
Structures of RNA phosphotransferase Tpt1 reveal distinct binding modes for an RNA 2′-PO4 splice junction versus a 5′-PO4 mononucleotide
- 1Molecular Biology Program, Memorial Sloan Kettering Cancer Center, New York, New York 10065, USA
- 2Chemistry Department, McGill University, Montreal, Quebec, Canada H3A0B8
- Corresponding author: shumans{at}mskcc.org
-
Handling editor: Eric Phizicky
Abstract
Tpt1 is a widely distributed enzyme that removes an internal RNA 2′-phosphate by transfer to NAD+, via a two-step reaction in which: (i) the RNA 2′-PO4 attacks NAD+ to form an RNA-2′-phospho-(ADP-ribose) intermediate and expel nicotinamide; and (ii) the ADP-ribose O2″ attacks the RNA 2′-phosphodiester to form 2′-OH RNA and ADP-ribose-1″,2″-cyclic phosphate products. Tpt1 can also execute a single-step ADP-ribosyltransferase reaction at a 5′-monophosphate nucleic acid terminus that installs a 5′-phospho-ADP-ribose cap structure. Here we present crystal structures of Tpt1 bound to an RNA containing an internal 2′-PO4 mark (the substrate for the canonical Tpt1 pathway) and in a complex with 5′-AMP. We find that Tpt1 has distinct binding modes, whereby the RNA 2′-PO4 and the AMP 5′-PO4 are engaged by the same set of active site amino acids, but the 2′-PO4 nucleoside and the 5′-nucleoside occupy different sites on the enzyme.
Keywords
INTRODUCTION
The enzyme Tpt1—an NAD+-dependent RNA 2′-phosphotransferase—is widely prevalent in bacterial, archaeal, and eukaryal taxa (Spinelli et al. 1998). Tpt1 was initially identified as an essential agent of fungal transfer RNA (tRNA) splicing that removes an internal RNA 2′-phosphate at the splice junction formed by fungal tRNA ligase (McCraith and Phizicky 1990, 1991; Culver et al. 1997). Tpt1 also removes the internal 2′-phosphate mark installed by Ark1 kinase at a V-loop uridine of archaeal tRNAs (Ohira et al. 2022). The Tpt1 mechanism entails two unique chemical steps (Fig. 1A). First, NAD+ reacts with the RNA 2′-phosphate to expel nicotinamide and generate a 2′-phospho-ADP-ribosylated RNA intermediate. Then, transesterification of the ADP-ribose 2″-OH to the tRNA 2′-phosphate displaces the 2′-OH RNA product and generates ADP-ribose-1″,2″-cyclic phosphate (McCraith and Phizicky 1991; Culver et al. 1993; Spinelli et al. 1999; Steiger et al. 2005; Munir et al. 2018a; Dantuluri et al. 2020; Jacewicz et al. 2023).
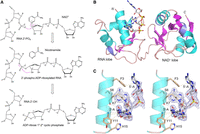
Tpt1 reaction mechanism and structure of PhoTpt1 in complex with 2′-PO4 RNA. (A) The Tpt1 reaction pathway comprises the two chemical steps shown in which: (i) the RNA 2′-PO4 reacts with NAD+ to expel nicotinamide and form a 2′-phospho-ADP-ribosylated RNA intermediate; and (ii) transesterification of the ADP-ribose 2″-OH to the RNA 2′-PO4 displaces the RNA 2′-OH and generates ADP-ribose-1″,2″-cyclic phosphate. (B) Tertiary structure of PhoTpt1, depicted as a cartoon model with magenta β-strands, cyan α-helices, and blue 310 helices. The ApA2′PpA substrate (stick model with gray carbons and yellow phosphorus atoms) occupies the RNA lobe. (C) Stereo view of the ApA2′PpA substrate overlaid with a simulated annealing omit density map (blue mesh contoured at 1 σ), highlighting its atomic interactions with amino acids in the N-terminal α1 helix of PhoTpt1 (depicted as stick models with beige carbons). Hydrogen bonds are denoted by black dashed lines. Van der Waals contacts are depicted as beige dashed lines.
Tpt1 proteins, typically ranging in size from ∼180 to 230 aa, consist of an N-terminal RNA lobe and a C-terminal NAD+ lobe separated by a flexible protease-sensitive hinge (Kato-Murayama et al. 2005; Sawaya et al. 2005; Banerjee et al. 2019; Alphonse et al. 2021). Alanine scanning mutagenesis of fungal and bacterial Tpt1 proteins has defined a conserved Arg–His–Arg–Arg catalytic tetrad essential for Tpt1 activity in vivo and in vitro (Sawaya et al. 2005; Munir et al. 2018a). Insights into the basis for 2′-PO4 recognition, NAD+ binding, and catalysis of the chemical steps emerged from a series of crystal structures of bacterial, archaeal, and mammalian Tpt1 enzymes captured at several discrete steps along the reaction pathway, including: (i) binary complexes with substrate NAD+ or product derivative ADP-ribose-1″-PO4 in the NAD+ lobe; (ii) 2′-terminal-phosphate-containing ligands 2′,5′-ADP and NADP+ in the RNA lobe; and (iii) a ternary product complex with ADP-ribose-2″-PO4 in the NAD+ lobe and a 2′-OH RNA in the RNA lobe (Banerjee et al. 2019; Jacewicz et al. 2023; Yang et al. 2023). These structures highlighted how flipping of the 2′-PO4 nucleobase into a Tpt1 surface crevice and atomic contacts to the vicinal 3′-PO4 and 2′-PO4 groups might ensure recognition of the splice junction.
Here we build on this theme, and gain further insights into catalysis, by solving the structure of an archaeal Tpt1 enzyme in complex with a minimal RNA substrate containing an internal 2′-PO4 mark. We also begin to interrogate the basis for an alternative ADP-ribosylation reaction performed by Tpt1 enzymes, namely the transfer of ADP-ribose from NAD+ to a nucleic acid 5′-phosphate end to form a distinctive RNA/DNA-5′-phospho-(ADP-ribose) cap structure (Munir et al. 2018b; Munnur et al. 2019). By solving the structure of an archaeal Tpt1 in complex with 5′-adenosine monophosphate (AMP), we find that Tpt1 has distinct binding modes for a 5′-PO4 mononucleotide versus the RNA 2′-PO4 substrate, whereby their respective 5′ and 2′ phosphates are engaged by the same constellation of active site amino acids but the 2′-PO4 nucleoside and the 5′-nucleoside occupy different sites within the RNA lobe.
RESULTS AND DISCUSSION
Mechanistic insights from a structure of PhoTpt1 in complex with a 2′-PO4 RNA substrate
We solved the structure of a Pyrococcus horikoshii Tpt1 (PhoTpt1) crystal that was soaked in precipitant solution containing a synthetic 2′-PO4-containing RNA trinucleotide ApA2′PpA, which comprises a minimal effective RNA substrate for Tpt1 enzymes (Spinelli et al. 1999; Steiger et al. 2001; Munir et al. 2018a). The refined model at 2.0 Å resolution (Rwork/Rfree = 19.4/24.3) (Table 1) contained ApA2′ppA in the RNA lobe (Fig. 1B). The electron density map is well defined for the pA2′Pp nucleotide and the two flanking ribose sugars (Fig. 1C), albeit less so for the flanking adenine nucleobases. The junction adenosine is flipped into a deep crevice of the RNA lobe (Fig. 2A), where it is sandwiched in a cation-π stack with Arg62 on one side and the aliphatic arm of Lys7 on the other side (Fig. 2B). The junction adenosine is in the anti conformation and receives a hydrogen bond to adenine-N1 from the Ser6 hydroxyl group (Figs. 1C, 2B). Lys7-Nζ donates a hydrogen bond to the junction adenosine ribose-O4′ (Fig. 2B). The N-terminal α1 helix engages the nucleosides flanking the junction nucleotide via: (i) a hydrogen bond from the 3′-flanking adenosine ribose 2′-OH to the Lys7 mainchain carbonyl; (ii) a hydrogen bond from the Tyr11 mainchain amide to the 3′-flanking ribose O3′; (iii) van der Waals interactions of Tyr11 with the 3′-flanking ribose C1′; (iv) a hydrogen bond from Lys7-Nζ to the 5′-flanking ribose O2′; and (v) stacking of Phe3 on the 5′-flanking adenine (Fig. 1C).
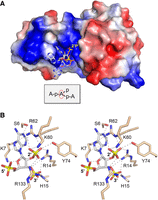
Surface electrostatics and atomic interaction with 2′-PO4 RNA. (A) Surface electrostatic model of the binary substrate complex of PhoTpt1 with ApA2′PpA. The junction adenosine nucleobase is flipped into a deep crevice in the RNA lobe. (B) Stereo view highlighting atomic interactions of the junction nucleotide pA2′Pp (stick model with gray carbons) with Tpt1 amino acids (stick models with beige carbons). Hydrogen bonds are denoted by black dashed lines. Van der Waals contacts are depicted as beige dashed lines.
PhoTpt1 crystallographic data and refinement statistics
The junction 2′-PO4 is extensively coordinated by Arg14, Lys60, Arg62, and Tyr74 in the RNA lobe and Arg133 in the NAD+ lobe (Fig. 2B). The junction nucleotide 3′-PO4 is engaged by Arg14 and His15 (Fig. 2B). To gain insights into the mechanism of step 1 of the Tpt1 pathway, we superimposed the present PhoTpt1•2′PO4 RNA junction structure and the PhoTpt1•NAD+ structure solved previously (Jacewicz et al. 2023; PDB 8TFZ). In the resulting hybrid model of a bisubstrate complex (Fig. 3A), the 2′-PO4 of the junction pA2′pp nucleotide is pointing toward the C1″ of NAD+ at a distance of 4.4 Å, as befits the proposed step 1 reaction chemistry entailing an SN1-like attack of a 2′-PO4 oxygen at C1″ through an oxocarbenium ion transition state. The nicotinamide moiety of NAD+ is packed tightly against the interior surface of the substrate pocket via hydrogen bonding of nicotinamide amide nitrogen and oxygen atoms to the Gly96 carbonyl oxygen and amide nitrogen atoms, respectively (Fig. 3A). Thus, as suggested previously (Jacewicz et al. 2023), we envision that the ADP-ribose sugar moves away from the nicotinamide leaving group during the SN1 displacement, thereby closing the distance between C1″ and the attacking 2′-PO4.
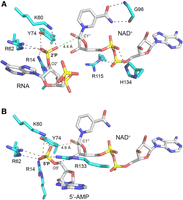
Models of bisubstrate complexes of PhoTpt1 bound to NAD+ and either 2′-PO4 RNA or 5′-AMP. (A) A Tpt1 bisubstrate complex with the junction nucleotide pA2′Pp and NAD+ was modeled by superimposing the Tpt1•ApA2′PpA and Tpt1•NAD+ structures. Hydrogen bonds to the 2′-PO4, the NAD+ phosphates, and the NAD+ nicotinamide are denoted by black dashed lines. The distance from the 2′-PO4 to the C1″ atom of NAD+ is indicated by a green dashed line. (B) A bisubstrate complex with 5′-AMP and NAD+ was modeled by superimposing the Tpt1•5′-AMP and Tpt1•NAD+ structures. Hydrogen bonds to the 5′-PO4 are denoted by black dashed lines. The distance from the 5′-PO4 to the C1″ atom of NAD+ is indicated by a green dashed line.
Reference to the available structures of Tpt1 enzymes bound to products 2′-OH RNA and ADPRp (Banerjee et al. 2019; Jacewicz et al. 2023) suggests that the enzymic contacts to the NAD+ phosphates and the junction 2′-PO4 in the bisubstrate complex model are preserved during the ensuing chemical steps in which the junction phosphate is transferred to NAD+. The phosphate-coordinating amino acids Arg14, His15, Arg62, and Arg115 comprise an “R–H–R–R catalytic tetrad” that is conserved among Tpt1 orthologs and is essential for Tpt1 activity in vivo (Sawaya et al. 2005; Munir et al. 2018a). It is noteworthy that the hydrogen bond of Arg14 with the bridging ribose O2′ of the junction 2′-PO4 in the RNA substrate complex (Fig. 3A) is retained as a hydrogen bond to the junction nucleotide ribose 2′-OH in the RNA product complex (Jacewicz et al. 2023). We propose that Arg14 acts as a general acid catalyst in donating a proton to the bridging ribose O2′ of the RNA-2′-phospho-adenosine diphosphate ribose (ADPR) intermediate during the step 2 transesterification reaction of the Tpt1 pathway.
Because step 1 chemistry occurs at the NAD+ C1″ atom and step 2 chemistry occurs at the 2′-phosphorus atom (Fig. 1A), perturbation of the phosphate contacts can differentially impact the rates of these two steps. This is exemplified by kinetic analyses of the effects of alanine mutations in the catalytic tetrad of Tpt1 from the bacterium Runella slithyformis, which showed that the arginines that coordinate the junction 2′-PO4 in the RNA substrate (corresponding to Arg14 and Arg62 in PhoTpt1) accelerate the rate of step 2 transesterification by 710-fold and 210-fold, respectively (Munir et al. 2018a). Thus, we envision that the first arginine of the tetrad (in addition to its imputed role as general acid; vide supra) and the second arginine collaborate to stabilize a pentacoordinate phosphorane transition state of the 2′-1″ phosphodiester during the step 2 transesterification reaction. By contrast, the two phosphate-coordinating arginines play distinct roles during step 1 catalysis. Whereas mutating the second arginine to alanine slowed step 1 by only threefold, alanine mutation of the first arginine elicited a 380-fold decrement in the step 1 rate constant (Munir et al. 2018a). The present structure of the PhoTpt1•2′-PO4 RNA complex highlights how Arg14 makes four hydrogen bonds to four different phosphate oxygens of the RNA substrate—two to the junction 2′-PO4 and two to the flanking 3′-PO4 (Fig. 2B)—that ensure the proximity and correct orientation of the junction nucleotide for an attack by the 2′-PO4 on the C1″ atom of NAD+.
Structure of PhoTpt1 in complex with 5′-AMP
The catalytic repertoire of Tpt1 enzymes is not limited to the removal of internal RNA 2′-phosphate groups. To wit, several Tpt1 orthologs, including PhoTpt1, can catalyze NAD+-dependent ADP-ribosylation of an RNA or DNA 5′-monophosphate terminus that generates a 5′-phospho-ADP-ribosylated cap structure as an end product (Munir et al. 2018b; Munnur et al. 2019). One-step synthesis of a 5′-phospho-ADP-ribosylated cap structure entails the same chemistry as step 1 of the Tpt1 pathway, except that the polynucleotide 5′-PO4 is the species that attacks the ribose C1″ of NAD+ and displaces nicotinamide. A crucial distinction between the 2′-PO4 and 5′-PO4 ADP-ribosyltransferase reactions is that Tpt1 is apparently unable to catalyze a second chemical 5′-phosphoryltransferase step once the 5′-phospho-ADPR cap structure is formed. Previously, we speculated that the different reaction endpoints with a 5′-PO4 versus 2′-PO4 nucleophile reflect variations in the geometry of the 5′-PO4 and 2′-PO4 linked ADP-ribose moieties in the Tpt1 active site, such that the 2′-phospho-ADP-ribosylated RNA is oriented so that the RNA ribose O2′ is positioned apically relative to the ADP-ribose O2″ in a manner conducive to step 2 cyclization and expulsion of the RNA 2′-OH product, whereas the orientation of the terminal O5′ and the ADP-ribose O2″ in the capped nucleic acid is inimical to cyclization and displacement of the 5′-OH. This model implies that Tpt1 has distinct binding modes for 5′-PO4 and 2′-PO4 substrates.
To gain insights into how Tpt1 might engage a 5′-monophosphate end, we solved the structure of a PhoTpt1 crystal that was soaked in precipitant solution containing 5′-AMP. The refined model at 1.7 Å resolution (Rwork/Rfree = 18.5/20.9) (Table 1) contained two molecules of AMP, one in the NAD+ lobe and one in the RNA lobe (Fig. 4A). The AMP in the NAD+ lobe fits into the surface exposed pocket that accommodates the AMP moiety of the NAD+ substrate (Fig. 4B). The enzymic contacts to this AMP recapitulate those observed in the PhoTpt1•NAD structure (Jacewicz et al. 2023), including: (i) coordination of the AMP ribose 2′-OH and 3′-OH by hydrogen bonding to His95 and Thr97; (ii) engagement of the adenine nucleobase via hydrogen bonds from the Lys111 mainchain amide to adenine-N7 and from adenine-N6 to the Gly109 carbonyl; and (iii) coordination of the AMP phosphate by Arg115 (Fig. 4D).
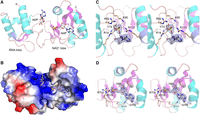
Structure of PhoTpt1 in complex with 5′-AMP. (A) Tertiary structure of PhoTpt1 (cartoon model) with two bound AMP nucleotides (stick models with gray carbons and yellow phosphorus atoms). One AMP is bound to the RNA lobe, and one AMP occupies the position of the NAD+ adenylate in the NAD+ lobe. (B) Surface electrostatic model of PhoTpt1 bound to the two AMP nucleotides (stick models). (C) Stereo view of the AMP in the RNA lobe overlaid with a simulated annealing omit density map (blue mesh contoured at 1 σ), highlighting Tpt1 hydrogen bond contacts to the 5′-PO4 and 3′-OH (black dashed lines). (D) Stereo view of the AMP in the NAD+ lobe overlaid with a simulated annealing omit density map (blue mesh contoured at 1.5 σ), highlighting Tpt1 hydrogen bond contacts to the adenine nucleobase, ribose hydroxyls, and 5′-PO4.
The salient findings from this structure are that whereas the 5′-PO4 of the AMP in the RNA lobe occupies nearly the same position and is engaged by the same amino acids as the junction 2′-PO4 of the canonical Tpt1 RNA substrate, entailing a network of hydrogen bonds to the four AMP phosphate oxygens from Arg14, Lys60, Arg62, Tyr74, and Arg133 (Fig. 4C), the nucleobase moiety of AMP is not flipped into the deep surface crevice that accommodates the junction nucleobase (Fig. 4B). Unlike the junction nucleobase, which makes a π-cation stack on Arg62 (Fig. 2B), the adenine of 5′-AMP makes a π-cation stack on Arg133 (Fig. 4C).
By superimposing the present PhoTpt1•5′-AMP structure and the PhoTpt1•NAD+ structure (Fig. 3B), we see that the nearest 5′-PO4 oxygen is situated 4.6 Å from the C1″ of NAD+. If the position of the 5′-AMP in this structure reflects how the 5′-PO4 end of a nucleic acid binds to Tpt1 during the capping reaction, then we can infer from the bisubstrate model that once the transfer of ADPR to the 5′-PO4 is accomplished, the terminal ribose-O5′ of the polynucleotide will likely not be in a proper orientation relative to the ribose-O2″ of ADPR to allow a second transesterification step to proceed. However, it is possible that the 5′-AMP binding mode, in which the nucleobase resides outside of the cleft that accepts the RNA 2′-PO4 junction nucleobase, is not a true mimetic of a bisubstrate complex of the 5′-PO4 capping reaction.
In conclusion, the present structures further illuminate principles of substrate recognition and catalysis by Tpt1 enzymes. The major challenge at this point is to attain structural snapshots of Tpt1 enzymes as complexes with the RNA-2′-phospho-(ADP-ribose) intermediate of the canonical Tpt1 pathway and a capped RNA-5′-phospho-(ADP-ribose) product of the capping reaction.
MATERIALS AND METHODS
Recombinant PhoTpt1
Tag-free PhoTpt1 was purified from soluble extracts of isopropyl β-d-1-thiogalactopyranoside (IPTG)-induced Escherichia coli BL21(DE3)-CodonPlus–pET28b-His10Smt3•PhoTpt1(2-177) cells as described previously (Jacewicz et al. 2023).
Structure determination
Diffraction data from single crystals were acquired at Argonne National Laboratory (APS) beamline ID-24-C. Reduction of the crystallographic data was performed using program suites (CCP4 1994; Kabsch 2010a,b; Evans 2011; Evans and Murshudov 2013) installed in automated processing RAPD pipeline at APS. Phasing and structure refinement were performed in PHENIX (Liebschner et al. 2019). Iterative model building and adjustment were executed on COOT (Emsley and Cowtan 2004). The diffraction and refinement statistics are compiled in Table 1.
Structure of PhoTpt1 in complex with 2′-PO4 RNA or 5′-AMP
Structures of PhoTpt1 in complex with 2′-PO4 RNA or 5′-AMP were obtained in a series of ligand soaking experiments using preformed crystals of PhoTpt1 that were grown by sitting drop vapor diffusion after mixing aliquots (0.5–2.0 µL) of a 0.2–0.27 mM PhoTpt1 solution with an equal volume of precipitant solution composed of 0.1 m Tris-Bicine (pH 8.3 or 8.5), 0.1 M amino acid additives mixture (0.02 M each of DL-glutamic acid monohydrate, DL-alanine, glycine, DL-lysine monohydrate, and DL-serine), 16%–30% ethylene glycol, and 8%–15% PEG-8000.
Soaks with 2′-PO4 RNA substrate were prepared by transferring PhoTpt1 crystals to a soaking solution containing 0.1 m Tris-Bicine (pH 8.5), 0.1 M amino acid additives mixture, 25% ethylene glycol, 12.5% PEG-8000, and supplemented with 0.195 mM 2′-PO4-containing RNA trinucleotide ApA2′PpA. Crystals were incubated in soaking solution for 60 min and then cryoprotected by transfer to a solution containing 0.1 M Tris-Bicine (pH 8.5), 0.1 M amino acid additives mixture, 30% ethylene glycol, and 10% PEG-8000 before flash-freezing them in liquid nitrogen. A crystal in space group P212121 diffracted to 2.0 Å and contained one PhoTpt1 protomer per asymmetric unit (ASU). The structure was solved by isomorphous replacement in PHENIX.REFINE using the ligand-free 1.97 Å PhoTpt1 polypeptide structure (PDB 8TFI) as a model. Electron density for ApA2′PpA was apparent in the RNA lobe. The final PhoTpt1• AA2′PpA model was refined to Rwork/Rfree of 19.4/24.3 with 172 of 177 amino acids in the favored regions of the Ramachandran plot and no outliers (Table 1).
Soaks with 5′-AMP were prepared by transferring PhoTpt1 crystals to a soaking solution containing 0.1 m Tris-Bicine (pH 8.3), 0.1 M amino acid additives mixture, 32% ethylene glycol, 16% PEG-8000, and supplemented with 10 mM 5′-AMP (Millipore Sigma). Crystals were incubated in soaking solution for 4 h and then cryoprotected by transfer to a solution containing 0.05 M Tris-Bicine (pH 8.3), 0.05 M amino acid additives mixture, 18% PEG-8000, and 36% ethylene glycol before flash-freezing them in liquid nitrogen. A crystal in space group P212121 diffracted to 1.71 Å and contained one PhoTpt1 protomer per ASU. The structure was solved by isomorphous replacement in PHENIX.REFINE using the ligand-free PhoTpt1 structure (PDB 8TFI) as a model. Electron density for 5′-AMP was apparent in the NAD+ lobe and in the RNA lobe, with the latter at a marginally lower occupancy of 0.8. The final PhoTpt1•5′-AMP model was refined to Rwork/Rfree of 18.5/20.9 with 174 of 177 amino acids in the favored regions of the Ramachandran plot and no outliers (Table 1).
DATA DEPOSITION
Structure coordinates of the PhoTpt1•ApA2′PpA and PhoTpt1•5′-AMP complexes are available via the RCSB Protein Data Bank under PDB IDs 8TG5 and 8TKB, released on June 5, 2024 and June 19, 2024, respectively.
ACKNOWLEDGMENTS
This research was supported by the National Institutes of Health (NIH) grant R35-GM126945 to S.S. The MSKCC structural biology core laboratory is supported by National Cancer Institute grant P30-CA008748. M.J.D. is supported by the Natural Sciences and Engineering Research Council of Canada Discovery Grants 2016-05126. X-ray diffraction data were collected at synchrotron facilities supported by grants and contracts from the National Institutes of Health (P30-GM124165, HEI-S10OD021527) and the Department of Energy (DE-AC02-06CH11357). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080444.125.
- Received March 4, 2025.
- Accepted April 9, 2025.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








