all T-boxes can form a complex with the tRNAGlyGCC ligand
The formation of a Stem I:tRNAGly complex for each T-box was examined using electrophoretic mobility shift assays (EMSAs). The binding properties of the L. monocytogenes glyQS T-box and C. tetani glyS T-box, both of which feature a typical Stem I domain, show slightly different binding affinities, as evidenced by their respective Kd values. The L. monocytogenes glyQS T-box demonstrates a Kd of 2.858 ± 0.433 μM (Fig. 7A), indicating a relatively strong and specific interaction between its Stem I domain and the tRNAGly ligand. On the other hand, the C. tetani glyS T-box exhibits a slightly higher Kd value of 3.425 ± 0.537 μM, which suggests a slightly weaker binding affinity compared to the L. monocytogenes glyQS T-box (Fig. 7B). Moreover, the Stem I domain from the S. pneumoniae glyQS T-box also has a comparable binding affinity, with a Kd value of 3.545 ± 0.807 mM (Fig. 7C), falling within the same range like the other two Stem I domains. In contrast, the Stem I domain of the S. pyogenes glyQS T-box displays the highest observed Kd value (4.424 ± 0.829 mM; Fig. 7D), indicating a relatively weaker binding affinity for the same tRNAGly.
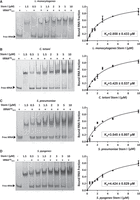
EMSAs between increasing concentrations of (A) L. monocytogenes Stem I, (B) C. tetani Stem I, (C) S. pneumoniae Stem I, and (D) S. pyogenes Stem I domains from the respective glycyl T-boxes and 2 μM of tRNAGlyGCC. At the left, the Native PAGE images are shown, while at the right the binding curve of the bound tRNAGlyGCC against each Stem I is indicated. Also, the estimated Kd values for each complex are shown.
Overall, these results highlight that despite the structural variations in Stem I domains, they can all successfully form complexes with the tRNA, emphasizing their functional versatility and adaptability in the recognition and binding of their target molecules.
The interaction pattern with tRNA is conserved among all four glycyl T-boxes
To identify possible differences in the interaction between T-boxes and the tRNA ligand, which can be associated with the structural idiosyncrasies among these T-boxes, we performed chemical probing analyses in the presence of increasing concentrations of the B. subtilis tRNAGlyGCC. The protection sites that are induced by the tRNAGlyGCC ligand could be caused either directly by the binding of the tRNA on the T-box, or indirectly when the interaction between the tRNA and the T-box leads to a different conformation of a region or of specific nucleotides that become inaccessible to modifications by DMS or KE.
In the L. monocytogenes glyQS T-box, the nucleotide A23 of the K-turn, positions A53 and A57 of the AG bulge and G68 and G69 of the Apical loop, and position A87 of the 3′ side of the C-loop were found protected by the tRNA. The 3′ side of the specifier loop was observed extensively protected by the tRNA at positions A97, A98, G99, G100, and C101 (Supplemental Fig. S1). The latter 3 nt compose the codon-like triplet that interacts with the tRNA anticodon, while immediately adjacent to the A97 and A98 are part of the S-turn (or Loop E) motif that contributes to the correct presentation of the specifier codon for interaction with the tRNA. Analysis of the linker and the Stem III domains revealed that in the presence of the tRNA, the positions A118, A119, and A120 of the linker and A146 and A147 of the Stem III were protected, like most of the T-box bulge nucleotides (Supplemental Fig. S1). Altogether, the L. monocytogenes glyQS T-box and the tRNA exhibit a pattern of interaction and protection that is highly congruent with the recent cocrystal and cryo-EM structures of the T-box–tRNA complexes. The same interactions were also observed for the C. tetani glyS T-box:tRNA complex. In this case, except for the G96, G97 and C98 specifier codon, the positions G92 and G99 appeared protected as well by the tRNAGlyGCC ligand, which is consistent with local structural changes caused by the binding of the tRNA anticodon (Supplemental Fig. S2). Analysis of the linker and Stem III domains revealed that the presence of the tRNA induced the protection of the sites G116 and G124. The T-box bulge was also observed protected by tRNA at G144 and G145 that belong to the 5′ UGG 3′ trinucleotide that interacts with tRNA 5′ CCA 3′ end, for sensing its aminoacylation status.
In the case of the S. pneumoniae glyQS T-box, tRNA induced the protection of A48, A49, and G52 of the AG bulge and A63 and G64 of the Apical loop (Supplemental Fig. S3). In the specifier loop of this T-box, all the nucleotides of the specifier codon G95, G96, and C97 were found protected, similar to what has been described for the other T-boxes. Interestingly, tRNA binding also caused strong protection at G22, which may suggest a conformational change in this region that serves to redirect the T-box trajectory in place of a K-turn. In the linker and Stem III regions, no nucleotides were found protected except for A131, which flanks the Stem III. Finally, the T-box bulge positions G143 and G144 were also found protected, as expected, along with A145. As the flanking purines of Stem III participate in the T-box discriminator conformation, the protection of A131 could be a result of this interaction (Supplemental Fig. S3).
The tRNA-induced protections of A47, A48, and A52 of the AG bulge and A62, A66, and A68 of the Apical loop of the S. pyogenes glyQS T-box were similar to other T-boxes (Supplemental Fig. S4). In the specifier loop, positions A89, G92, G93, and C94 were also protected, as expected. Of note, the apparent Stem II domain that exists in this T-box does not seem to interact with the tRNA, as there was no discernible tRNA protection of this region. Taken together, the probing analysis indicates that these glycyl T-boxes from pathogens exhibit similar interaction patterns with the tRNA ligand, and that the sequence and structural diversification of the T-box, while maintaining the conserved core features, retained the tRNA-binding functions.
T-box structural idiosyncrasies do not affect in vivo response to glycine starvation
Using a previously described in vivo antitermination assay, we tested all four glycyl T-boxes for response to glycine starvation in a heterologous environment using appropriate Escherichia coli strains (Stamatopoulou et al. 2017; Giarimoglou et al. 2022b). All T-boxes could induce transcription of the dTomato gene in response to glycine deprivation, presumably due to uncharged tRNAGly binding (Fig. 8). The greatest transcription induction ratio was observed for the L. monocytogenes glyQS T-box (Supplemental Fig. S4). The streptococcal glycyl T-boxes exhibited similar transcription induction levels compared to the C. tetani glyS T-box (Supplemental Fig. S4). These results are normalized by the transformed strain that contains only the plasmid that includes each glycyl T-box tested, which is used as a control of possible nonspecific transcription induction caused by the endogenous E. coli tRNAGly isoacceptors. Interestingly, the dTomato fluorescence signal was lower than in the strains carrying a plasmid expressing the tRNAGlyGCC ligand (Supplemental Fig. S5). The in vivo functional data corroborate the chemical probing analyses of the T-boxes in the absence and presence of tRNA. Taken together, our data indicate that despite substantial structural divergences, these T-boxes are functionally competent in binding cognate tRNAs, sensing their aminoacylation status using a limited set of conserved recognition sites, and the induction of the necessary conformational changes that render the appropriate genetic decisions.
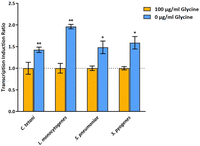
Transcription induction ratio of dTomato gene of C. tetani, L. monocytogenes, S. pneumoniae, and S. pyogenes glyQS T-boxes under glycine-replete and glycine-starvation conditions, normalized to each T-box-containing strain grown in minimal media supplemented with glycine (orange bars). The values and error bars represent mean and SD, n = 3 biologically independent samples. Significance stars represent ns: P > 0.05, (*) P ≤ 0.05, (**) P ≤ 0.01, (***) P ≤ 0.001.
DISCUSSION
T-box riboswitches display a wide range of sequence and structural variations among various bacterial genera and species. When examining T-boxes across a broader range of organisms and considering their regulatory functions within pathogens, a remarkable variability in terms of length and unique structural patterns is observed. It is important to note that the apparent variability cannot be directly attributed to a coevolutionary relationship with their cognate tRNA molecules. Instead, the structural idiosyncrasies observed in T-boxes seem to align with the specific modulatory needs which depend on the expression of associated tRNA molecules in different organisms and under different environmental conditions (Green et al. 2010; Saad et al. 2013; Wencker et al. 2021).
T-box riboswitch comparative studies have led to better understanding of how bacterial RNA-dependent regulatory systems modulate adaptation to diverse environmental niches. To deepen current knowledge on T-boxes and to identify potential variations regarding their structural features, the corresponding glycyl T-boxes from four prominent human pathogens (C. tetani, L. monocytogenes, S. pneumoniae, and S. pyogenes) were examined in the present study, with existing information being relatively limited (Wilton et al. 2015; Lebreton and Cossart 2017; Eichner et al. 2022; Pavlova and Penchovsky 2022).
In silico and chemical probing analysis of the secondary structure of each glycyl T-box revealed that L. monocytogenes glyQS and C. tetani glyS T-boxes contain all the characteristic structural features of a T-box riboswitch, and their secondary structures do not deviate significantly from those of glycyl T-boxes from Bacilli, in contrast to glyQS T-boxes from S. pneumoniae and S. pyogenes that exhibit several structural differences. More specifically, all four T-boxes exhibited a characteristic Stem I which contains conserved nucleotides in the 5′ and 3′ sides of the specifier loop, AG bulge, and Apical loop. However, some variations were observed, such as a different motif in the 5′ side of the specifier loop in the streptococcal T-boxes. All T-boxes contained a conserved specifier codon and an adjacent purine, which contribute to the intermolecular helix formation. Notably, the C. tetani glyS T-box contained the 5′ GGA 3′ specifier codon, in contrast to the other glycyl T-boxes containing the 5′ GGC 3′ specifier codon.
To our surprise, the analysis showed that a K-turn motif could not be detected and verified in the streptococcal glyQS T-boxes. So far, K-turns were considered highly conserved structural motifs found not only in T-box riboswitches, but also in ribosomal RNA and other riboswitches involved in small molecule binding (Huang and Lilley 2016). In the S. pyogenes glyQS T-box, a loop is formed at the location where a K-turn is typically observed, which contains the characteristics of a C-loop motif. This loop could potentially play a role in facilitating the required bend for efficient tRNA interaction, in place of a K-turn. Experimental evidence has previously demonstrated that mutations disrupting the K-turn motif lead to the loss of tRNA-dependent transcription antitermination in vivo (Winkler et al. 2001), and deletion of this motif has been observed to have minimal impact on the Kd for tRNA binding to Stem I (Zhang and Ferré-D'Amaré 2013). Many bacteria, particularly Gram-positive species, express K-turn-binding proteins with high affinities, potentially aiding in maintaining their kinked conformation (Baird et al. 2012; Huang and Lilley 2013). However, certain T-box riboswitches, such as the glyQS riboswitch, have been shown to function efficiently in vitro even in the absence of K-turn-binding proteins, which raises questions regarding the general significance of these proteins for T-box functionality in vivo. A previous in silico analysis of 200 putative T-boxes derived from various microorganisms showed a surprisingly high percentage (29%) that lack a recognizable GA motif. Instead, these T-boxes contain an extended helix without an internal loop. Interestingly, no significant correlation was found between the presence of the GA motif and the amino acid classes represented. However, a trend was observed regarding the occurrence of this motif in different organisms. Strikingly, a high representation of this motif (94%) was observed in organisms belonging to the Bacillus–Clostridium group, suggesting its prevalence in this taxonomic group. In contrast, the GA motif was found to be extremely rare in leaders from Staphylococcus and Streptococcus, with a mere 2% occurrence (Winkler et al. 2001). Indeed, the S. aureus glyS T-box was also previously reported to contain a noncanonical K-turn, while the S. aureus ileS T-box was proposed to function without a K-turn (Grundy et al. 1997; Apostolidi et al. 2015). These observations further support the presumed absence of a K-turn in Streptococcal T-boxes examined in the present study, although the existence of an alternative strategy cannot be excluded.
Equally interesting was the finding of a Stem II domain in the S. pyogenes glyQS T-box, and although in other T-boxes the Stem II is immediately followed by a Stem IIA/B pseudoknot, in this case, the latter was not detected (Battaglia et al. 2019; Suddala and Zhang 2019a). All the previously characterized glycyl T-boxes lack a Stem II domain, an absence which does not impair their regulatory function. Glycyl T-boxes efficiently bind their cognate tRNAGly by pairing the GGC codon of the T-box with the GCC anticodon, resulting in the formation of a highly stable 3 bp duplex. Additionally, the heteroduplex is structurally reinforced by the presence of tR37 above and another purine below, which together sandwich the codon–anticodon duplex, enhancing its axial stability. The robust codon–anticodon interactions provided by this pairing likely eliminate the necessity for additional lateral stabilization from the Stem II domain. Moreover, it has been proposed that in Stem II-containing T-boxes, the K-turn is believed to be essential for accurate positioning of Stem II, as Stem II itself lacks the necessary sequence complementarity to independently locate its docking site on Stem I (Zhang 2020; Suddala et al. 2023). This does not apply for the S. pyogenes glyQS T-box, where the presence of the Stem II domain is not accompanied by the presence of a K-turn. However, the loop that we detected at the base of Stem I could possibly substitute the role of the K-turn.
A linker region of variable length and a Stem III domain were present in all T-boxes, with low conservation. Additionally, all T-boxes displayed a typical terminator/antiterminator stem structure. Furthermore, the Stem I domain of all four T-boxes showed a similar ability to form a complex with the tRNAGlyGCC ligand, albeit with differential affinities that can be attributed to the observed structural variations. This observation underscores the effectiveness and adaptability of this molecular architecture in precisely and efficiently recognizing tRNAs, and highlights the independent selection of this structural motif multiple times throughout evolution and its functional importance across diverse biological contexts. These findings provide supporting evidence for the evolutionary adaptation of different Stem I domains to accommodate and interact with their specific ligands, showcasing the remarkable ability of biological systems to adapt and thrive in different environments. Despite these structural variations, all glycyl T-boxes were capable of inducing transcription in response to glycine deprivation and uncharged tRNAGly binding, underscoring their functional robustness and adaptability.
One more intriguing observation is the clear presence of a Stem II domain in the S. pyogenes glyQS T-box and the absence of the K-turn from the streptococcal glyQS T-boxes, all of which are able to capture their tRNAGly and to form a complex and to function in vivo. This could possibly represent an adaptation of the S. pyogenes glyQS T-box to accommodate additional regulatory elements (such as RNA-binding proteins other than L7Ae/YbxF) or to optimize its interaction with the cognate tRNA. These structural variations in different glycyl T-boxes may reflect the diverse genetic backgrounds and environmental niches in which these organisms reside, allowing them to fine-tune their regulatory mechanisms accordingly. Such adaptations are essential for ensuring optimal gene expression control and cellular response to varying conditions, underscoring the remarkable versatility and adaptability of these regulatory elements in maintaining cellular homeostasis.
To further elucidate the structural and functional variations among T-box riboswitches, several experimental approaches could be used. For example, performing electrophoretic mobility shift assay (EMSA) or isothermal titration calorimetry (ITC) on T-boxes that deviate from the typical T-box pattern like those derived from S. pneumoniae, S. pyogenes, or other streptococcal species, will help discern the impact of the absence of the K-turn motif on tRNA binding affinity. These assays will reveal whether the absence of the K-turn motif in streptococcal T-boxes may result in altered binding affinities, or is potentially compensated for by other adjacent structural elements, such as the C-loop motif observed in Bacilli T-boxes. Additionally, by using site-directed mutagenesis, the functional roles of specific nucleotides within crucial regions of these T-box riboswitches can be evaluated. By strategically introducing mutations targeting key residues implicated in essential structural elements beyond Stem I, such as the stabilizing “latch,” long-range interactions within the discriminator, the G167-U185 wobble pair, and Stem III contacts, one is expected to quantify their precise contributions to T-box structural integrity and function, and compare with the previously characterized Bacilli T-boxes (Li et al. 2019). Finally, mutation experiments in these key regions will test the proposed secondary structures and provide further insights into the adaptive evolution and comparative functionality of these riboswitches. Collectively, these approaches will deepen our understanding of the evolutionary adaptations in T-box riboswitches and their role in shaping gene expression across diverse bacterial genera and could pave the way for advancements in the development of RNA-targeting therapeutics.
MATERIALS AND METHODS
Chemicals, enzymes, and plasmid vectors
The primers used for primer extension analysis, cloning, in vivo antitermination assays, and in vitro transcription of T-boxes and tRNAGly were synthesized by Eurofins Scientific (Supplemental Table S1). Restriction enzymes, DNase I, T4 DNA ligase, T7 RNA polymerase, inorganic pyrophosphatase, RNase inhibitor, and M-MuLV Reverse Transcriptase were purchased from New England Biolabs. High-fidelity DNA polymerase was purchased from Kapa Biosystems. DMS and KE reagents used for chemical modification experiments were purchased from Sigma-Aldrich. The Q5 Site-Directed Mutagenesis Kit that was used was purchased from New England Biolabs. All chemicals and antibiotics were purchased from Sigma-Aldrich or PanReac AppliChem. Plasmid DNA extraction was conducted using the NucleoSpin Plasmid Mini Kit, and PCR products were purified using the NucleoSpin Gel and PCR Clean-up Kit from Macherey-Nagel. [γ-32P] ATP (6000 Ci/mmol) was purchased from Hartmann Analytic. pXS-dTomato was a gift from Karen Guillemin (Addgene plasmid 117387; http://n2t.net/addgene:117387; RRID: Addgene 117387) (Wiles et al. 2018).
Sequence alignment
The sequences of the glyQS T-boxes from C. tetani, L. monocytogenes, S. pneumoniae, and S. pyogenes, which were used for the multiple sequence alignment, were identified using the RegPrecise database (Novichkov et al. 2013). The broader full-length T-box riboswitch region (containing the specifier loop's sequence) was validated further using the KEGG and TBDB (T-box Riboswitch Annotation Database) databases (Aoki and Kanehisa 2005; Marchand et al. 2021). The multiple sequence alignment was performed utilizing the Clustal Omega sequence alignment tool and was illustrated using Jalview 2.11.1.7 (Waterhouse et al. 2009; Goujon et al. 2010). Moreover, the tRNAGly isoacceptor sequences from different bacterial organisms, which were used in a different multiple sequence alignment, were retrieved using the tRNAscan-SE 2.0 tool and TBDB database (Lowe and Chan 2016; Chan and Lowe 2019; Marchand et al. 2021).
Site-specific mutagenesis of the C. tetani glyS T-box specifier codon
The point mutation A98C was performed in the C. tetani glyS T-box to change the specifier codon from 5′ GGA 3′ to 5′ GGC 3′, to achieve full complementarity with the anticodon of B. subtilis tRNAGlyGCC. For this purpose, the Q5 Site-Directed Mutagenesis Kit (NEB) was used and specific primers were designed (Supplemental Table S1). The pXS-dTomato plasmid vector carrying the C. tetani glyS T-box riboswitch was used as a DNA substrate and a PCR was performed according to manufacturer's instructions. Subsequently, a second reaction was performed with the KLD mix that contains the enzymes kinase, ligase, and DpnI. This reaction was also performed following the manufacturer's protocol. The reaction was incubated at room temperature for 15 min followed by transformation in E. coli DH5α chemically competent cells. The next day, a single colony was used to inoculate the LB medium, and the culture was incubated overnight at 37°C under 200 rpm shaking. After the incubation, isolation of the plasmid was performed and the plasmid was sequenced to verify the integrity of the cloned sequence.
In vivo antitermination assays
C. tetani, L. monocytogenes, S. pneumoniae, and S. pyogenes glyQS T-boxes were placed under the control of the constitutive promoter vegII and before the dTomato gene (derivative of orange fluorescent protein) in pXS-dTomato plasmid vector, as previously described (Wiles et al. 2018; Giarimoglou et al. 2022b). The B. subtilis tRNAGlyGCC were previously cloned in the pBAD-18-Kan vector, as previously described (Stamatopoulou et al. 2017). The E. coli strain M5154 was chemically transformed using both recombinant plasmids (Saad et al. 2013). The single- and double-transformed strains were subsequently used to monitor the fluorescence emission of the dTomato protein produced in each condition. In the cells cultured in minimal growth conditions where glycine is absent, the uncharged tRNAGly binds on the T-box inducing the antitermination conformation, allowing the RNA polymerase to continue transcribing the downstream gene, i.e., dTomato. Overnight, cultures in minimal medium (1× M9 salts, 2 mM MgSO4, 0.1 mM CaCl2, 0.2% glycerol supplemented with 25 μg/mL l-tryptophan) were used to inoculate cultures, starting from an OD595 of 0.1 in minimal media under starvation (0 μg/mL) and nonstarvation conditions (100 μg/mL glycine) for 4 h. 0.1% l-Arabinose was added in all the cultures to induce the expression of tRNAGly. After 4 h of incubation, each culture was measured at OD595 to determine bacterial growth, and the fluorescence of dTomato was measured in FLUOstar Optima with excitation at 550 nm and emission at 590 nm. The fluorescence values of dTomato for each culture were divided by using the respective OD595 values, and the resulting values for each condition were further normalized to that of the wild-type T-box-containing strain grown in minimal medium supplemented with glycine (data not shown). For each condition examined, the effect of endogenous tRNAs on nonspecific interaction with wild-type and mutated T-boxes was subtracted. The values and error bars represent mean and standard deviation (SD); n = 3 biologically independent samples.
RNA preparation of glycyl T-boxes and B. subtilis tRNAGly
The RNA of the whole region or Stem I domain of all glycyl T-boxes under investigation as well as B. subtilis tRNAGlyGGC was performed as previously described with minor modifications (Giarimoglou et al. 2022b). All T-boxes were PCR amplified with high-fidelity DNA polymerase, introducing a T7 promoter leader sequence. The PCR products were subsequently used as templates for in vitro transcription using T7 RNA polymerase. Run-off in vitro transcription reactions were carried out at 30°C for the T-boxes and at 37°C for the B. subtilis tRNAGlyGCC for 16 h in the presence of inorganic pyrophosphatase (8 U). Following DNase I treatment and extraction with phenol:chloroform:isoamyl alcohol (25:24:1), the transcripts were purified using an 8% PAGE/8 M urea gel. The bands corresponding to the T-boxes and B. subtilis tRNAGlyGCC were excised after visualization under a UV lamp, and they were subsequently eluted in RNA elution buffer, comprising 20 mM Tris–HCl (pH 7.5), 250 mM sodium acetate, 1 mM EDTA (pH 8), 0.25% SDS, and 1% phenol.
Chemical probing of all glycyl T-boxes in the presence or absence of tRNAGly
The chemical modification reactions of the T-boxes were performed using DMS and KE as previously described (Apostolidi et al. 2015; Giarimoglou et al. 2022b). Twenty picomoles of the transcripts of the glycyl T-boxes, alone or in combination with a transcript of the B. subtilis tRNAGlyGCC molecule (200 and 400 pmol), were added to 1x modification buffer [70 mM HEPES-KOH, pH 7.8, 10 mM Mg(OAc)2, and 270 mM KOAc] and incubated at 60°C for 10 min for denaturation. Then, they were slow cooled in a water bath starting from 60°C, until the temperature reached 25°C, in order to obtain the correct conformations. Reactions were incubated at 25°C for 30 min and then placed on ice for 10 min. After 10 min, DTT (1 mM) was added to each reaction. Then the modification with DMS or KE followed. Either 1 µL DMS (diluted 1:1 with 100% ethanol), or 1 µL KE (diluted 1:1 with DEPC ddH2O) was added to each reaction and the reactions were incubated at 30°C for 30 min. After the incubation, each reaction was stopped with the respective Stop Buffer for DMS (0.25 M Tris-acetate [pH 7.5], 0.25 M β-mercaptoethanol, 0.3 M sodium acetate [pH 9.2], and 0.025 mM EDTA) and KE (0.3 M sodium acetate [pH 6] and 25 mM potassium borate [pH 6]) and phenol extraction and ethanol precipitation followed. In the primer extension reactions, 1 pmol of each modified transcript was hybridized with the corresponding 5′-[γ-32P]-primer in 1× hybridization buffer (2 mM Tris-acetate, pH 7.9, 0.25 M KOAc, 0.2 mM EDTA) in a final volume of 10 μL. The reactions were incubated at 80°C for 10 min, placed in a water bath at the same temperature to slow cool until the temperature slowly dropped to 47°C. Subsequently, the reactions were induced by adding 1× primer extension buffer (23 µL) [20 mM Tris-acetate pH 8.3, 10 mM Mg(OAc)2, 5 mM DTT, 1 mM of each dNTP] in the presence of 2U RNasin and 5U M-MuLV reverse transcriptase in a final volume of 42 µL and incubated at 47°C for 1 h. After adding 158 µL of 1× TE buffer (10 mM Tris-acetate pH 7.5, 0.1 mM EDTA), the produced cDNAs were extracted in an equal volume of phenol/chloroform/IAA (25:24:1) and precipitated with ethanol. Resuspension was performed in 4 µL 0.1 N NaOH and 8 µL f-EDTA loading buffer and the cDNAs were denatured at 80°C for 2 min before running on a 6% PAGE/8 M urea denaturing gel. Finally, five additional primer extension reactions were performed in each experiment using unmodified T-box transcripts. The first was used as a negative control of the reaction while the remaining four were used as sequencing reactions, in the presence of 0.17 mM of each ddNTP (ddTTP, ddGTP, ddCTP, ddATP). Finally, the results of primer extension analysis were visualized using phosphoimager (Fuji lm FLA 3000 platform) and were quantified using the AIDA image analyzer software (Raytest version 5.0).
Electrophoretic mobility shift assays
EMSAs were performed between the Stem I domains of the glycyl T-boxes from L. monocytogenes, C. tetani, S. pneumoniae, S. pyogenes, and B. subtilis tRNAGlyGCC; 2 μM of the tRNAGly transcript and 0.5, 1, 1.5, 2, 3, 5, and 10 μM of each Stem I domain were mixed with 1× binding buffer (10 mM Tris–HCl, pH 7.4, 100 mM KCl, 10 mM MgCl2) and were refolded at 90°C for 3 min, and then they were snap-cooled (−3°C/sec) to 12°C and incubated for 5 min. Subsequently, 2 μM of the tRNAGly transcript in 1× binding buffer was mixed with each concentration of Stem I also in 1× binding buffer, and 1U RNasin was added to each reaction. The reactions were incubated at room temperature for 30 min. After the incubation, the reactions were placed on ice for 10 min. Before their loading in 6% Native PAGE, they were mixed with 1× Native gel loading buffer (50 mM Tris–HCl pH 7.0, 10% v/v glycerol, 0.012% w/v bromophenol blue, 0.012% w/v xylene cyanol in DEPC-treated H2O). The gel was pre-run at 150 V for 20 min and the reactions ran at 150 V for ∼4–5 h. Two control reactions were used for each Stem I domain, one that contained only 2 μM of the tRNAGly ligand and one that contained 1.5 μM of the respective Stem I domain. The gels were then dyed in 5% EtBr in ddH2O, and they were visualized in a UV lamp. The gels were further analyzed in AIDA image analyzer software (Raytest version 5.0). The tRNAGly band was quantified in each reaction, and the integral-background value for the tRNAGly in the presence of each concentration for each Stem I was subtracted from the integral-background value for the control tRNAGly reaction, to quantify the value of the tRNAGly that formed a complex with Stem I. Then the value of the bound tRNAGly was divided with the value of the control tRNAGly reaction. GraphPad Prism Software (version 8.0.1) was used to plot these values in nonlinear regression and to generate the corresponding Scatchard plots (Bound/free vs. Bound), and the Kd (dissociation constants) were calculated via one-site-specific binding analysis = according to the equation: slope = −1/Kd. All experiments were performed in duplicate, and each value used in Kd determination was the mean of two distinct measurements.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
The research project was supported by the Hellenic Foundation for Research and Innovation (H.F.R.I.) under the “1st Call for H.F.R.I. Research Projects to support Faculty Members and Researchers and the Procurement of High-cost Research Equipment Grant” (Project Number: 739) to C.S., and by the Intramural Research Program (IRP) of the National Institutes of Health (NIH), the National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK) (ZIADK075136 to J.Z.). The publication fees of this manuscript have been financed by the Research Council of The University of Patras. This work was funded by the Hellenic Foundation for Research and Innovation (HFRI-FM17-739 to C.S.), NIH IRP, and NIDDK (ZIADK075136 to J.Z.)
Author contributions: N.G., A.K., J.Z., V.S., and C.S contributed to the manuscript writing and the figures design and approved the article for publication.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080071.124.
- Received April 22, 2024.
- Accepted June 17, 2024.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Nikoleta Giarimoglou is the first author of this paper, “Structural idiosyncrasies of glycyl T-box riboswitches among pathogenic bacteria.” Nikoleta is a recent PhD graduate from the RNA biology group at the Department of Biochemistry, School of Medicine, University of Patras (you can follow us at: https://www.facebook.com/RNAupatras). The main focus of their research was the functional and structural characterization of glycyl T-box riboswitches from prominent human pathogens
What are the major results described in your paper and how do they impact this branch of the field?
The major results described in our paper reveal significant structural variations in T-box riboswitches among different bacterial species. Specifically, we found that while L. monocytogenes and C. tetani glycyl T-boxes maintain the characteristic structural features of T-boxes, S. pneumoniae and S. pyogenes exhibit notable deviations like the lack of the conserved K-turn motif, which is typically considered essential for their function. Another notable finding is the presence of a Stem II domain in the S. pyogenes glyQS T-box, which is absent from all the previously characterized glycyl T-boxes.
These findings challenge the previously held notion that the K-turn motif is universally required for T-box functionality and highlight the adaptability of T-box riboswitches to different environmental conditions and regulatory needs. Our research underscores the evolutionary plasticity of T-box riboswitches, demonstrating that structural variations can still support their regulatory role in gene expression despite the absence of canonical motifs.
By providing detailed structural and functional analyses of glycyl T-box riboswitches from prominent human pathogens, our study contributes to a deeper understanding of bacterial RNA-dependent regulatory systems and their adaptation to diverse environmental niches. This knowledge can pave the way for future research aimed at developing RNA-targeting therapeutics and further exploring the evolutionary mechanisms that underpin bacterial gene regulation.
What led you to study RNA or this aspect of RNA science?
Our interest in studying RNA, and particularly T-box riboswitches, was sparked by the interplay between two RNA molecules and the fascinating complexity and versatility of RNA molecules in regulating gene expression. What we wanted to explore were alternative roles of tRNA molecules outside translation. The T-box:tRNA system fascinated us because of its unique mechanism where the T-box recognizes and interacts with its cognate tRNA molecule through a codon–anticodon-like interaction, unlike the broader class of riboswitches that typically bind relatively small ligands. These T-box riboswitches bind cognate tRNAs that are closely linked metabolically to the genes they regulate, and they exert their mechanism by sensing the aminoacylation status of their cognate tRNAs. These intricate ways in which RNA structures interact with various cellular components to modulate biological processes, especially under different environmental conditions, inspired us to explore these mechanisms further. Moreover, understanding how pathogenic bacteria adapt their regulatory systems at the RNA level has significant implications for developing new therapeutic strategies, driving our passion to delve deeper into this aspect of RNA science.
During the course of these experiments, were there any surprising results or particular difficulties that altered your thinking and subsequent focus?
During the course of our experiments, we encountered several surprising results that prompted us to reevaluate our initial hypotheses and adjust our research focus accordingly. One notable finding was the absence of the K-turn motif in the streptococcal glycyl T-box riboswitches, which challenged conventional understanding of their structural requirements for function. Moreover, while Stem II-containing T-boxes typically rely on the K-turn for accurate positioning of Stem II, our study of the S. pyogenes glyQS T-box revealed the presence of a Stem II domain without a K-turn. These discoveries led us to explore alternative structural elements such as the C-loop motif and consider their implications for tRNA binding and regulatory mechanisms. Additionally, navigating the variability in T-box riboswitch structures among different bacterial pathogens posed significant analytical challenges, requiring us to refine our experimental approaches, such as finding the correct conditions for the proper finding of each RNA molecule and molecular techniques to accurately characterize these regulatory RNA elements.
What are some of the landmark moments that provoked your interest in science or your development as a scientist?
Several landmark moments have significantly shaped my interest in science and development as a scientist. Early exposure to hands-on laboratory experiments during my undergraduate studies sparked my fascination with scientific inquiry and discovery. However, it was working alongside this particular research group and under the guidance of my supervisor, Professor C. Stathopoulos, a specialist with a deep passion for tRNA molecules, that truly molded my scientific identity. Their mentorship, dedication to rigorous research, and encouragement to explore new ideas have been pivotal in shaping my approach to scientific investigation and fostering my passion for unraveling complex biological phenomena.
If you were able to give one piece of advice to your younger self, what would that be?
If I could advise my younger self, I would say to embrace uncertainty and change. Life often takes unexpected turns, and it is okay not to have everything figured out right away. Trust in your abilities to adapt and learn from every experience, as these are the moments that shape you into who you are meant to become. Viewing setbacks as opportunities for growth, rather than obstacles, has been crucial in developing my resilience and determination in pursuing scientific inquiry, and this is something that I try to remind myself every day.








