
Rho-dependent transcription termination: mechanisms and roles in bacterial fitness and adaptation to environmental changes
- Thuy Duong Do1,2,
- Nara Figueroa-Bossi3,
- Johnathan C. Black1,2,
- Eric Eveno1,
- Mildred Delaleau1,2,
- Lionello Bossi3 and
- Marc Boudvillain1,2
- 1Centre de Biophysique Moléculaire, CNRS UPR4301, affiliated with Université d'Orléans, 45071 Orléans cedex 2, France
- 2ED 549, Sciences Biologiques & Chimie du Vivant, Université d'Orléans, 45067 Orléans Cedex 2, France
- 3Université Paris-Saclay, CEA, CNRS, Institut de Biologie Intégrative de la Cellule (I2BC), 91190 Gif-sur-Yvette, France
- Corresponding author: marc.boudvillain{at}cnrs.fr
Abstract
The bacterial transcription termination factor Rho is a rare example of an RNA helicase that functions as a ring-shaped ATP-powered six-subunit motor. Recent studies have linked Rho's distinctive architecture to a variety of regulatory mechanisms that shape the bacterial transcriptome at the global scale and control the transcription of individual genes in a context-dependent manner. In this review, we provide a comprehensive overview of the molecular mechanisms by which Rho triggers transcription termination. We examine the two prevailing modes of Rho's action: the “catch-up” mode, where Rho actively translocates along RNA and collides with the RNA polymerase to terminate transcription, and the “stand-by” mode where Rho, recruited by transcription elongation factor NusG, remains poised to engage RNA polymerase at specific sites or under particular constraints. Additionally, we highlight Rho's interplay with nucleoid-structuring protein H-NS in the regulation of bacterial chromatin transcription, as well as the crucial role played by Rho in the conditional regulation of specific genomic loci. We discuss how these mechanisms contribute to the fine-tuning of gene activity and integrate into broader regulatory networks, supporting bacterial adaptation to environmental changes and resilience to external challenges.
Keywords
INTRODUCTION
Bacteria are highly adaptable microorganisms capable of surviving and thriving in diverse, often hostile environments. To cope with ecosystem fluctuations, such as changes in temperature, hydration, pH, redox potential, light exposure, nutrient availability, competition, or host immune response, they have evolved a variety of general and taxon-specific adaptive mechanisms. These mechanisms enable rapid gene expression adjustments in response to external stimuli, allowing bacteria to fine-tune metabolism, activate stress responses, and produce the virulence factors that optimize the balance between growth and survival (Zhu and Dai 2024).
A key player in this dynamic regulatory landscape is the transcriptional machinery, which governs RNA synthesis from the bacterial chromosome(s) and, when present, adaptive plasmid(s). Transcription termination is a crucial step in this process, ensuring proper gene and operon delineation while regulating their timely expression. For instance, premature termination prevents transcription from extending into the structural portions of genes or operons through a regulatory mechanism known as transcription attenuation (Turnbough 2019). Triggered by specific stimuli, transcription attenuation shuts down gene expression at an early stage, making it particularly relevant in bacteria, where transcription and translation are often tightly coupled (Woodgate and Zenkin 2023) and where the synthesis of polycistronic RNAs imposes a high metabolic cost.
Two primary mechanisms govern transcription termination in bacteria (Ray-Soni et al. 2016). Intrinsic (Rho-independent) termination relies on specific transcript motifs—a stable hairpin structure followed by a run of uracil residues—sufficient to disrupt the transcription elongation complex (TEC). In contrast, Rho-dependent termination (RDTT) requires the ring-shaped, ATP-dependent translocase/helicase Rho. Rho binds to nascent transcripts at poorly defined Rho utilization (Rut) sites and uses its ATP-dependent motor activity to disrupt the TEC (see below for details). Rho is broadly conserved across bacteria but is noticeably absent in Cyanobacteriota, Mycoplasmatota, or Streptococcaceae (D'Heygere et al. 2013; Moreira et al. 2024). Rho is also dispensable in Bacillota under laboratory growth conditions, even though its inactivation significantly alters the transcriptome in species such as Bacillus subtilis (Nicolas et al. 2012; Bidnenko et al. 2023) and Staphylococcus aureus (Mader et al. 2016; Nagel et al. 2018). While the relative contributions of intrinsic termination and RDTT vary across taxa (Ahmad et al. 2023), recent findings reveal a composite regulatory landscape where these mechanisms are not strictly independent (Mandell et al. 2022b). Notably, transcription factors, including Rho, can enhance termination at weak intrinsic terminators in B. subtilis (Mandell et al. 2022a) and Escherichia coli (Ahmad et al. 2022), blurring the distinction between factor-dependent and independent pathways. In this review, we summarize the current understanding of the mechanisms of RDTT and highlight recent examples of its integration into complex regulatory networks, underscoring its critical role in bacterial adaptation and fitness.
THE ARCHITECTURE OF THE RHO MOTOR AND ITS MECHANISM(S) OF ACTION
The Rho factor is a ring-shaped assembly of six identical subunits. Each Rho subunit is divided structurally and functionally into two major domains (Fig. 1A). The N-terminal domain (NTD) harbors the primary binding site (PBS), which mediates Rho's recognition of and binding to Rut sites in nascent transcripts. The C-terminal domain (CTD) contains motifs essential for hexamer cohesion, ATP hydrolysis, and RNA translocation, which occurs via the secondary binding site (SBS) loops within the central Rho channel. The CTD is highly conserved, while the NTD is more variable and often contains large polypeptide insertions (Fig. 1A). These insertions are particularly common in Actinomycetes and Bacteroidota (D'Heygere et al. 2013; Moreira et al. 2024) where they likely aid Rho in recognizing genuine Rut sites (Nowatzke et al. 1997; D'Heygere et al. 2015; Simon et al. 2021). In some species, the NTD insertion domain is intrinsically disordered, facilitating either liquid–liquid phase separation (Krypotou et al. 2023) or prion-like aggregation (Yuan and Hochschild 2017) of Rho.
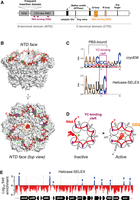
Transcription termination factor Rho and its interaction with RNA. (A) Diagram of the primary structure of Rho. (B) Structure of the closed Rho hexamer from Escherichia coli bound to λtR1 RNA (PDBs 8E3H and 8E6W). RNA is in red, with dinucleotides in YC-binding pockets shown in green. The YC-binding clefts (PBS) are in magenta, and the SBS residues in orange. (C) Consensus RNA binding motifs deduced from (top) the alignment of the λtR1 RNA segments bound to the Rho hexamer shown in B, or (bottom) differential MEME enrichment of the 2511 E. coli putative Rut peaks consistently detected by Helicase-SELEX versus 3000 Rut-less control sequences (1315 sites; E-value: 5.8 × 10−30) (Delaleau et al. 2022). (D) Diagram depicting the trajectory of RNA along the crown-like PBS site of Rho, its entry in the central Rho channel, and its interaction with the SBS loops, which triggers the closure and catalytic activation of the Rho hexamer. (E) Helicase-SELEX with Rho revealed putative Rut sites along the E. coli genome, visualized as discrete sequence enrichment peaks (blue) in this representative region of the (+) strand. Approximately one-third of the putative Rut sites are located upstream of or within genes (Delaleau et al. 2022), representing primary candidate sites for conditional RDTT.
A recent cryo-electron microscopy (cryo-EM) study of E. coli’s Rho bound to the prototypical λtR1 terminator RNA provides structural insights into how Rho's composite, crown-like PBS recognizes and binds a Rut site (Fig. 1B; Molodtsov et al. 2023). Each Rho subunit engages five consecutive single-stranded nucleotides (nt) separated by unbound linkers of 4–5 nt. These contacts extend beyond the 5′YC-binding PBS clefts (where Y represents a U or C residue) previously identified in crystal structures of E. coli’s Rho (Bogden et al. 1999; Skordalakes and Berger 2003). Although only two of these PBS clefts appear to be occupied by YC dinucleotides in the cryo-EM structure (Fig. 1B, PBS clefts in purple with bound dinucleotides in green), the nucleotide identities should be considered tentative given the resolution of the structure (4.27 Å).
Alignment of the six λtR1 RNA segments bound to the Rho subunits provides a tentative RNA consensus sequence for PBS interaction, with A and C residues preferred at most positions (Fig. 1C, top). Analysis of the hundreds of putative Rut sites recently identified in the E. coli genome by Helicase-SELEX (Fig. 1E; Delaleau et al. 2022) yields a similar, albeit shorter consensus sequence (Fig. 1C, bottom), possibly reflecting PBS preferences of individual subunits and the minimal specificity of intersubunit RNA linkers.
Overall, these findings reinforce the long-standing view that Rut sites are difficult to define, aside from their preferential localization in C>G-biased regions with limited secondary structure stability (Alifano et al. 1991; Peters et al. 2012; Nadiras et al. 2018a; Delaleau et al. 2022). The optimal Rut site length appears to be in the 60–90 nt range (Hart and Roberts 1994; Walmacq et al. 2004; Molodtsov et al. 2023), though shorter (∼30 nt) RNA sequences or hairpin-like structures can still bind Rho tightly (Schneider et al. 1993) or elicit Rho activity (Delaleau et al. 2022). This suggests that not all PBS clefts need to be occupied or bridged uniformly, to trigger Rho's catalytic isomerization. Due to these cryptic features, predicting Rut sites (and by extension, Rho-dependent terminators) is challenging, requiring cautious use of specialized algorithms (Nadiras et al. 2018a; Di Salvo et al. 2019). Nonetheless, Helicase-SELEX screening suggests that Rut sites occur only at discrete locations (Fig. 1E), collectively covering <5% of the E. coli genome (Delaleau et al. 2022). This implies that Rut site recognition by Rho is highly specific, positioning it as a key regulatory checkpoint in the RDTT pathway (see below for further discussion).
Extensive structural and biophysical data confirm that the Rho hexamer alternates between open (Skordalakes and Berger 2003; Skordalakes et al. 2005; Canals et al. 2010; Hao et al. 2021a; Said et al. 2021; Saridakis et al. 2022) and closed (Thomsen and Berger 2009; Thomsen et al. 2016; Molodtsov et al. 2023) ring conformations. The open-ring form predominates when Rho is free of RNA or during PBS-mediated RNA loading (Skordalakes and Berger 2003; Canals et al. 2010). In this state, the corkscrewed subunit arrangement is catalytically inactive but guides the RNA chain toward the central ring channel. Upon entry into the central cavity, RNA interacts with Rho's SBS loops, triggering ring closure and catalytic activation (Fig. 1D; Thomsen and Berger 2009; Thomsen et al. 2016). This conformational transition is rate limiting in vitro (Kim and Patel 2001; Walmacq et al. 2004) and serves as another important regulatory checkpoint, for instance, modulated by elongation factor NusG (Lawson et al. 2018) or blocked by the drug Bicyclomycin (Lawson et al. 2016).
Once activated, Rho translocates along the RNA in the 5′→3′ direction in an ATP-dependent manner (Steinmetz and Platt 1994; Schwartz et al. 2007, 2009), moving rapidly (Gocheva et al. 2015) but with moderate processivity (Walmacq et al. 2004; Soares et al. 2014). During translocation, SBS–RNA interactions are remodeled, while the Rut site remains bound to the PBS (Fig. 2A; Steinmetz and Platt 1994; Koslover et al. 2012; Gocheva et al. 2015). This “tethered tracking” mechanism (Steinmetz and Platt 1994) allows Rho to catch up with the RNA polymerase (RNAP), typically at transcriptional pause sites (Richardson and Richardson 1996; Guerin et al. 1998; Hollands et al. 2014). Once in contact with the RNAP, the Rho motor either fully dissociates the TEC or only disengages the RNA from the TEC, while the RNAP remains bound to the DNA template (Song et al. 2022). This “catch-up” model is backed by a wealth of studies (Table 1), including kinetic coupling experiments with Rho and RNAP velocity mutants (Jin et al. 1992; Shashni et al. 2012), and a recent cryo-EM reconstruction of the E. coli pretermination complex where Rho interacts with RNAP through its CTD face (Fig. 2B, in gray; Molodtsov et al. 2023). Elongation factor NusG stabilizes this pretermination complex by linking Rho and RNAP (Fig. 2B), potentially aiding termination by acting as a stator for the Rho motor that restricts its rotation with respect to RNAP (Molodtsov et al. 2023). The organization of this “catch-up” pretermination complex is consistent with previous structures of binary Rho–RNA (Thomsen and Berger 2009; Thomsen et al. 2016) and Rho–NusG (Lawson et al. 2018) complexes.
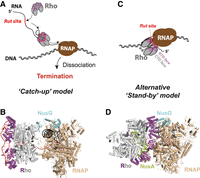
Models of Rho-dependent termination of transcription (RDTT). (A) Diagram of the “catch-up” RDTT model and (B) cryo-EM reconstruction of the “catch-up” pretermination complex (PDBs 8E3F, 8E3H, and 8E6W). (C) Diagram of the “stand-by” RDTT model and (D) cryo-EM reconstruction of the “stand-by” pretermination complex (PDB 6Z9P). Note that the Rho hexamer is in an open, inactive configuration in the “stand-by” complex. To better visualize the orientation of the Rho hexamer relative to RNAP, Rho's NTD and CTD segments are shown in magenta and gray, respectively, in all panels.
Comparison of the catch-up and stand-by models (not all relevant references are cited, for brevity)
Distinct cryo-EM reconstructions of the E. coli pretermination complex (Hao et al. 2021a; Said et al. 2021) support an alternative “stand-by” model in which an open, catalytically inactive Rho hexamer is prebound to RNAP in an inverted CTD→NTD face orientation (Fig. 2C,D). In this model, Rho binds RNAP in noncoding or untranslated regions of nascent transcripts, scans the growing RNA chain from this RNAP-bound position, and triggers termination by allosteric inactivation and destabilization of the pretermination complex upon detecting a Rut site (Epshtein et al. 2010; Hao et al. 2021a; Said et al. 2021). Although the RNA trajectory cannot be definitely determined from the cryo-EM reconstructions, it was proposed that the RNA chain might be guided by the transcription factor NusA to contact Rho's SBS before the PBS, in a reverse order when compared to the traditional “catch-up” model (Hao et al. 2021a,b). While this proposal remains subject to debate (Table 1), RNA is not required for the “stand-by” Rho-RNAP assembly, which is stabilized by NusA and, to a lesser extent, by NusG (Fig. 2D; Hao et al. 2021a,b; Said et al. 2021).
Which model best describes RDTT is still debated, as there are pros and cons for each proposed mechanism (Hao et al. 2021b; Rashid and Berger 2023), while an alternative perspective incorporates “stand-by” complexes into the classical “catch-up” scheme (see Table 1 and below). One appealing feature of the “stand-by” model is that Rho is co-localized with the nascent transcript through its interaction with RNAP, providing a straightforward way to segregate genuine RNA targets from the bulk of cellular RNAs, which are RNAP-free (Hao et al. 2021a,b; Said et al. 2021). However, target discrimination or subcellular localization could also be achieved by other mechanisms, such as liquid–liquid phase separation (Nandana and Schrader 2021), protein filament polymerization (Mance et al. 2024), or topological constraints imposed on the nucleoid by various DNA- and RNA-binding proteins (Hustmyer and Landick 2024). For example, the Rho factor of Bacteroides thetaiotaomicron contains an unusually large intrinsically disordered region (IDR) inserted within its NTD (Fig. 1A), which promotes the formation of Rho condensates independently of RNAP presence (Krypotou et al. 2023). These Rho condensates are detectable in vivo by immunofluorescence, unless the IDR is deleted, highlighting their importance for the bacterium's fitness. Alternatively, Rho polymerization may regulate the spatial distribution of inactive versus active Rho factors, similar to the mechanism observed for other transcription factors (Mance et al. 2024). A recent work suggests that E. coli’s Rho can indeed oligomerize into long filaments under certain conditions, including cellular stresses (Wang et al. 2025). Further investigations using advanced super-resolution microscopy approaches will be necessary to determine the subcellular oligomeric states of Rho and whether all Rho particles, or only spatially defined subsets, actively participate in transcription termination.
Intriguingly, interactions that stabilize the “stand-by” pretermination complex (Fig. 2D) include Rho contacts to a lineage-specific insertion sequence in the β subunit of RNAP (Said et al. 2021; Simon et al. 2021). While some of these interactions are supported by genetic or biochemical evidence (Epshtein et al. 2010; Hao et al. 2021a; Said et al. 2021), it remained unclear whether “stand-by” pretermination complexes could form in species distant from E. coli (Simon et al. 2021). Recent cryo-EM reconstructions of the pretermination complex from Thermus thermophilus (Murayama et al. 2023) provide some insight. In this species, the Rho factor binds the TEC in a configuration that closely resembles the “catch-up” pretermination complex observed in E. coli (Fig. 2B). However, 3D classification of the cryo-EM particles revealed an alternative binding mode where the NTD face of the Rho hexamer contacts RNAP in a “stand-by”-like manner. Nonetheless, this configuration differs from the “stand-by” model as Rho adopts a closed-ring conformation while RNAP is in an apo-like rather than TEC-like state. Biochemical probing of the Rho-RNAP interfaces suggested that the “catch-up” complex is the genuine pretermination state, while the “stand-by”-like complex may be an off-pathway intermediate (Murayama et al. 2023).
Recent work by Hohng and colleagues provides key insights that may help reconcile the “catch-up” versus “stand-by” views. Using single-molecule fluorescence assays, these authors demonstrated that Rho could engage in both pathways in vitro (Song et al. 2022). Across all terminators tested, the “catch-up” RDTT pathway, leading to RNA release and subsequent one-dimensional sliding of RNAP on DNA, was the most frequently used and the fastest (Song et al. 2023). In contrast, the “stand-by” RDTT route was the slowest and, intriguingly, the only pathway in which termination efficiency positively correlated with the duration of RNAP pausing along DNA (Song et al. 2023). This finding, together with the inactive (open) conformation of the Rho hexamer in the “stand-by” complex (Fig. 2D), may reflect the time needed for the complex to transition into a productive state. This suggests that the “stand-by” complex could represent either a very early intermediate or an off-pathway state, preceding Rho's engagement in the active RDDT pathway. Such inactive Rho association with the TEC may explain the previously noted similarity between Rho and RNAP distributions along the E. coli genome, as observed in ChIP-chip analyses (Mooney et al. 2009). Notably, “catch-up” Rho-RNA complexes (Fig. 2A) are unlikely to be efficiently cross-linked to DNA and, thus, may go undetected in ChIP-chip experiments. These observations raise the possibility that two Rho recruitment pathways coexist: one in which Rho diffuses freely in solution before engaging the transcript, and another in which Rho is prebound to RNAP and subsequently transfers to the nascent RNA, ultimately triggering TEC inactivation and RDTT. Evidence supporting the coexistence of two distinct Rho recruitment pathways in vivo comes from examining how Rho and NusG alterations impact RDTT profiles at the genome-wide level (see section below).
RDTT IN GLOBAL TRANSCRIPTION MANAGEMENT
Transcriptome analyses comparing the RNA profiles of untreated cells with those of cells undergoing Rho inhibition or depletion have enabled the mapping of the RDTT landscape in key model bacteria. These studies have uncovered significant differences in how various bacteria rely on Rho to terminate transcription at the end of genes. For instance, while only ∼20% of E. coli genes carry Rho-dependent terminators at their 3′ ends (Peters et al. 2012), the vast majority of genes in Mycobacteria have their transcripts terminated by Rho (Botella et al. 2017; Ahmad et al. 2023). However, a common finding across all studied organisms is the central role played by RDTT in suppressing pervasive antisense transcription within coding regions (Nicolas et al. 2012; Peters et al. 2012; Leela et al. 2013; Mader et al. 2016; Raghunathan et al. 2018). This activity is thought to help prevent the formation of deleterious transcriptional R-loops upstream of RNAPs (Leela et al. 2013; Raghunathan et al. 2018; Pandiyan et al. 2024) and to facilitate the removal of stalled TECs, which can otherwise create conflicts with DNA replication and lead to DNA breaks (Washburn and Gottesman 2010; Dutta et al. 2011; Jain et al. 2019). One striking finding from early transcriptomic studies was that Rho, assisted by NusA and NusG, silences the expression of horizontally acquired A/T-rich regions in the E. coli genome (Cardinale et al. 2008). Among Rho-silenced targets are toxic prophage genes (Cardinale et al. 2008; Menouni et al. 2013; Hafeezunnisa et al. 2021; Baniulyte and Wade 2025), which partially accounts for the essentiality of NusG and Rho in E. coli (Cardinale et al. 2008). This elevated termination activity in A/T-rich regions was initially attributed to the presence of codons with a lower adaptation index, which cause translational stalling and thereby promote RDTT (see below) (Cardinale et al. 2008).
An alternative scenario emerged from the finding that Rho-mediated silencing sites coincide with regions bound by the nucleoid-structuring protein H-NS (Saxena and Gowrishankar 2011; Peters et al. 2012). H-NS, a highly abundant DNA-binding protein, plays a central role in silencing horizontally acquired genes (Lucchini et al. 2006; Navarre et al. 2006). It binds to A/T-rich DNA sequences, forming nucleoprotein filaments that block RNAP access to promoters over extended regions (Lucchini et al. 2006; Landick et al. 2015). This repression not only prevents transcription from regular promoters but also silences the multitude of spurious intragenic promoters that arise due to the high A/T content of these regions (Singh et al. 2014; Lamberte et al. 2017). The presence of Rho-dependent terminators within H-NS-bound regions suggests that H-NS-mediated repression of transcription initiation is incomplete and that Rho is required to terminate elongating transcripts that occasionally escape this control. This functional cooperation between H-NS and Rho was recently demonstrated in Salmonella, where certain rho mutations or NusG depletion result in the constitutive activation of H-NS-silenced pathogenicity islands (Bossi et al. 2019). In the case of Salmonella pathogenicity island 1 (SPI-1), readthrough transcripts originating several hundred base pairs upstream of the master regulator gene hilD, relieve H-NS-mediated repression without actually reaching the hilD promoter. Remarkably, repression is restored by inserting a strong gyrase site (SGS) within the intervening sequence (Figueroa-Bossi et al. 2024). This led to a model in which transcription destabilizes H-NS:DNA filaments at a distance via the generation of positive DNA supercoiling (Fig. 3). Drawing from in vitro evidence that H-NS can constrain negative supercoils (Tupper et al. 1994), the model proposes that H-NS filaments in vivo form on negatively supercoiled plectonemic DNA, with H-NS bridging the opposing arms of the plectoneme. Rotational diffusion of positive supercoils generated by neighboring transcription (Liu and Wang 1987; Wu et al. 1988) may cause the H-NS-bound plectoneme to “unroll,” disrupting H-NS bridges and leading to transient filament disassembly. After the passage of RNAP, the filament can rapidly reassemble, aided by the accumulation of negative supercoils behind the moving transcription complex (Fig. 3). This dynamic scenario aligns with ChIP data showing that substantial amounts of H-NS remain bound to SPI-1 even when the island is actively transcribed (Figueroa-Bossi et al. 2022, 2024).
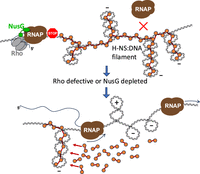
Model for the role of RDTT in H-NS-mediated gene silencing. H-NS:DNA filaments form on negatively supercoiled DNA and constrain negative supercoils (Tupper et al. 1994). H-NS represses transcription initiation from multiple promoters within the filament, including spurious intragenic promoters. Transcripts from promoters that escape repression are intercepted and terminated by Rho, aided by NusG (top). This function is crucial for maintaining the silenced state. If Rho or NusG is defective, RNAP invades the H-NS:DNA filament, generating positive DNA supercoiling as it translocates along the DNA (Wu et al. 1988). As positive and negative supercoils neutralize each other, the resulting loss of negative superhelicity destabilizes the H-NS:DNA complex, leading to transient filament disassembly (bottom). The filament rapidly reassembles after RNAP passage, driven by the surge of negative supercoils accumulating behind the elongating RNAP.
A major conclusion from these studies is that, in vivo, H-NS alone is insufficient to halt elongating transcription complexes and instead relies on Rho to perform this function. H-NS:DNA filaments have been shown to prolong RNAP pausing (Kotlajich et al. 2014; Boudreau et al. 2018), creating a window for NusG-mediated recruitment of Rho. This recruitment must occur early during elongation, before the surge of positive supercoiling leads to filament destabilization. Interestingly, a Rho mutant defective in Rut site recognition (Y80C) was found to retain full capacity to support H-NS-mediated gene silencing (Bossi et al. 2019). This observation suggests that RDTT at H-NS-repressed loci occurs predominantly via the “stand-by” pathway, which is expected to be less dependent on strong RNA binding and more influenced by the duration of RNAP pausing (Song et al. 2023).
Genome-wide studies in E. coli have hinted at the presence of Rho-dependent terminators in the 5′UTRs of various genes (Peters et al. 2009; Sedlyarova et al. 2016, 2017; Adams et al. 2021), implying that Rho could play a significant role in the biogenesis of noncoding RNAs and/or in transcription attenuation mechanisms. This hypothesis is further supported by a recent map of putative Rut sites generated using Helicase-SELEX (Delaleau et al. 2022), a mapping approach that is not biased by the extensive posttranscriptional 3′-end processing of Rho-dependent transcripts (Dar and Sorek 2018). Alongside the frequent occurrence of RDTT in horizontally acquired portions of the E. coli genome, these observations suggest that RDTT could serve as a major adaptive mechanism, enhancing bacterial fitness to environmental fluctuations. Genome-wide studies of other bacteria have since revealed that these features are not exclusive to E. coli (D'Halluin et al. 2023; Petroni et al. 2023). Moreover, individual cases of mechanisms of conditional Rho-dependent regulation have been identified across various species (Table 2). In the following section, we briefly present these mechanisms and highlight the most recent discoveries. For a more comprehensive discussion of previously identified mechanisms, readers may refer to other recent reviews (Kriner et al. 2016; Bastet et al. 2018; Roberts 2019; Turnbough 2019; Bossi et al. 2020).
Genes and operons regulated by Rho in a conditional manner
RDTT IN THE REGULATION OF SPECIFIC GENOMIC LOCI
Mechanisms of conditional Rho activity
Conditional RDTT requires the direct or indirect regulation of Rho activity by an inducer at one or more stages of the RDTT pathway. These stages include Rho loading onto RNA, catalytic isomerization, translocation along RNA, formation of the pretermination complex with RNAP, and Rho-mediated disassembly of the transcription complex.
Among these possibilities, the regulation of Rho loading onto RNA is perhaps the best documented. In the bacteria where transcription and translation are coupled (Woodgate and Zenkin 2023), actively translating ribosomes protect nascent mRNAs by physically obstructing Rho's access to intragenic Rut sites (Fig. 4A). Factors that uncouple translation from transcription expose intragenic Rut sites, leading to RDTT-induced silencing of the corresponding gene(s) and downstream genes of the same operon. This process is called transcriptional polarity (Richardson et al. 1975; Ratner 1976; Wek et al. 1987). Ribosome-targeting antibiotics (Zhu et al. 2019) or the depletion of translation elongation factors (Elgamal et al. 2016) promote ribosome stalling, thereby enhancing Rho-mediated transcriptional polarity. Similarly, conditions that impair or reduce the frequency of translation initiation also induce polarity effects (de Smit et al. 2009).
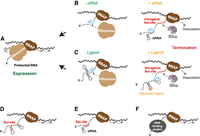
Regulation of RDTT by blocking of the Rut site. (A) Translation–transcription coupling protects intragenic Rut sites from Rho action. (B) Binding of a sRNA to the mRNA leader at/near the RBS prevents translation and unmasks intragenic Rut site(s) leading to RDTT. (C) Restructuring of the mRNA leader upon ligand binding to its cognate riboswitch masks the RBS, allowing RDTT from intragenic Rut sites. Inhibition of RDTT may also stem from sequestering of the Rut site in (D) a secondary RNA structure, or (E) a sRNA–mRNA duplex, or (F) from blocking of the Rut site by a RNA-binding protein (hypothetical case not discussed in main text).
This principle is widely exploited in conditional silencing mechanisms. For instance, sRNA binding to 5′ UTRs (Bossi et al. 2012) or riboswitch-induced restructuring of mRNA leader sequences (Bastet et al. 2017) sequester the ribosome binding site (RBS) within an RNA duplex, preventing ribosome association and promoting Rho-dependent polarity (Fig. 4B,C; see examples in Table 2). In E. coli, RNAP pausing near the RBS and start codon in the 5′UTR (Larson et al. 2014) likely facilitates sRNA binding or mRNA leader restructuring before the ribosome associates with the nascent transcript (Chauvier et al. 2019, 2021). A similar attenuation system was discovered recently in Vibrio cholerae where the sRNA inducer, OppZ, is the last product of its own operon, oppABCDFZ (Table 2; Hoyos et al. 2020). OppZ binding to the intergenic region between oppA and oppB blocks oppB translation exposing an intragenic Rho-dependent terminator that silences downstream genes via a negative feedback loop (Hoyos et al. 2020). In all these cases, the combination of transcriptional and translational repression ensures that the inducer triggers robust gene/operon silencing.
Riboswitches and sRNAs can also regulate RDTT more directly by sequestering Rut sites within secondary RNA structures or RNA duplexes, preventing Rho from loading onto the nascent RNA (Fig. 4D,E). One likely case leads to transcription antitermination and the stimulation of rpoS expression upon binding of sRNA DsrA, ArcZ, or RprA to the 5′UTR of rpoS (Sedlyarova et al. 2016) at a site overlapping with a putative Rut site mapped by helicase-SELEX (Delaleau et al. 2022). While this steric interference mechanism has been proposed in several other instances (Table 2), the impact of the inducer on Rho's affinity for its RNA target has rarely been tested. However, a recent study demonstrated that temperature specifically modulates Rho binding to the mRNA leader sequences of genes and operons involved in the E. coli’s cold shock response, including cspA, cspB, cspG, and nsrR-rnr-yjfHI (Delaleau et al. 2024). At 37°C, Rho efficiently binds these mRNA leaders, contributing to the repression of cold shock gene expression (Fig. 5, top). Upon cold shock, the mRNA leaders adopt “closed” conformations that are less favorable to Rho binding while exposing RBS sequences, thereby preventing RDTT and optimizing the translation of the cold shock proteins (Fig. 5, middle). During cold acclimation, the cold shock proteins provide negative feedback regulation by binding the mRNA leaders once they reach a sufficient concentration, switching them back to conformations more favorable for RDTT (Fig. 5, bottom; Delaleau et al. 2024). This discovery challenges the long-held view that the cold shock response is primarily a posttranscriptional program (Zhang et al. 2018). Instead, it highlights RDTT as an integral component of a complex regulatory circuit that ensures precise control of gene expression (Fig. 5; Delaleau et al. 2024).
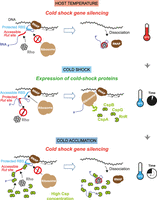
Diagram depicting the regulation of cold shock genes during the cold shock. At 37°C (top), the mRNA leaders adopt OFF conformations that favor RDTT and disfavor translation, inhibiting expression of the cold shock proteins (csp). Temperature-dependent restructuring of the mRNA leaders upon cold shock (middle) has an opposite effect, masking the Rut sites and unmasking the RBSs, to allow csp expression. During cold acclimation (bottom), the concentrations of csp proteins increase until they become sufficient for the csp proteins to bind their mRNA leaders and promote their restructuring into OFF conformations favoring RDTT and disfavoring translation.
Translation of a small upstream open reading frame (uORF) provides another mechanism for conditional RDTT regulation (Table 2). A classic example is the regulation of the tryptophanase operon (tnaCAB) via translation of the tnaC uORF (Fig. 6A; Stewart et al. 1986). At high L-tryptophan concentration, tryptophan invades the ribosomal exit tunnel, inducing ribosome stalling at the tnaC termination codon and preventing cleavage and release of the tnaC leader peptide (Gong et al. 2001; van der Stel et al. 2021). The stalled ribosome then blocks Rho from accessing an adjacent Rut site in the tnaC–tnaA intergenic region, allowing expression of the downstream tnaA (tryptophanase) and tnaB (tryptophan transporter) genes (Stewart et al. 1986; Gish and Yanofsky 1995). A similar uORF-dependent attenuation-by-RDTT mechanism regulates the expression of the ornithine decarboxylase gene (speF) in E. coli and Salmonella in response to ornithine availability (Table 2; Ben-Zvi et al. 2019; Herrero Del Valle et al. 2020). However, other instances of uORF-dependent attenuation-by-RDTT deviate from this paradigm and instead activate rather than inhibit RDTT in response to the inducer (Table 2). In the cruR-bfrG-bp2921 operon of Bordetella pertussis, high copper ion concentrations trigger ribosome release from the cruR uORF where it otherwise masks an intragenic Rut site (Fig. 6B; Roy et al. 2022). Copper thus exposes the Rut site within cruR, activating RDTT-mediated attenuation of the downstream bfrG gene, which encodes a putative Cu importer, likely contributing to Cu homeostasis. The molecular mechanisms underlying ribosome stalling by the CruR peptide and ribosome release by Cu ions remain unclear but appear to involve two conserved motifs in the peptide: CXXC (middle) and RAPP (C-terminal) (Roy et al. 2022).
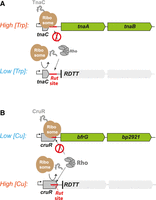
uORF-dependent attenuation-by-RDTT. (A) Diagram depicting the regulation of the E. coli tnaCAB operon by tryptophan. At high concentration, tryptophan prevents release of the ribosome from the termination codon, shielding the downstream Rut site and allowing expression of tnaAB. (B) Diagram depicting the regulation of the Bordetella pertussis tnaCAB cruR-bfrG-bp2921 operon by Cu ions. The ribosome is stalled on the cruR stop codon in absence of Cu ions, shielding the intragenic Rut site and allowing expression of the putative Cu importer bfrG. In the presence of Cu ions, the ribosome is efficiently released from the cruR stop codon, allowing RDTT and silencing of bfrG (Roy et al. 2022).
The mechanisms by which uORFs govern Rho-dependent attenuation in the corA, mgtA, and mgtC genes, involved in magnesium homeostasis in Salmonella (Table 2), have been studied extensively (for review, see Kriner et al. 2016; Bossi et al. 2020). In these cases, Mg2+ concentration modulates uORF translation efficiency, which in turn influences how the downstream mRNA leader sequence folds during transcription (Kriner and Groisman 2015, 2017; Gall et al. 2016). Under low Mg2+ conditions, ribosome stalling in the uORF promotes the formation of an RDTT-resistant mRNA conformation, enabling the expression of downstream Mg2+ importer genes. Conversely, high Mg2+ levels enhance uORF translation and drive the mRNA leader into an RDTT-prone conformation promoting transcriptional attenuation. For the mgtC attenuator, however, mRNA leader restructuring does not directly regulate Rho access to the Rut site. Instead, it controls a Rho-antagonizing RNA element (RARE) motif in the nascent transcript, which can trap the Rho–RNA complex in an inactive state (Sevostyanova and Groisman 2015). When uORF translation is inefficient, RARE remains single-stranded and inhibits Rho, allowing mgtCBR expression. In contrast, when uORF translation is efficient, RARE is sequestered in a stem–loop structure, preventing its inhibitory effect and facilitating RDTT-mediated attenuation of mgtCBR. Given the growing number of newly identified uORFs (Orr et al. 2020; Adams et al. 2021), particularly in taxa such as Mycobacteria where RDTT also plays a critical role (Botella et al. 2017; Ahmad et al. 2023), these known cases of uORF-dependent attenuation-by-RDTT may represent just the tip of the iceberg.
RDTT can also be regulated stochastically, enabling bimodal expression of fitness or virulence factors while allowing bacterial pathogens to evade host immune responses through phase variation and phenotypic heterogeneity (Bidnenko and Bidnenko 2018). Notable examples include invertible DNA elements upstream of fimbriae operons in E. coli (Hinde et al. 2005) and flagella operons in Clostridioides difficile (Warren Norris et al. 2024). These invertible elements encode Rut site(s) in only one orientation, enabling the operons to switch between ON and OFF states depending on the stochastic activity of recombinases (Table 2). Additionally, stochastic readthrough of Rho-dependent terminators provides another mechanism for bimodal gene expression. This has been observed in Salmonella, where it regulates virulence genes (Figueroa-Bossi et al. 2022), and in B. subtilis, where it influences motility and cell differentiation genes (Bidnenko and Bidnenko 2018).
Protein factors that contribute to regulate RDTT
Besides H-NS, whose functional interaction with Rho was discussed earlier, various proteins regulate RDTT at specific terminators or under particular conditions such as stress responses or the hijacking of bacterial transcription by bacteriophages (Table 3). These protein factors and their roles in RDTT have been studied primarily for E. coli and may not be universally conserved across bacteria (Reay et al. 2004; Kalarickal et al. 2010).
Proteins that regulate RDTT
In E. coli, two major Rho partners are transcription elongation factors NusA and NusG (Table 3). Both factors can bind Rho and RNAP within pretermination complexes (see above and Fig. 2B,D; Hao et al. 2021a; Said et al. 2021; Molodtsov et al. 2023) and regulate RDTT at specific subsets of terminators in E. coli (Peters et al. 2012; Qayyum et al. 2016) and Salmonella (Carlomagno and Nappo 2003; Bossi et al. 2019). Notably, NusG enhances RDTT at suboptimal Rut sequences with little or no C>G bias (Peters et al. 2012; Delaleau et al. 2022). In contrast, NusA inhibits Rho's ATPase activity (Schmidt and Chamberlin 1984) and typically shifts the termination window downstream while reducing RDTT efficiency in vitro (Lau and Roberts 1985; Chen and Richardson 1987; Sigmund and Morgan 1988; Burns et al. 1998; Qayyum et al. 2016; Simon et al. 2021; Zhu et al. 2022), although a stimulatory effect has been reported in a few cases (Kainz and Gourse 1998; Hao et al. 2021a). Reasons for this discrepancy remain unclear.
NusA and NusG also participate in multipartite antitermination complexes that protect the transcription of ribosomal rrn operons (which are untranslated) and lambdoid phage early genes from Rho action (Table 3; Santangelo and Artsimovitch 2011; Wang and Artsimovitch 2020). These rrn and λN antitermination complexes share several core components (NusA/G/B/E) but also include distinct factors (S4 and SuhB vs. λN) that collectively transform the TEC into a highly processive, RDTT-resistant form (Fig. 7A; Krupp et al. 2019; Huang et al. 2020). A key step in the assembly of these complexes is the recruitment of NusB/E by a boxA sequence in the nascent RNA (Fig. 7B; Nodwell and Greenblatt 1993).
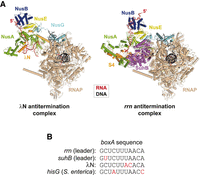
Cryo-EM reconstructions of the λN (PDB 6GOV) and rnn (PDB 6TQO) antitermination complexes show how the antitermination factors bind RNAP and the nascent transcript to shield them efficiently from Rho action (compare with Fig. 2B,D). The complexes also sequester NusG-CTD (by NusE) and prevent RNAP pausing or backtracking (Krupp et al. 2019; Huang et al. 2020).
Interestingly, however, boxA recognition by Nus factors can sometimes promote rather than inhibit RDTT. For example, recruitment of a NusB/A complex by an intragenic boxA sequence enhances RDTT at a suboptimal terminator in the hisG gene of Salmonella (Carlomagno and Nappo 2003). Similarly, a boxA sequence in the 5′ leader of the suhB mRNA in E. coli and Salmonella (Fig. 7B) cooperates with NusB/E to trigger RDTT within the suhB coding sequence (Baniulyte et al. 2017). Formation of a boxA-NusB/E complex near the RBS appears to inhibit suhB translation through a cryptic, nonsteric occlusion mechanism, thereby promoting Rho-dependent polarity. Computational analyses suggest that boxA-mediated regulation of RDTT is phylogenetically widespread, extending beyond these few reported cases (Baniulyte et al. 2017).
Several antitermination proteins can independently shield the TEC from RDTT using distinct molecular strategies. For instance, the NusG paralog RfaH recognizes 12 nt-long ops (operon polarity suppressor) nontemplate DNA sequences located in the 5′UTRs of a select group of long, xenogeneic operons encoding fitness and virulence factors in E. coli, Salmonella, and possibly other Enterobacteriaceae (Wang and Artsimovitch 2020). The ops sequence induces hyper-pausing of RNAP, which facilitates RfaH recruitment and its structural transition from an auto-inhibited to an active conformation. Once active, RfaH binds both RNAP and the ribosome, reinforcing transcription–translation coupling throughout the coding regions of the ops-bearing operons (Molodtsov et al. 2024; Zuber et al. 2024). Lambdoid phages use Q antitermination proteins, which recognize qut (Q utilization) DNA sequences near the promoter to associate with σ-dependent paused TECs. Once loaded, these proteins remain persistently associated with the TEC, ensuring the uninterrupted transcription of the phage late genes (Table 3; Roberts et al. 2008). The Q protein of phage 21 (Q21) alone forms a strong antitermination complex with the TEC (Shi et al. 2019; Yin et al. 2019), whereas the nonhomologous Qλ and Q82 proteins require NusA as a cofactor (Yang et al. 1989; Shankar et al. 2007; Yin et al. 2022). Nonetheless, structural analyses of Q21 (Shi et al. 2019; Yin et al. 2019) and Qλ (Yin et al. 2022) antitermination complexes suggest a shared mechanism: the Q protein forms a torus-like structure that extends and narrows the RNA exit channel of RNAP. This steric alteration prevents the formation of pause or termination-inducing RNA hairpins within the channel and likely interferes with Rho-mediated TEC disruption (Yang and Roberts 1989; Yarnell and Roberts 1992; Shankar et al. 2007), though this hypothesis requires further validation. Additionally, the UvrABD complex, which facilitates transcription-coupled repair (TCR), uses RNAP as a binding platform (Bharati et al. 2022). Indirect evidence suggests that UvrABD competes with Rho and NusG for the same RNAP binding surface, thereby promoting antitermination during TCR (Martinez et al. 2022).
Several antitermination proteins inhibit Rho directly rather than targeting RNAP (Table 3). One example is the Psu protein from phage P4, which supports late gene transcription in helper phage P2 by preventing Rho-dependent polarity (Linderoth and Calendar 1991). Recent cryo-EM reconstructions of the Rho–Psu complex suggest a mechanism in which Psu dimers bridge and lock two Rho hexamers in tail-to-tail, inactive open-ring conformation (Fig. 8A; Gjorgjevikj et al. 2025). Interestingly, the helical pitch of the Rho rings in this complex is higher than in crystal structures of Psu-free, open Rho hexamers (Skordalakes and Berger 2003; Skordalakes et al. 2005), allowing for the oligomeric expansion of Rho rings in a subset of the cryo-EM particles (Fig. 8A; Gjorgjevikj et al. 2025). This mechanism contrasts with that of the Sm-like protein Rof (aka YaeO), an endogenous Rho inhibitor that protects E. coli from oxidative stress (Kawamura et al. 2005) and is required for virulence and host invasion in Salmonella (Zhang et al. 2024). Rof molecules bind to the protomer interfaces of the Rho hexamer (one Rof per interface), collectively locking Rho in an inactive, open Rho ring configuration (Fig. 8B; Said et al. 2024; Zhang et al. 2024). Unlike Psu, binding of Rof does not appear to extend the helical pitch of the Rho ring. Additionally, bound Rof molecules partially occlude Rho's PBS, impairing its ability to interact with RNA or NusA/G-containing TECs in vitro (Said et al. 2024). Another recently discovered Rho inhibitor is the stress response kinase YihE (Wang et al. 2022). YihE expression is stimulated during the conjugative pilus expression (Cpx) stress response, where it helps protect E. coli from oxidative damage and lipid peroxidation. A low-resolution crystal structure of the Rho–YihE complex suggests that a single YihE molecule binds the PBS of the leftmost Rho subunit at the gap of an open Rho hexamer (Wang et al. 2022). However, whether this interaction inactivates Rho by preventing ring closure or by interfering with other steps of the RDTT pathway remains unclear. Notably, the structural data for the Rho–YihE complex have not been deposited by the authors (only that for a structure of Rho alone under PDB code 7X2R), which raises some doubts about the findings.
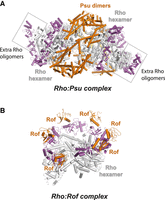
Cryo-EM reconstructions of the (A) Rho–Psu complex (PDB 9GCT) and (B) Rho–Rof complex (PDB 8W8D) show how the two Rho inhibitors use distinct strategies to keep Rho into an inactive configuration. Rho's NTD residues are colored in magenta.
Another intriguing endogenous Rho interactor and inhibitor is the RNA chaperone Hfq (Rabhi et al. 2011a), which stabilizes sRNA:mRNA complexes during sRNA-mediated gene silencing (Katsuya-Gaviria et al. 2022). Hfq forms a relatively stable binary complex with Rho (Kd ∼40 nM) but effective inhibition of Rho activity requires the simultaneous interaction of both proteins with their RNA target (Rabhi et al. 2011a). While Hfq's antitermination activity has been observed in E. coli using a plasmid reporter system carrying the prototypical λtR1 terminator (Rabhi et al. 2011a), its physiological antitermination targets remain to be firmly identified. A strong candidate is the Rho-dependent terminator in the 5′UTR of the rpoS stress response gene, which is inactivated when sRNA DsrA, ArcZ, or RprA binds to the rpoS mRNA leader sequence in an Hfq-dependent manner (Sedlyarova et al. 2016; Nadiras et al. 2018b). Virulence factors subject to opposing regulation by Rho and Hfq are also compelling possibilities (Rabhi et al. 2011a). Finally, a recent study reporting Hfq's association with hundreds of nascent transcripts in Pseudomonas aeruginosa further supports its potential role as a Rho interactor (Kambara et al. 2018).
Other Rho interactors have been identified by mass spectrometry (Table 3; Butland et al. 2005), but their direct interactions with Rho or their effects on RDTT remain unverified. Additionally, Rho has been identified via mass spectrometry in a large c-di-GMP-dependent protein complex upon overexpression and pull-down of the oxygen sensors DosC and DosP (Tuckerman et al. 2011). Intriguingly, the complex also contained the RNA degradosome components PNPase, enolase and RNase E, providing the first direct evidence of a condition-dependent link between Rho and the RNA degradosome in E. coli. In contrast, in Rhodobacter capsulatus, Rho appears to associate more stably with the RNA degradosome (Jager et al. 2001), though its proportion within the complex fluctuates depending on oxygen availability during growth (Jager et al. 2004).
Accessory nucleic acid sequences that regulate RDTT
Accessory DNA/RNA sequences or motifs play a crucial role in regulating RDTT, ensuring that this regulation remains confined to specific regions of the bacterial or phage genome. For example, the presence of a qut or ops sequence in the DNA template is essential for recruiting the antitermination factors Q and RfaH, respectively (see section above and Table 3). Accessory RNA sequences influence co-transcriptional RNA folding and modulate Rho access to Rut sites (Fig. 4B–D) or, in the case of the RARE sequence, drive the catalytic isomerization of the Rho–RNA complex (Sevostyanova and Groisman 2015). BoxA sequences within the nascent transcript recruit Nus factors either stimulating or inhibiting RDTT, depending on the genomic context (see section above). Similarly, a boxB sequence aids in assembling the λN antitermination complex by recruiting the λN protein (Vieu and Rahmouni 2004), whereas comparable promoter-proximal boxB sequences play a much less significant role in rrn antitermination in E. coli (Warner et al. 2023).
Regulation of RDTT can also occur through the binding of accessory RNA sequences to RNAP. For example, promoter-proximal sequences known as put sites, encoded by the phage HK022 genome interact with RNAP to transform the TEC into a termination-resistant form (King and Weisberg 2003). Cryo-EM reconstructions of putRNA–TEC complexes suggest that this interaction prevents “swiveling,” a conformational shift in RNAP that facilitates transcriptional pausing (Hwang et al. 2022). However, whether this anti-pausing activity fully accounts for putRNA-mediated inhibition of RDTT (King and Weisberg 2003) remains unclear. More recently, natural RNAP-binding aptamers (RAPs) have been identified in E. coli through genomic SELEX, including a subset of inhibitory RAPs (iRAPs) that promote RDTT (Sedlyarova et al. 2017). The authors have proposed that iRAP binding to RNAP hampers the progression of the trailing ribosome, uncoupling translation from transcription thereby eliciting Rho-dependent polarity (Sedlyarova et al. 2017). However, the subsequent discovery of iRAPs in noncoding, antisense regions (Magan et al. 2019) suggests that they may trigger RDTT through a different yet-undetermined mechanism. To date, the physiological roles of RAPs and iRAPs, as well as their modes of engagement—whether constitutive or conditional—remain largely unknown.
TWEAKING RHO-DEPENDENT ATTENUATION FOR SYNTHETIC BIOLOGY
Despite the key role of RDTT in the regulation of bacterial gene expression, its application in synthetic biology has been limited. However, certain rho mutants evolved under specific stress conditions, such as heat stress (Kishimoto et al. 2010; Deatherage et al. 2017), ethanol exposure (Freddolino et al. 2012; Haft et al. 2014), or carbon source limitation (Conrad et al. 2009; Worthan et al. 2024), may hold potential for bioproduction processes or other biotechnological applications. Notably the rho R109H mutation transforms Rho into a pH-dependent enzyme within the physiological pH range (Worthan et al. 2024), offering a unique opportunity for pH-responsive genetic control.
The tryptophan-responsive tnaC attenuator of the tnaCAB operon (Fig. 6A) is a rare example of Rho-dependent regulatory element integrated into synthetic biological circuits. Its dynamic response (Wang et al. 2020a) and orthogonal control can be fine-tuned, with the latter enhanced in nusB-depleted strains, where cross-regulation by other amino acids and natural metabolites is reduced (Sherman et al. 2021). The tnaC attenuator has been used to detect and enhance L-tryptophan production and to optimize metabolic fluxes where L-tryptophan serves as an intermediate (Fang et al. 2016; Liu et al. 2017; Chen et al. 2019; Wang et al. 2020b; Gwon et al. 2021). For example, in a synthetic E. coli co-culture platform designed for tryptamine bioproduction, the tetracycline resistance gene tetA was placed under tnaC control (Wang et al. 2020b). The tnaC-tetA sensor selectively promotes the growth of the subpopulation of cells that produce high amounts of L-tryptophan, the tryptamine precursor, in the presence of tetracycline. In another application, an amber UAG stop codon has been introduced into tnaC to tailor its sensing capability to an unnatural amino acid and its corresponding, synthetic aminoacyl-tRNA synthetase (Liu et al. 2011). Similarly, engineering the 5′UTR upstream of tnaC enables the attenuator to respond to RNA duplex formation with a complementary RNA, modulating RBS accessibility and downstream transcriptional output (Mihailovic et al. 2018).
The growing repertoire of natural Rho-dependent attenuators (Table 2) provides new opportunities for developing synthetic regulatory devices responsive to diverse signal inputs. Another promising avenue is the de novo engineering of synthetic RDTT attenuators. Certain synthetic aptamers and riboswitches contain C-rich sequences that can be modified into Rut-like switches enabling ligand-dependent Rho recruitment (Boudvillain M, Delaleau M, Eveno E, et al., unpubl.) in a way reminiscent of the CsrA-responsive pgaA attenuator (Table 2; Figueroa-Bossi et al. 2014). Additionally, switchable Rut motifs can be evolved de novo using a modified Helicase-SELEX approach. By applying alternating rounds of selection and counter-selection—depending on the presence and absence of a target ligand—this strategy yields RNA sequences that regulate Rho helicase activity in a ligand-dependent manner (Boudvillain et al. 2021). As a proof or principle, this approach has been successfully used to evolve synthetic serotonin-responsive Rut sequences capable of modulating Rho activity both in vitro and in vivo (Delaleau et al. 2022).
CONCLUSIONS AND OPEN QUESTIONS
Paraphrasing the title of a previous review of RDTT (Banerjee et al. 2006), and despite significant advances since its publication, many questions remain regarding the mechanisms and physiological roles of Rho. While both in vitro (Fig. 2; Hao et al. 2021a; Said et al. 2021; Song et al. 2022, 2023; Molodtsov et al. 2023; Murayama et al. 2023) and in vivo (Shashni et al. 2014; Bossi et al. 2019) studies support the existence of parallel RDTT pathways, their specific mechanisms and biological relevance require further investigation. Is there a pathway dominant under all conditions, or do different environmental or cellular contexts favor distinct RDTT mechanisms? Is the RDTT pathway for a given terminator permanently defined, or can it change dynamically in response to variable or inducible factor(s)? More broadly, are parallel RDTT pathways a common feature across Rho-bearing species or do they represent exceptions rather than the rule?
RDTT is traditionally regarded as the primary mechanism for monitoring translation–transcription coupling. However, recent findings suggest that this coupling may be less prevalent than previously assumed (Woodgate and Zenkin 2023) or that mRNA can be significantly “looped out” between RNAP and the ribosome, making it accessible to Rho (Qureshi and Duss 2024). If this is the case, how is spurious RDTT prevented? Is Rho activity spatially confined to specific subcellular or nucleoid regions? Could some RAPs function similarly to put sequences, shielding RNAP from Rho-dependent termination? Furthermore, how is the action of numerous modulators of RDTT (Table 3) coordinated to ensure a precise spatiotemporal regulation of Rho?
Recent findings suggest that E. coli Rho can undergo reversible inactivation through the formation of filamentous Rho hyper-oligomers, particularly under stress conditions that disrupt translation–transcription coupling such as exposure to translation inhibitors (Wang et al. 2025). However, do the Rho-containing cellular aggregates observed in the presence of these inhibitors truly represent inactive Rho filaments, given that similar inhibitory conditions have been shown to stimulate rather than suppress RDTT (Zhu et al. 2019)? Alternatively, could oligomeric Rho filaments serve other functions, such as facilitating the subcellular clustering of Rho activity, a phenomenon recently observed for other filament-forming transcription factors (Mance et al. 2024)?
Many Rho-regulated processes have now been identified and recognized to be critical for bacterial adaptation and fitness (Table 2). However, our understanding remains heavily biased toward the E. coli model. While RDTT continues to be intensely studied in this organism, research is expanding to uncover Rho's functions and mechanisms across the bacterial kingdom. Exciting discoveries suggest that RDTT can take diverse forms in phylogenetically distant bacteria, often influenced by taxon-specific variations in the Rho polypeptide (D'Heygere et al. 2015; Yuan and Hochschild 2017; Simon et al. 2021; Krypotou et al. 2023; Warren Norris et al. 2024). These variations extend to the regulation of Rho activity, which can occur through mechanisms such as liquid–liquid phase separation (Krypotou et al. 2023), β-amyloid aggregation (Yuan and Hochschild 2017), interaction with endogenous inhibitors (Zhang et al. 2024), RNA oxidation (Han et al. 2022), or growth-dependent fluctuations in intracellular Rho levels (Italiani et al. 2002; Silva et al. 2019; Bidnenko et al. 2024). What other regulatory surprises will emerge as RDTT is studied across the bacteria diversity? How does Rho function in extremophiles? Beyond bacterial fitness and adaptation, what roles does RDTT play in bacterial development programs or microbial community interactions?
Sophisticated experimental tools will be essential to tackle these timely questions. Genome-wide mapping of putative Rut sites is now feasible with Helicase-SELEX (Delaleau et al. 2022), which has already led to new discoveries such as Rho's involvement in the cold shock response of E. coli and Salmonella (Delaleau et al. 2024). Cryo-EM will likely continue to provide valuable structural insights into RDTT, potentially extending to cellular ultrastructure imaging through cryo-electron tomography in the near future. Additionally, super-resolution fluorescence microscopy approaches hold great promise for tracking single Rho particles—and their potential complexes with RNAP or Nus factors—in real time, assessing their subcellular distributions and velocities, and probing RDTT mechanisms in live cells. While super-resolution live-cell imaging of RNAP and NusG has been achieved (El Sayyed et al. 2024), extending this approach to Rho will require the development of an effective nondisruptive method for fluorescently labeling intracellular Rho molecules.
ACKNOWLEDGMENTS
This work was undertaken in partial fulfillment of the requirements for the doctoral degree of T.D.D. and was supported by grants from the French Agence Nationale de la Recherche to M.B. (ANR-19-CE44-0009-01 and ANR-22-CE44-0017). J.C.B. is a recipient of a PhD fellowship from the Université d'Orléans. L.B. and N.F.-B. acknowledge the support of the French National Research Council (CNRS).
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080486.125.
-
Freely available online through the RNA Open Access option.
This article, published in RNA, is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵








