
Rational design of eukaryotic riboswitches that up-regulate IRES-mediated translation initiation with high switching efficiency through a kinetic trapping mechanism in vitro
- Corresponding author: ogawa.atsushi.mf{at}ehime-u.ac.jp
Abstract
In general, riboswitches functioning through a cotranscriptional kinetic trapping mechanism (kt-riboswitches) show higher switching efficiencies in response to practical concentrations of their ligand molecules than eq-riboswitches, which function by an equilibrium mechanism. However, the former have been much more difficult to design due to their more complex mechanism. We here successfully developed a rational strategy for constructing eukaryotic kt-riboswitches that ligand-dependently enhance translation initiation mediated by an internal ribosome entry site (IRES). This was achieved both by utilizing some predicted structural features of a highly efficient bacterial kt-riboswitch identified through screening and by completely decoupling an aptamer domain from the IRES. Three kt-riboswitches optimized through this strategy, each responding to a different ligand, exhibited three- to sevenfold higher induction ratios (up to ∼90) than previously optimized eq-riboswitches regulating the same IRES-mediated translation in wheat germ extract. Because the IRES used functions well in various eukaryotic expression systems, these types of kt-riboswitches are expected to serve as major eukaryotic gene regulators based on RNA. In addition, the present strategy could be applied to the rational construction of other types of kt-riboswitches, including those functioning in bacterial expression systems.
Keywords
INTRODUCTION
Riboswitches are cis-acting and molecule-responsive gene regulatory RNAs that have been identified in nature primarily in untranslated regions (UTRs) of bacterial mRNAs (Roth and Breaker 2009). They are composed of two domains that are in general partially complementary to and/or overlapping with each other. The two domains are an aptamer, which recognizes its specific ligand molecule, and an expression platform (EP), which directly controls gene expression. When the former binds to its ligand, a conformational change or alternative folding of the latter is induced, which turns OFF or ON the downstream (or sometimes upstream) gene's expression.
We can utilize an in vitro selection method to obtain a user-defined ligand-specific aptamer (Ellington and Szostak 1990; Tuerk and Gold 1990), even though natural riboswitches (or their aptamer domains) are restricted to binding endogenous metabolites due to the limited variety of ligand molecules. By inserting such an in vitro-selected aptamer into mRNA and functionally linking it to an EP, it is possible to artificially construct a riboswitch that responds to the aptamer's ligand (Etzel and Mörl 2017; Hoetzel and Suess 2022). In fact, various types of artificial riboswitches (including those that function in eukaryotic expression systems) have been thus far constructed in this way. Since they can control the expression of arbitrary genes in a ligand-dose-dependent manner, they are expected to be versatile gene regulatory elements for creating biosensors and gene circuits in both cellular (Topp and Gallivan 2010; Chang et al. 2012; Ono et al. 2020; Fukunaga et al. 2023) and cell-free (Hodgman and Jewett 2012; Lentini et al. 2016; Thavarajah et al. 2020; Chushak et al. 2021; Tabuchi and Yokobayashi 2021) synthetic biology.
To successfully link an aptamer and an EP in constructing artificial riboswitches, it is useful to randomize sequences around the inserted aptamer and then screen them using the switching efficiency (i.e., the ON/OFF expression ratio) as an indicator (Lynch and Gallivan 2009; Felletti et al. 2016; Tabuchi and Yokobayashi 2022). In contrast, some methods to rationally or computationally design and slightly optimize the linkage sequences have also been reported (Ogawa 2011; Ceres et al. 2013; Wachsmuth et al. 2013; Borujeni et al. 2016). We have developed several unique rational methods to construct artificial riboswitches that control eukaryotic translation initiation, including more useful up-regulators, ON-riboswitches (Ogawa 2011, 2013; Ogawa et al. 2017a, 2023, Ogawa and Itoh 2020; Takahashi et al. 2022). Although it is impossible to functionally link an aptamer to the eukaryotic canonical translation platform, the 5′ end (or the 5′ cap), for ON-riboswitch construction (because there is no space upstream of the 5′ end), we have successfully constructed eukaryotic ON-riboswitches by using atypical EPs, such as internal ribosome entry sites (IRES).
However, the ON/OFF induction ratios of these eukaryotic ON-riboswitches are generally lower than those of artificial ON-riboswitches (especially ones obtained by screening methods) that regulate translation initiation in bacterial expression systems. For example, our eukaryotic ON-riboswitches responsive to theophylline, which has been commonly used as a ligand for artificial riboswitches, have ON/OFF ratios of around 10 even at 1 mM theophylline, which is up to sevenfold lower than those of theophylline-responsive, bacterial ON-riboswitches under similar cell-free conditions (Mishler and Gallivan 2014). This large difference in switching efficiencies can be attributed primarily to differences in regulatory mechanisms. We designed our previous eukaryotic ON-riboswitches to function by a thermodynamic equilibrium mechanism based on structures of post-transcriptional mRNA (Fig. 1A; Supplemental Fig. S1A). In contrast, the highly efficient bacterial ON-riboswitches obtained by screening methods function by a kinetic trapping mechanism that relies on mRNA structures during transcription (Fig. 1B; Supplemental Fig. S1B), similar to many natural bacterial riboswitches (Mishler and Gallivan 2014).
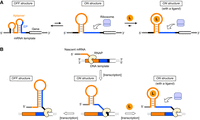
Comparison between an eq-ON-riboswitch and a kt-ON-riboswitch that each regulates translation initiation. (A) Schematic of an eq-ON-riboswitch. Although the inactive OFF structure (left) is more stable than the active ON structure (middle), both structures are in a dynamic equilibrium due to the relatively small ΔΔGTS. Therefore, ligand molecules can shift the equilibrium toward the latter (right). See the energy diagram in Supplemental Figure S1A. (B) Schematic of a kt-ON-riboswitch. This type of riboswitch folds into the ON structure (middle) right after transcription of its aptamer domain. While the reverse transition to the OFF structure takes place rapidly (left) after transcription of the downstream EP (before the ribosome binds to the EP intermolecularly), it is inhibited by stabilization with ligand binding for the subsequent translation (right). Note that the forward transition (i.e., retransition from OFF to ON) is considerably suppressed even in the presence of ligand molecules due to the large ΔΔGTS, which prevents the system from reaching equilibrium. RNAP means RNA polymerase. See the energy diagram in Supplemental Figure S1B.
We report here a rational design method for developing highly efficient eukaryotic ON-riboswitches that regulate translation initiation by the kinetic trapping mechanism. Three riboswitches constructed through this method, each having a different aptamer specific to its ligand, exhibited three- to sevenfold higher induction ratios (up to ∼90) than the previously reported eukaryotic ON-riboswitches that regulate the same IRES-mediated translation by the equilibrium mechanism. Given the versatility of the IRES used for the design, the present types of IRES-based ON-riboswitches are expected to be used as major eukaryotic gene regulators. In addition, the strategy of our rational method could be applied to creating bacterial riboswitches and other types of eukaryotic riboswitches that function by the kinetic trapping mechanism.
RESULTS AND DISCUSSION
Comparison between equilibrium and kinetic trapping mechanisms
Considering one type of ON-riboswitch that functions in the equilibrium mechanism (eq-ON-riboswitches) as a set of molecules, the structures of those molecules are in a dynamic equilibrium between an inactive OFF structure and a less stable but active ON structure, because the energy barrier for the forward transition from the OFF structure to the ON structure (ΔΔGTS) is not so large (Fig. 1A; Supplemental Fig. S1A). This equilibrium shifts toward the former in the absence of ligand molecules (i.e., under OFF conditions) but toward the latter in the presence of them (i.e., under ON conditions). Therefore, to achieve a sufficiently high ON/OFF induction ratio in this mechanism, the ligand-free equilibrium constant should be as low as possible and the ligand concentration should be quite high (Beisel and Smolke 2009). However, such high concentrations often inhibit expression itself and/or are difficult to prepare. Nonetheless, if an eq-ON-riboswitch is designed so that the OFF structure is less stable in order to make the riboswitch responsive to low to moderate ligand concentrations, more frequent leakage of gene expression would occur under OFF conditions, leading to lower switching efficiency.
In contrast, an ON-riboswitch functioning in the kinetic trapping mechanism (kt-ON-riboswitch) determines its structure during transcription from DNA to mRNA as shown in Figure 1B and Supplemental Figure S1B (Mishler and Gallivan 2014). Since transcription occurs from the 5′ end, the upstream aptamer domain folds into an ON structure right after its transcription whether under OFF or ON conditions. Under the former conditions, after (or during) transcription of the subsequent downstream EP, the reverse transition to an OFF structure occurs quickly (Fig. 1B, center to left). On the other hand, when the aptamer binds to its ligand before the reverse transition (or it folds into the ON structure using its ligand as a template), the ON structure is well stabilized, which considerably slows the reverse transition: the ligand kinetically traps the riboswitch in the ON structure (Fig. 1B, center to right). A characteristic feature of this mechanism is that ΔΔGTS is large and thus the forward transition (from OFF to ON) is considerably slow. This means that it is difficult even for the ligand to induce the forward conformational change, unlike the case in eq-ON-riboswitches. Because the kinetic trapping mechanism not only exploits the kinetic ligand/aptamer binding literally but also avoids the expression leakage under OFF conditions, it could allow kt-ON-riboswitches to exhibit higher ON/OFF ratios at low to moderate ligand concentrations (Beisel and Smolke 2009).
Structural features of a highly efficient bacterial kt-ON-riboswitch that regulates translation initiation
Compared to eq-ON-riboswitches, kt-ON-riboswitches are much more challenging to rationally design from scratch due to their more complex mechanism. Therefore, we decided to first develop a design strategy with reference to one of the highly efficient bacterial kt-ON-riboswitches identified through screening that are responsive to theophylline (riboswitch C) (Fig. 2A; Topp et al. 2010; Mishler and Gallivan 2014). Although it is difficult to predict the transition-state structure and ΔΔGTS of this riboswitch, we can predict its OFF and ON structures and their features, which should provide some clues for designing kt-ON-riboswitches. The predicted features of each structure are as follows:
-
In the OFF structure, the EP composed mainly of the bacterial ribosome binding site (RBS) forms a long 16-bp stem (with one bulge, one mismatched pair, and one G–U wobble base pair) to suppress the translation initiation. This long and stable inhibitory stem (IS) makes ΔΔGTS considerably large, severely limiting the forward transition to the ON structure (Supplemental Fig. S1B).
-
In the ON structure, the EP is freely accessible and instead the upstream aptamer forms short 5-bp upper and 4-bp lower stems (US and LS, respectively) at both ends of its core sequences to facilitate binding with the ligand (theophylline). Nonetheless, because these stems are much shorter than the IS in the OFF structure, this ON structure is much less stable without the ligand than the OFF structure. The short stems also make the energy barrier for the reverse transition, ΔΔGTS(rev), small, easily transforming the ON structure to the OFF structure unless the ligand binds to the aptamer (Supplemental Fig. S1B).
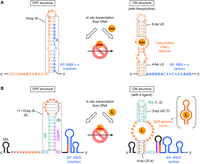
Rational design of eukaryotic kt-ON-riboswitches with reference to a bacterial kt-ON-riboswitch. (A) Predicted secondary structures of a highly efficient bacterial kt-ON-riboswitch that regulates translation initiation in response to theophylline (riboswitch C). The aptamer core sequences and the EP core sequences (i.e., the RBS and the start codon) are shown in bold. A 5′ region of the aptamer sometimes forms a weak secondary structure in the OFF structure, depending on the upstream sequence. (B) Predicted secondary structures of rationally designed eukaryotic kt-ON-riboswitches that regulate translation initiation mediated by the PSIV IRES. The IS length varies depending on the 3′-side LS sequence (ZZZZ), which hybridizes to the 5′-side LS (XXXX) in the ON structure. The secondary structures of the IRES are roughly simplified. The red bars represent the 8-nt aIRES (see Supplemental Fig. S2). The purposes of the 5′ terminal stem–loop structure (5SL) are to enhance T7 transcription, stabilize transcripts, and prevent the eukaryotic canonical translation. A schematic of the original splittable aptamer is shown in parentheses. The numbers in parentheses correspond to those of the design procedure in the main text.
Therefore, the key to successfully constructing a kt-ON-riboswitch should be that its EP forms a relatively long and stable IS in the OFF structure, while its aptamer domain placed upstream of the EP forms relatively short and unstable US and LS in the ON structure. It is probably possible to control ΔΔGTS and ΔΔGTS(rev) to some extent by adjusting the lengths (or stabilities) of these stems. However, although we can refer to riboswitch C for the lengths of these stems, we cannot use its bacterial EP to construct a eukaryotic riboswitch. In addition, the EP is sequestered by a part of the core sequences of the theophylline aptamer in the OFF structure, so that the arrangement of the two domains (the aptamer and the EP) cannot be directly applied to other aptamers (even if the EP could be used as-is). We thus sought not only to use an IRES for eukaryotic expression instead of the bacterial EP, but also to completely decouple the two domains (i.e., to eliminate their complementarity).
Design strategy of eukaryotic kt-ON-riboswitches that regulate translation initiation
The EP we selected for eukaryotic kt-ON-riboswitches was the intergenic IRES of the Plautia stali intestine virus (PSIV) (Sasaki and Nakashima 1999), which we have used for the rational construction of eukaryotic eq-ON-riboswitches (Ogawa 2011; Ogawa and Itoh 2020; Takahashi et al. 2022; Ogawa et al. 2023). We previously found that insertion of an 8-nt anti-IRES sequence complementary to a portion of the long IRES (aIRES) immediately before the IRES inactivated the IRES-mediated mRNA translation sufficiently (1.4% relative to an aIRES-free positive control mRNA) in a eukaryotic cell-free system, wheat germ extract (WGE) (Sawasaki et al. 2002; Ogawa 2011). We also previously confirmed that a further upstream 8-nt sequence complementary to the aIRES (an anti-aIRES sequence, aaIRES) reactivated the translation well by forming the aaIRES/aIRES duplex (79% to the control) (Ogawa 2011). This terminal aaIRES is more promising as a target to be easily sequestered in the OFF structure than an internal part of the IRES is. However, since the lengths of aIRES and aaIRES were optimized for conventional mRNA translation in WGE, we here used DNA templates instead of mRNA to reexamine the optimal lengths in a coupled in vitro transcription/translation (cIVTT) system based on T7 transcription and WGE translation, which we have recently reported (Supplemental Fig. S2; Takahashi and Ogawa 2021). The results showed that the 8-nt aIRES inhibited gene expression in the cIVTT system as well as in the mRNA translation (1.1% relative to an aIRES-free positive control DNA, PC-DNA-Y), while the 8-nt aaIRES restored the expression efficiency insufficiently (37% to PC-DNA-Y), and two more nucleotides were required to exhibit a sufficient recovery (77% to PC-DNA-Y) in the cIVTT system. We thus regarded the PSIV IRES with the 8-nt aIRES and the 10-nt aaIRES as the whole EP and set the aaIRES as the target site to be inhibited in an IS in the OFF structure.
Based on the above results, we designed eukaryotic kt-ON-riboswitches through the following steps (Fig. 2B):
-
Replace a functionally unimportant intermediate loop of a stem (corresponding to a US) in a splittable aptamer (i.e., an aptamer that can be split at a loop into two parts without significant loss of affinity for its ligand) with a sequence complementary to the target aaIRES (a modulator sequence, MS). This allows us to completely decouple the aptamer core from the EP.
-
Set the MS to the exact same 8-nt sequence as the aIRES (5′ AGACCUCU 3′) to use its possible internal 2-bp stem at its ends (5′ AG 3′/5′ CU 3′) as a part of the US in the ON structure.
-
Adjust the US length to 5 bp, the same as in riboswitch C, by adding a 3-bp stem (5′ GCG 3′/5′ CGC 3′) to the MS ends instead of the original US.
-
Adjust the LS length to 4 bp, the same as in riboswitch C. The LS sequences depend on the sequence of the 3′-side aptamer core (vide infra).
-
Insert a GCG triplet between the 3′-side LS and the aaIRES so that it could bind to a part of the 3′-side US to form an 11-bp IS including the 8-bp MS/aaIRES in the OFF structure.
-
Optimize the IS length by varying the LS sequences so that the 3′-side LS could partially (or fully) bind to the 3′-side aptamer core to extend the perfectly matched IS up to 15 bp, which should be more stable than the imperfectly matched 16-bp IS in riboswitch C.
It should be noted that there is no need to sequester the 5′-side LS and/or the 5′-side aptamer core with another stem in the OFF structure if the IS is stable enough. Such a stem would adversely prevent the riboswitch from folding into the ON structure before the transcription of the EP.
PSIV IRES-based eukaryotic kt-ON-riboswitches responsive to theophylline
To demonstrate this design strategy, we first chose as a model an in vitro-selected splittable aptamer binding to theophylline (KD is ∼300 nM) (Jenison et al. 1994; Chang et al. 2014), incorporated the aptamer into the design, and optimized the IS length. Specifically, we individually incubated DNA templates, each encoding a designed riboswitch with 11, 12, 13, 14, or 15 bp of IS (theo-ISL, where L represents 11, 12, 13, 14, or 15, respectively, as shown in Fig. 3A), followed by the YPet gene (Nguyen and Daugherty 2005). These incubations were carried out in the WGE-based cIVTT system, with or without theophylline at a concentration of 1 mM (Fig. 3B). In the absence of theophylline (i.e., under an OFF condition), the expression was suppressed in theo-IS14 and theo-IS15 as well as in the aIRES control (T7-aa0/a8-Y, Supplemental Fig. S2). This indicated that the ΔΔGTS(rev) values of theo-IS14 and theoIS-15 were small enough to efficiently induce the reverse transition to the OFF structures (or they just folded into the OFF structures) and that their ΔΔGTS values were large enough to suppress the retransition to the ON structures (or their equilibria just shifted well toward the OFF structures). However, as the IS length was shortened, the expression leakage gradually increased. Because ΔΔGTS(rev) is not likely to be largely affected by the IS length, this increase in expression leakage can be attributed mainly to more frequent retransitions with smaller ΔΔGTS. In the presence of 1 mM theophylline (i.e., under an ON condition), higher expression efficiencies were observed in theo-IS12, theo-IS13, and theo-IS14, suggesting that the ligand trapped these riboswitches in the ON structures and/or shifted their equilibria to the ON structures. One possible reason for the precipitous loss of efficiency in theo-IS15 is that the sequence of the 5′-side LS is exactly the same as that of the 5′ region of the 3′-side aptamer core (5′ CCUU 3′): the 3′-side LS right after its transcription binds not to the 5′-side LS to form the LS but quickly to the closer, 3′-side aptamer core, which prevents the riboswitch from folding into the ON structure in the first place. Consequently, theo-IS14 exhibited the highest ON/OFF induction ratio, 35.4 ± 4.8. This switching efficiency is fourfold higher than that of the best PSIV IRES-based eq-ON-riboswitch (theo5) at the same concentration (1 mM) of theophylline when using mRNA as a template in WGE (Ogawa 2011) and is more than half that of riboswitch C, the referenced bacterial kt-ON-riboswitch, at 2 mM theophylline in a bacterial cell-free cIVTT system (Mishler and Gallivan 2014). Incidentally, when we tested theo5 by using its DNA template with the YPet gene in the WGE-based cIVTT system, it showed a moderate ON/OFF ratio (13.2 ± 4.0 at 1 mM theophylline), which was roughly equivalent to its ON/OFF ratio with conventional mRNA translation in WGE, indicating that theo5 does not function well in the kinetic trapping mechanism.
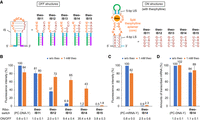
Optimization of PSIV IRES-based eukaryotic kt-ON-riboswitches responsive to theophylline. (A) Predicted partial secondary structures of rationally designed eukaryotic kt-ON-riboswitches with the split theophylline aptamer (theo-ISL). (B) Relative fluorescence intensities of YPet expressed from DNA templates encoding theo-ISL before the YPet gene (T7-theo-ISL-Y) in the absence or presence of 1 mM theophylline (theo) in the WGE-cIVTT system. (C) Relative fluorescence intensities of YPet expressed by conventional translation of mRNA with theo-IS14 (theo-IS14-Y) in the absence or presence of 1 mM theophylline in WGE. (D) Relative amounts of mRNA transcribed from the DNA template encoding theo-IS14 (T7-theo-IS14-Y) in the absence or presence of 1 mM theophylline under the cIVTT conditions. The error bars represent the standard deviation (n = 4).
To confirm if theo-IS14 is a kt-ON-riboswitch, we next directly used in vitro-transcribed mRNA instead of DNA as a template for conventional translation in WGE (Fig. 3C; Sawasaki et al. 2002; Takahashi and Ogawa 2021). As a result, the ON/OFF ratio at 1 mM theophylline decreased considerably, to 2.5 ± 0.6, with much lower expression efficiency, showing that ΔΔGTS of theo-IS14 is undoubtedly large enough to suppress the forward transition even with the ligand: theo-IS14 does not function well in the equilibrium mechanism, but does function well mainly in the kinetic trapping. Incidentally, we also confirmed that theophylline did not affect mRNA production (i.e., T7 transcription) under the cIVTT conditions except in the absence of WGE (Fig. 3D).
We then carried out some experiments to characterize theo-IS14 regarding ligand dose–response, ligand specificity, and versatility. First, the DNA template encoding theo-IS14 (and the YPet gene) was incubated at various concentrations of theophylline in the cIVTT system (Fig. 4A). The ON/OFF induction ratio increased dose-dependently with an approximately fourfold higher sensitivity (precisely, with an approximately fourfold steeper linear slope at least from 1 µM to 100 µM) than that of theo5 (Ogawa 2011). However, theo-IS14 showed no response to caffeine (an N7-methyl analog of theophylline) even at 1 mM, reflecting high specificity to theophylline (Fig. 4B, center). In addition, when mutations (U24A and G26A) were introduced into the aptamer core to prevent binding to theophylline (m-theo-IS14), responsiveness to theophylline was completely lost (Fig. 4B, right). These results clearly show that specific ligand/aptamer binding is required to turn the expression on. Lastly, the YPet gene was substituted with the nanoluciferase (nLuc) gene (Hall et al. 2012) to verify the versatility of theo-IS14 (Fig. 4C). Given the fact that this gene alteration had little effect on the ON/OFF ratio, theo-IS14 functions independently from the regulated gene and is thus likely to be able to regulate any gene.

Characterization of theo-IS14. (A) The ON/OFF induction ratios of theo-IS14 at various concentrations of theophylline. Inset: Expansion at low concentrations. (B) Relative fluorescence intensities and images of YPet expressed from DNA templates encoding theo-IS14 or m-theo-IS14 before the YPet gene (T7-theo-IS14-Y or T7-m-theo-IS14-Y, respectively) in the absence or presence of 1 mM theophylline (theo) or caffeine (caf) in the WGE-cIVTT system. The leftmost lane is the positive control (PC-DNA-Y). A schematic of the mutations in m-theo-IS14 is also shown. (C) Relative luminescence intensities and images of nLuc expressed from the DNA template encoding theo-IS14 before the nLuc gene (T7-theo-IS14-nL) in the absence or presence of 1 mM theophylline in the WGE-cIVTT system. The error bars represent the standard deviation (n = 4).
PSIV IRES-based eukaryotic kt-ON-riboswitches responsive to other ligands
Finally, we used two other in vitro-selected splittable aptamers to further validate the design strategy. One is an aptamer binding to tetracycline with a two-order faster binding rate and a three-order higher affinity (KD is 0.77 nM) (Müller et al. 2006; Förster et al. 2012; Chang et al. 2014). Because the optimal IS length should depend on the sequence and/or structure of the aptamer used, several kinds of riboswitches, each with the split tetracycline-binding aptamer and a different length of IS, were designed (tc-ISL, wherein L represents the IS length, Fig. 5A), as in the case of the theophylline-responsive riboswitches. When each DNA template encoding tc-ISL (and the YPet gene) was incubated in the cIVTT system, tc-IS12 showed the highest ON/OFF ratio, 87.4 ± 3.8 at 100 µM tetracycline (Fig. 5C), which was sevenfold higher than that of the best PSIV IRES-based eq-ON-riboswitch responsive to tetracycline (12.6 ± 1.6 at 100 µM tetracycline by tc7 with a negligible U55A mutation in the aptamer) (Müller et al. 2006; Ogawa 2011). This highly efficient riboswitch was also expected to function with the kinetic trapping mechanism judging from the results that in vitro-transcribed mRNA significantly lost switching efficiency and that tetracycline did not affect T7 transcription (Supplemental Fig. S3). Therefore, its much higher ON/OFF ratio compared to that of theo-IS14 at 100 µM theophylline (14.3 ± 1.8) is due to faster ligand/aptamer binding in addition to stronger binding, both of which should increase the expression efficiency under ON conditions.
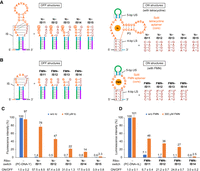
PSIV IRES-based eukaryotic kt-ON-riboswitches responsive to other ligands. (A,B) Predicted partial secondary structures of rationally designed eukaryotic kt-ON-riboswitches with the split tetracycline aptamer (tc-ISL) and the split FMN aptamer (FMN-ISL), respectively. The circled nucleotides probably hybridize to their complementary nucleotides to form the P3 duplex. The 5′ terminal A in the 5′-side tetracycline aptamer's core may hybridize to the 3′ terminal U in the 3′-side thereof in the ON structure. The A*–U* base pair is required for binding to FMN. It is impossible to design FMN-IS15 because the IS's loop would be only 2 nt. (C) Relative fluorescence intensities of YPet expressed from DNA templates encoding tc-ISL before the YPet gene (T7-tc-ISL-Y) in the absence or presence of 100 µM tetracycline (tc) in the WGE-cIVTT system. (D) Relative luminescence intensities of nLuc expressed from DNA templates encoding FMN-ISL before the nLuc gene (T7-FMN-ISL-nL) in the absence or presence of 300 µM FMN in the WGE-cIVTT system. The error bars represent the standard deviation (n = 4).
The other aptamer binds to flavin mononucleotide (FMN) with a binding rate and affinity comparable to those between theophylline and its aptamer (KD is 500 nM) (Burgstaller and Famulok 1994; Chang et al. 2014). Similarly, several kinds of DNA templates, each encoding a riboswitch with the split FMN-binding aptamer and a different length of IS (FMN-ISL, wherein L represents the IS length, Fig. 5B), were prepared. Incidentally, since FMN is a green fluorescent molecule, the nLuc gene was used instead of the YPet gene. As a result of the cIVTT reactions of these templates, FMN-IS13 exhibited the highest ON/OFF ratio, 24.8 ± 0.7 at 300 µM FMN (Fig. 5D), which is comparable to that of theo-IS14 at 300 µM theophylline (24.3 ± 2.1) and is threefold higher than that of the best PSIV IRES-based eq-ON-riboswitch responsive to FMN (FMN4) at the same concentration of FMN (Ogawa 2011). These results suggest that FMN-IS13 is also a kt-ON-riboswitch, which was confirmed by the same experiments as those performed on theo-IS14 and tc-IS12 (Supplemental Fig. S4).
Conclusions
In summary, we successfully established a rational strategy for using in vitro-selected aptamers to design and optimize eukaryotic kt-ON-riboswitches that ligand-dependently up-regulate PSIV IRES-mediated translation. The success of this approach relied on referencing the OFF and ON structures of a highly efficient bacterial kt-ON-riboswitch identified through screening, enabling us to bypass direct consideration of ΔΔGTS values and complex kinetic parameters. Additionally, decoupling the two domains made this strategy applicable to various aptamers. The kt-ON-riboswitches optimized through this strategy exhibited three- to sevenfold higher ON/OFF ratios (up to approximately 90) than previously reported eq-ON-riboswitches based on the PSIV IRES at low to moderate ligand concentrations. To obtain higher switching efficiency, a ligand/aptamer pair that forms a more stable complex more quickly would be better. Although an aptamer to be implanted must be splittable without impairing its function, such an aptamer can be obtained via in vitro selection from an initial pool with a stem–loop structure (Ogawa 2015). In addition, the PSIV IRES functions well in various cell-free and cell-based eukaryotic expression systems (and surprisingly also in bacteria) (Kamoshita et al. 2009; Hertz and Thompson 2011; Colussi et al. 2015; Ogawa et al. 2017b). As a result, it is anticipated that these PSIV IRES-based kt-ON-riboswitches can be generated using different splittable aptamers and will serve as prominent regulators of cell-free (and possibly cell-based) eukaryotic gene expression. Other types of Dicistroviridae intergenic IRES can also be used (with the corresponding aaIRES/aIRES pairs) instead of the PSIV IRES depending on the expression system used (Hertz and Thompson 2011). Moreover, the present design strategy itself could be applied to the rational construction of bacterial kt-riboswitches and other types of eukaryotic kt-riboswitches, including those regulating transcription termination or self-cleavage. Further studies along these lines are under way.
MATERIALS AND METHODS
Synthetic nucleic acids
Primers for PCR were synthesized by Eurofins Genomics or Thermo Fisher Scientific. Their sequences are described in the Supplemental Material. The sequences of three synthetic plasmids, pTheo5-PSIV-IRES encoding the PSIV IRES, pHis-SRY-YPet encoding the YPet gene, and pEU-nLuc-Venus encoding the nLuc gene, have been reported elsewhere (Ogawa and Itoh 2020; Ogawa et al. 2014, 2017a, 2023).
Preparation of DNA templates
A 5′ segment encoding the PSIV IRES and a 3′ segment encoding the YPet (or nLuc) gene with an 1183-nt (or 106-nt) 3′-UTR were sequentially PCR-amplified with PrimeSTAR MAX DNA Polymerase (Takara Bio) from pTheo5-PSIV-IRES and pHis-SRY-YPet (or pEU-nLuc-Venus), respectively. The two segments digested by Spe I were ligated with Ligation high Ver.2 (Toyobo) to construct core DNA (Supplemental Table S1; Ogawa 2014). A DNA template for the WGE-based cIVTT system was then constructed by using core DNA as the first template in the post-ligation sequential PCRs (Supplemental Table S2). A workflow schema for the template preparation is shown in Supplemental Figure S5. Incidentally, all PCR products in the sequential PCRs, except those in the final PCR, were agarose gel-purified with a NucleoSpin Gel and PCR Clean-up kit (Takara Bio) and then used as templates in the next PCR. The final PCR products were simply purified with the same kit and quantified by absorbance at 260 nm.
cIVTT in WGE
cIVTT was performed in WGE under previously optimized conditions (Takahashi and Ogawa 2021). A total of 10 µL of mixture was prepared, consisting of a DNA template at a concentration of 5 nM, various concentrations of ligand molecules, T7 RNA Polymerase at a concentration of 2.5 U/µL (containing 2.5 v/v% glycerol), additional NTPs/MgCl2 at a concentration of 1.5 mM, WEPRO1240 (WGE, 2 µL), creatine kinase (CK) at a concentration of 40 ng/µL, and SUB-AMIX at a 1× concentration (containing 2.7 mM Mg(OAc)2, 1.2 mM ATP, and 0.25 mM GTP). The latter three components (WEPRO1240, CK, and SUB-AMIX) were part of the WEPRO1240 Expression Kit (CellFree Sciences). The mixture was then incubated at 26°C for 3 h. The translation efficiency was evaluated using either the fluorescence intensity of translated YPet or the chemiluminescence intensity of translated nLuc, as previously described (Ogawa et al. 2017a; Takahashi and Ogawa 2021). For translation-free transcription under the cIVTT conditions, WEPRO1240 and CK were not added. The obtained transcripts were purified with a QIAquick Nucleotide Removal Kit (Qiagen) and quantified by absorbance at 260 nm.
Conventional mRNA translation in WGE
Each mRNA was prepared by in vitro transcription of the corresponding DNA template with T7 RNA Polymerase ver.2.0 (Takara Bio) according to the manufacturer's protocol. The obtained transcripts were purified and quantified as described above. Cell-free translation of mRNA (3 pmol) was carried out with the WEPRO1240 Expression Kit in a 10 µL solution in the absence or presence of ligand molecules (specific concentrations indicated) at 26°C for 3 h, as previously described (Ogawa 2011; Takahashi and Ogawa 2021). The translation efficiency was evaluated as described above.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
This work was partially supported by Japan Society for the Promotion of Science (JSPS) KAKENHI Grant Number JP19K05697.
Author contributions: A.O. designed the project. H.T. and A.O. wrote the manuscript. All authors cooperatively performed the experiments and analyzed the data.
Footnotes
-
Abbreviations: UTR, untranslated region; EP, expression platform; IRES, internal ribosome entry site; eq-ON-riboswitch, up-regulating riboswitch functioning in an equilibrium mechanism; kt-ON-riboswitch, up-regulating riboswitch functioning in a kinetic trapping mechanism; RBS, ribosome binding site; IS, inhibitory stem; US, upper stem; LS, lower stem; PSIV, Plautia stali intestine virus; PC, positive control; WGE, wheat germ extract; cIVTT, coupled in vitro transcription/translation; aIRES, anti-IRES sequence; aaIRES, anti-aIRES sequence; MS, modulator sequence; 5SL, 5′ stem–loop; nLuc, nanoluciferase; FMN, flavin mononucleotide
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079778.123.
- Received July 20, 2023.
- Accepted September 5, 2023.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








