
An easy tool to monitor the elemental steps of in vitro translation via gel electrophoresis of fluorescently labeled small peptides
- Valeriya I. Marina1,2,10,
- Medina Bidzhieva3,4,10,
- Andrey G. Tereshchenkov2,5,
- Dmitry Orekhov6,
- Vladislava E. Sagitova2,
- Nataliya V. Sumbatyan2,
- Vadim N. Tashlitsky2,5,
- Artem S. Ferberg2,
- Tinashe P. Maviza1,
- Pavel Kasatsky3,
- Olga Tolicheva3,
- Alena Paleskava3,4,
- Vladimir I. Polshakov7,
- Ilya A. Osterman1,
- Olga A. Dontsova1,2,5,8,
- Andrey L. Konevega3,4 and
- Petr V. Sergiev1,2,5,9
- 1Center for Molecular and Cellular Biology, Skolkovo Institute of Science and Technology, Skolkovo 121205, Russia
- 2Department of Chemistry, Lomonosov Moscow State University, Moscow 119991, Russia
- 3Molecular and Radiation Biophysics Division, Petersburg Nuclear Physics Institute named by B.P. Konstantinov of NRC “Kurchatov Institute,” Gatchina 188300, Russia
- 4Institute of Biomedical Systems and Biotechnologies, Peter the Great St. Petersburg Polytechnic University, Saint Petersburg 195251, Russia
- 5A.N. Belozersky Institute of Physico-Chemical Biology, Lomonosov Moscow State University, Moscow 119991, Russia
- 6R&D Department, VIC Animal Health, Severny, Belgorod Region 308519, Russia
- 7Faculty of Fundamental Medicine, Lomonosov Moscow State University Moscow, Moscow 119991, Russia
- 8Department of Functioning of Living Systems, Shemyakin-Ovchinnikov Institute of Bioorganic Chemistry, Moscow 117997, Russia
- 9Institute of Functional Genomics, Lomonosov Moscow State University, Moscow 119991, Russia
- Corresponding authors: petya{at}genebee.msu.ru, konevega_al{at}pnpi.nrcki.ru
-
↵10 These authors contributed equally to this work.
-
Handling editor: Eric Westhof
Abstract
Several methods are available to visualize and assess the kinetics and efficiency of elemental steps of protein biosynthesis. However, each of these methods has its own limitations. Here, we present a novel, simple and convenient tool for monitoring stepwise in vitro translation initiated by BODIPY-Met-tRNA. Synthesis and release of very short, 1–7 amino acids, BODIPY-labeled peptides, can be monitored using urea–polyacrylamide gel electrophoresis. Very short BODIPY-labeled oligopeptides might be resolved this way, in contrast to widely used Tris-tricine gel electrophoresis, which is suitable to separate peptides larger than 1 kDa. The method described in this manuscript allows one to monitor the steps of translation initiation, peptide transfer, translocation, and termination as well as their inhibition at an unprecedented single amino acid resolution.
Keywords
INTRODUCTION
Translation, or protein biosynthesis, is one of the key steps of gene expression. The central role in translation is played by the ribosome, a giant ribonucleoprotein machine. Insight into the molecular mechanism of ribosome functionality is paramount for fundamental biology. The 70S ribosome is also the primary target for the treatment of bacterial infections. The arsenal of antibiotics, subdivided into defined classes, targets ribosomes at nearly all stages of translation (Wilson 2014).
Translation in bacteria starts with the interaction of the ribosomal small subunit (30S) with mRNA via the Shine–Dalgarno sequence. At the same time, the mRNA start codon interacts with the initiator fMet-tRNAfMet, with the assistance of initiation factors IF1, IF2·GTP, and IF3. Upon recruitment of the large 50S ribosomal subunit and GTP hydrolysis by IF2, the elongation cycle begins. During elongation, the ribosome interacts with amino-acyl(aa)-tRNA·EF-Tu·GTP complex, where the tRNA anticodon should be cognate for the mRNA codon in the A site. Upon correct codon recognition, the GTP hydrolysis triggers EF-Tu·GDP dissociation and accommodation of aa-tRNA into the A/A site. The peptidyl transferase center of the 50S subunit catalyzes transpeptidation between peptidyl-tRNA in the P site and aa-tRNA in the A site. After the peptide bond formation, EF-G·GTP assists in the translocation of two tRNA molecules between the binding sites together with the mRNA movement by one codon. When the ribosome encounters one of three stop codons, release factor (RF) 1 or 2 assists in peptidyl-tRNA hydrolysis, which is followed by RF1(2) removal with RF3·GTP, and ribosome dissociation into the subunits is catalyzed by the ribosome recycling factor (RRF) and EF-G·GTP (Rodnina 2018).
Protein biosynthesis has been studied for several decades and a huge armory of methods has been developed to monitor the efficiency of basic steps of the translation cycle. Translation efficiency might be evaluated by reporter constructs in low-throughput (Grentzmann et al. 1998; Osterman et al. 2013) and high-throughput (Evfratov et al. 2017; Osterman et al. 2020) assays, including genome-wide assessment, aided by ribosome profiling (Ingolia et al. 2009). At a smaller scale, synthesis of a particular protein in vitro might be monitored by the activity of the protein synthesized (Kisly et al. 2021), radioactive (Bartetzko and Nierhaus 1988; Polikanov et al. 2014) or fluorescent label inclusion (Gite et al. 2000; Mittelstaet et al. 2013; Koubek et al. 2015; Buhr et al. 2016). However, synthesis of a full-length protein, or lack of it, is not indicative of a particular stage of translation which might be affected by a mutation, antibiotics, ribotoxins, and/or auxiliary protein factors (Lancaster et al. 2008; Wilson 2014; Buhr et al. 2016). To gain a deeper insight into the efficiency of partial steps of protein synthesis over a range of conditions, a set of more specific methods can be utilized to assess the process. Monitoring the fluorescence intensity changes or Forster resonance energy transfer (FRET) (Rodnina et al. 1997; Milon et al. 2007; Lai and Ermolenko 2018) efficiency in bulk solution or with the single-molecule setup provides a wealth of data, although these procedures are low throughput, demand a high workload and require sophisticated equipment which may not be accessible for the majority of molecular biologists. Codon-wise progression of the ribosome along a particular mRNA in an in vitro translation system(s) can be conveniently monitored via toe-printing, a method that utilizes the reverse transcription along the mRNA to determine the ribosome positions (Hartz et al. 1988; Ermolenko et al. 2013; Orelle et al. 2013; Egorova et al. 2019). Other methods aimed at deciphering the efficiency of peptidyltransferase and peptide release reaction are available, such as SPARK, a scintillation proximity assay for ribosomal kinetics (Polacek et al. 2002), a method combining radioactive (tritium) and biotinylated labels to detect the products of the peptidyl transferase reaction by their immobilization on the scintillant-containing beads. A more direct approach uses radioactive di- or tripeptide resolution via HPLC combined with scintillation counting of the fractions (Vorstenbosch et al. 1996). Such methods are labor intensive and generally not suited for high throughput (Osterman et al. 2016a).
Introduction of an assortment of fluorophores into the components of protein biosynthesis apparatus has been a common tool for studying translation since the early days of ribosome research (Wintermeyer and Zachau 1974). Few naturally fluorescent tRNA nucleotides allow steady-state or pre-steady-state experiments monitoring fluorescence changes and/or FRET assays to study various elemental steps of protein biosynthesis (Paulsen et al. 1982). Additionally, several methods have been developed to introduce fluorescent labels into the components of the translation apparatus. Thorough side-by-side kinetic experiments demonstrated that fluorescently labeled and unlabeled components are functionally equivalent (Robertson et al. 1986; Betteridge et al. 2007; Fei et al. 2010; Stevens et al. 2012).
Molecules of tRNA labeled by proflavine at D16/17, or fluorescein at s4U8 residues proved to be reliable tools (Cooperman 2021). Its application resulted in huge amounts of data which allowed to construct kinetic models of initiation (Milon et al. 2008), decoding (Blanchard et al. 2004; Gromadski et al. 2006; Mittelstaet et al. 2013), and translocation (Rodnina et al. 1997; Savelsbergh et al. 2003).
Introducing fluorophores into amino acids was needed to analyze the stepwise growing of the peptide chain (Gite et al. 2000; Mittelstaet et al. 2013; Buhr et al. 2016; Thommen et al. 2022), as well as to study the process of protein folding by measuring the distance between two fluorophores by FRET (Woolhead et al. 2004; Liutkute et al. 2020).
Originally introduced by Treibs and Kreuzer (1968), tiny fluorophore BODIPY was used to substitute the primary amino group on Met-tRNAfMet (Gite et al. 2000), thus creating labeled amino acid moiety on initiator tRNA. Despite the placement of the fluorophore in the peptidyl transferase center, BPY-Met-tRNAfMet (Fig. 1A) was completely competent in all reactions, including the full cycle of initiation, decoding, A-site binding of the next aminoacyl-tRNA, translocation, and following elongation cycles. Side-by-side comparison of BPY-Met-tRNAfMet with fMet-tRNAfMet did not reveal any significant differences in all possible reactions in vitro (Mittelstaet et al. 2013; Paleskava et al. 2021).
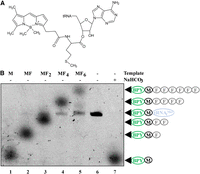
BPY-Met-tRNAfMet structure (A) and products generated using BPY-Met-tRNAfMet and a transcription–translational coupled system (B). The mRNA coding sequences of DNA templates that were used are depicted above each lane, and the products visualized on the gel are labeled (shown in arrows). The original BPY-Met-tRNAfMet corresponds to lane 6, while lane 7 contains the BPY-Met formed by ester bond hydrolysis by 1 M NaHCO3. Other spots (lanes 1–5) correspond to the products of translation: M, MF, MF2, MF4, and MF6. BPY is marked with a green oval—BODIPY label; black circle—methionine; gray circles—phenylalanine residues; blue oval—tRNAfMet.
Usage of BPY-Met-tRNAfMet in a fully controlled reconstituted in vitro translation system, followed by a separation of the resultant peptides by Tris-tricine gel (Schägger and Von Jagow 1987) allowed to decipher the unique mechanism of translational bypassing (Samatova et al. 2014). Bright fluorophore, stoichiometrically (1:1) bound to all synthesized peptides, demonstrated ultra-high sensitivity. Less than 0.1 pmol of peptide per band can be detected. Moreover, BPY-labeled initiator tRNA was successfully used to measure the efficiency of 30S and 70S (Goyal et al. 2015; Vinogradova et al. 2020) initiator complex formation.
While being a nice tool to measure peptide chains longer than 50 amino acids, the Von Jagow system (Schägger and Von Jagow 1987) is not applicable in studies of short (2–20 amino acids ) peptides, thus limiting the application of fluorescently labeled amino acids.
In this study, we applied a procedure of translation product labeling by the fluorescent N-terminal BODIPY group introduced via initiator BPY-Met-tRNAfMet. Here we show that short oligopeptides (1–7 amino acids long), N-terminally labeled by BODIPY can be efficiently resolved and visualized by denaturing polyacrylamide gel electrophoresis, urea PAGE, routinely used for the nucleic acid's separation. This method provides a novel possibility to monitor the growth of peptide chain and its release in the reconstituted in vitro translation system.
RESULTS
Short BODIPY-containing peptides could be visualized together with BPY-Met-tRNA by a denaturing urea PAGE
Our initial goal was to visualize peptidyl-tRNAs containing BPY-Met separated by electrophoresis. Indeed, one can easily detect BPY-Met-tRNAfMet, as a thin band on the RNA urea PAGE (Fig. 1B, lane BPY-Met-tRNAfMet). However, while using the BPY-Met-tRNAfMet as an initiator tRNA in the in vitro translation kit (NEB) programmed with AUG UAA (M); AUG UUC UAA (MF); AUG (UUC)2 UAA (MF2); AUG (UUC)4 UAA (MF4); and AUG (UUC)6 UAA (MF6) templates, we also unexpectedly observed the products of translation on the same gel (Fig. 1B, lanes M, MF, MF2, MF4, MF6). The mobility of peptides decreases with an increase in their length in the expected order. This observation was completely unanticipated, since normally gel electrophoresis is unable to resolve oligopeptides of such an extremely short length. Moreover, normally to resolve peptides, SDS is used to denature and charge protein chains (Schägger and Von Jagow 1987), while SDS was not present in urea PAGE.
BODIPY-labeled peptides bound to the tRNA can be distinguished after hydrolysis by NaHCO3
While BODIPY-labeled small peptides synthesized on the ribosome and released could be easily distinguished by PAGE, they could not be distinguished while they were still bound to the tRNA (Fig. 2A, lanes corresponding to the products of translation by a kit devoid of RFs without NaHCO3 treatment). To distinguish different peptides bound to tRNA, we analyzed four different variants of alkaline agents for hydrolysis of peptidyl-tRNA, namely Tris (pH 9.0), Tris-HCl (pH 8.0), 1 M NaHCO3, and 1 M Na2CO3 (Fig. 2B). Treatment with 1 M Na2CO3, Tris (pH 9.0), and 1 M NaHCO3 appeared to be most effective, the latter being optimal due to the minimal further degradation of the released peptide. We make use of an in vitro translation PURExpress ΔRF123 Kit (NEB) devoid of RFs to obtain ribosomes carrying BPY-M-tRNAfMet, BPY-MF-tRNAPhe, BPY-MF2-tRNAPhe. Treatment of the translation mixtures with 1 M NaHCO3 released M, MF, and MF2 peptides and made it possible to resolve the peptides via PAGE (Fig. 2A, lanes corresponding to the products of translation by a kit devoid of RFs with NaHCO3 treatment).
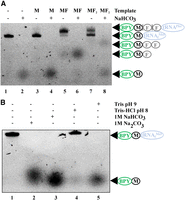
Peptides attached to tRNA could be distinguished by urea PAGE following hydrolysis by 1 M NaHCO3. (A) Intact BPY-M-tRNAfMet (1) and products of M, MF, and MF2—coding mRNA translation, BPY-M-tRNAfMet (3), BPY-MF-tRNAPhe (5), BPY-MF2-tRNAPhe (7), and the products of their hydrolysis by 1 M NaHCO3 (2, 4, 6, 8) separated by urea PAGE. (B) Testing the conditions for BPY-M-tRNAfMet hydrolysis. Lanes correspond to the lack of treatment (1) and treatment with 1 M Na2CO3 (2), 1 M NaHCO3 (3), Tris-HCl (pH 8.0) (4), and Tris (pH 9.0) (5) as indicated. BPY is marked with a green oval—BODIPY label; black circle—methionine; blue oval—tRNAfMet or tRNAPhe.
Separation and visualization of short BODIPY-containing peptides is compatible with in vitro translation system from the purified components and commercial BPY-labeling reagent
To verify that the bands observed correspond to the expected products of translation, we chemically synthesized BPY-MF2 and BPY-MF4 peptides and found their mobilities to be identical to those of the BPY-MF2 and BPY-MF4 peptides translated in vitro (Fig. 3A).
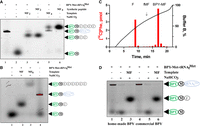
Separation of the BPY-labeled short peptides produced by a variety of systems. (A) Comparison of electrophoretic mobilities of MF2 (3, 4) and MF4 (5, 6) peptides obtained in a course of coupled transcription–translation process (3, 5) and those of chemically synthesized peptides (4, 6). The components added to the translation reaction or loaded to the gel (in the case of chemically synthesized peptides) are designated above the lanes. BPY is marked with a green oval—BODIPY label; black circle—methionine; gray circles—phenylalanine residues; blue oval—tRNAfMet. Lanes 1 and 2 correspond to the BPY-M-tRNAfMet and BPY-M, respectively. (B) Electrophoretic separation of MF and MF6 peptides produced in a translation system assembled from the purified components with premade mRNA. Designations are the same as for panel A. (C) Separation of dipeptides (BPY-M[14C]F, fM[14C]F), synthesized by in vitro translation reconstituted from the purified components, by HPLC in convex gradient of acetonitrile as described in Materials and Methods. BPY-labeled MF dipeptide is eluted four fractions later than nonlabeled MF dipeptide (depicted by arrow). (D) Comparison of the suitability of the BPY-M-tRNAfMet samples produced with a “homemade” sulfosuccinimide ether of BPY (lanes 1–3) or commercial sulfosuccinimide ether of BPY (lanes 4–6) for in vitro translation, reconstituted from the purified components, of MF-coding mRNA. Designations are the same as for panel A.
To check that the observed products are not artifacts of the coupled in vitro transcription–translation system, we set up a translation system from individual components (Paleskava et al. 2021) supplied by presynthesized MF and MF6-coding mRNA (Fig. 3B). It appears that the detection method presented here is compatible with translation mixture from purified components. The bands observed could not be attributed to hypothetical by-products of RNA polymerase or other enzymes present in the coupled transcription–translation mixture.
Since we presented here the new method for short peptide detection, we set up to compare it with the standard method used previously. To this end, we performed in vitro translation of the MF-coding mRNA in the protein synthesis system assembled from the purified components using BPY-Met-tRNAfMet and EF-Tu⋅GTP⋅[14C]Phe-tRNAPhe. Dipeptide formation was monitored by the standard HPLC-based separation of small peptides followed by scintillation counting (Fig. 3C). We detected an efficient synthesis of BPY-Met—[14C]Phe, whose mobility in HPLC was expectedly decreased relative to that of fMet—[14C]Phe. Needless to say, HPLC-based detection of the radioactive peptide is more labor demanding and requires an isotope laboratory, in contrast to the method based on urea PAGE and fluorescence scanning.
The synthesis of sulfosuccinimide ether of BPY fluorophore is complex and requires equipment unavailable for the majority of molecular biology laboratories. We checked a commercially available BPY sulfosuccinimide ether (Thermo Fisher Scientific, cat. D6140) to modify Met-tRNAfMet. BPY-M-tRNAfMet labeled by a commercially available reagent was compared side-by-side with BPY-M-tRNAfMet prepared with the home-made sulfosuccinimide ether of BPY in a translation of MF-coding mRNA (Fig. 3D). Both home-made and commercially available fluorophores were found to be identically suitable for short peptide synthesis and detection.
Leucine-containing peptides might be visualized similarly
Since its development, the poly(U)-dependent system for synthesis poly(Phe) (Bartetzko and Nierhaus 1988) was the most frequently used model to study the translation machinery in vitro. Yet, we demonstrated that urea PAGE is suitable for the separation and detection of oligo(Phe) peptides, starting with BPY-Met. However, the polymerization of other amino acids must be evaluated to draw more general conclusions upon in vitro protein biosynthesis studies. To this end, we set up to check if other BODIPY-labeled short peptides might be visualized similarly. Translation of MLL-coding mRNA followed by a separation of the products by urea PAGE (Fig. 4) resulted in a visualization of ML2 peptide formed with approximately the same yield and migrating with approximately the same mobility to that of MF2 peptide. Thus, short peptides containing uncharged amino acids, such as leucine, can be visualized similarly.
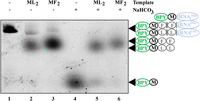
Comparison of electrophoretic mobilities of BPY-ML2 (2, 5) with that of BPY-MF2 (3, 6). Products of the coupled transcription–translational system PURExpress Kit using mRNAs coding for MFF and MLL peptides. Lanes 1 and 4 correspond to the BPY-M-tRNAfMet and BPY-M, respectively. The products of peptidyl-tRNA hydrolysis by 1 M NaHCO3 are separated in lanes 4–6. BPY marked with a green oval—BODIPY label; black circle—methionine; gray circles—phenylalanine residues or leucine residues; blue oval—tRNAfMet.
Activity of translation termination factors can be monitored by BODIPY-peptides gel electrophoresis
To determine the efficiency of translation termination, we performed the in vitro translation of MF-coding mRNAs with different stop codons (MF-UAA, MF-UAG, MF-UGA) in the presence of RFs combinations (RF1 + RF3; RF2 + RF3) (Fig. 5A). Specific stop codon recognition by either RF1 or RF2 proteins could be observed. The RF2 + RF3 combination is less effective than RF1 + RF3, in agreement with earlier observations (O'Connor and Gregory 2011). RF-dependent peptidyl-tRNA hydrolysis might be compared with that facilitated by NaHCO3 treatment (Fig. 5B). Thus, the system is suitable to monitor the polypeptide synthesis, as well as peptide release.
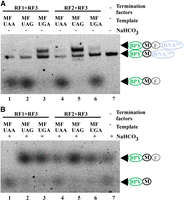
Monitoring translation termination efficiency by urea PAGE. MF-coding mRNAs with different stop codons (lanes 1, 4—UAA; 2, 5—UAG; 3, 6—UGA) were translated in vitro in the presence of BPY-M-tRNAfMet by PURExpress ΔRF123 Kit with the addition of either RF1 and RF3 (1–3) or RF2 and RF3 (4–6) RFs as indicated above the lanes. Lane 7 corresponds to the BPY-M-tRNAfMet. (A) Separation of translation products without chemical post-treatment. (B) Separation of translation products after hydrolysis by 1 M NaHCO3. BPY is marked with a green oval—BODIPY label; black circle—methionine; gray circle—phenylalanine residue; blue oval—tRNAfMet.
BODIPY-labeled peptide separation might be used to study peptidyl transferase inhibitors
Another useful application of this method is the possibility to reveal the mode of action for the specific inhibitors which are able to block the peptidyl transferase, translocation, or termination steps. We used the panel of antibiotics, as presented in Figure 6. Among the antibiotics, we have tested the inhibitors of translocation (such as thiostrepton blocking EF-G catalyzed translocation [Walter et al. 2012]), antibiotics that bind within the ribosome exit tunnel and block translation after synthesis of the several (specific) amino acids (such as erythromycin [Hansen et al. 2002; Sothiselvam et al. 2014], tylosin, and etamycin [Osterman et al. 2016b]), and antibiotics that block formation of the first peptide bond (such as madumycin [Osterman et al. 2017] and retapamulin [Yan et al. 2006]). We also tested the activity of blasticidin S, the inhibitor of peptide bond formation and termination (Svidritskiy and Korostelev 2018), and puromycin, an antibiotic that binds to the ribosomal A site and intercepts a growing peptide chain (Semenkov et al. 1992).
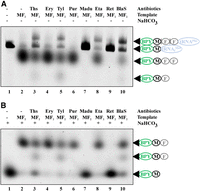
Antibiotics' mechanism of action might be addressed by BODIPY-peptides separation by urea PAGE. Shown are the products of MF2-coding mRNA in vitro translation in the presence of BPY-M-tRNAfMet and antibiotics indicated above the lanes. (A) No post-treatment of the translation mixtures. (B) Translation mixtures after synthesis were treated with 1 M NaHCO3. Lane 1 corresponds to the BPY-M-tRNAfMet. DMSO 1% (2) corresponds to a positive no antibiotic control, (Ths) thiostrepton (3), (Ery) erythromycin (4), (Tyl) tylosin (5), (Pur) puromycin (6), (Madu) madumycin (7), (Eta) etamycin (8), (Ret) retapamulin (9), (BlaS) blasticidin S (10). BPY is marked with a green oval—BODIPY label; black circle—methionine; gray circle—phenylalanine residue; blue oval—tRNAfMet or tRNAPhe.
As expected, we observed the complete inhibition of transpeptidation upon madumycin and retapamulin treatment. Partial inhibition of BPY-MF-tRNAPhe and BPY-MF2-tRNAPhe synthesis was observed for thiostrepton, tylosin, and blasticidin S treatments. Erythromycin and puromycin had no observable effects, likely due to the inability of erythromycin to inhibit short MF/MF2 synthesis, while the puromycin binding was too weak and slow to compete in a full translation system. Interestingly, etamycin blocked translation at a stage after the formation of BPY-MF2-tRNAPhe but prior to peptidyl-tRNA hydrolysis. Thus, the system we present here can be efficiently used to classify the mechanisms of action of various antibiotics.
DISCUSSION
Translation apparatus has been studied for decades and an enormous arsenal of methods has been applied to this field. However, many well-established methods are labor-intensive, give only partial information, or require specific equipment or facilities, such as isotope-laboratories, which are not available in most of the laboratories. Advancement of the ribosome along mRNA could be conveniently monitored by toe-printing (Hartz et al. 1988). This method, however, generally requires radioactive isotopes, and even if not (Egorova et al. 2019), still, it provides little information on the formation of the peptide bond. Assessment of the latter generally requires not only the usage of radioactive isotopes, but also HPLC-based separation of radioactive oligopeptides (Vorstenbosch et al. 1996).
It is hardly an overstatement that practically all molecular biology laboratories could run denaturing polyacrylamide gel electrophoresis, used routinely to separate nucleic acids. Here we show that this simple method may be combined with commercially available in vitro translation kits and commercially available BPY succinimide ether, which could be used to make BPY-Met-tRNAfMet. The method is suitable to monitor the efficiency of very short peptides, one to seven amino acids long, synthesis, and release. Previously published gel electrophoresis methods of peptide separation (Schägger and Von Jagow 1987) and visualization (Gite et al. 2000) are usable for separation of translation products >1 kDa, with optimal resolution for small proteins longer than 5 kDa. Such resolution is not suitable to monitor the efficiency of a particular step of the ribosomal working cycle, such as peptide transfer or peptide release. Serendipitous discovery of anomalous mobility of the very short BPY-labeled peptides, down to a single amino acid, in urea PAGE opens a possibility to conveniently study partial reactions of protein biosynthesis.
We also demonstrated that the method is suitable for at least Phe- and Leu-containing peptides as well as for assessment of antibiotics' mechanism of action. We are sure that this simple and elegant method has great potential to be widely applied for studying protein biosynthesis.
MATERIALS AND METHODS
Preparation of DNA and mRNA templates
DNA templates containing short open reading frames coding for M, MF, MF2, ML2, MF4, and MF6 were obtained by PCR using the plasmid pRFPCER (Osterman et al. 2013) as a template for PCR amplification. Each template harbored the T7 promoter and SD sequence, followed by a different codon set. For instance, the M template contained the start codon (AUG), followed immediately by the stop codon (UAA); the MF template encoded two amino acids, methionine and phenylalanine (AUG UUC) followed by the UAA stop codon, and so on. The complete list of templates can be found in Supplemental Table 1.
Preparation of fluorescently labeled initiator tRNA
Purified from an Escherichia coli strain, individual initiator tRNAfMet was aminoacylated and modified with BODIPY sulfosuccinimide ester (Supplemental Figs. S1 and S2). The resulting BPY-Met-tRNAfMet was purified (Supplemental Fig. S3) as described in the Supplemental Material. The detection limit for BPY-labeled Met-tRNAfMet with modern fluorescent scanners was found to be as low as 1 fmol (Supplemental Fig. S4).
In vitro translation
Coupled transcription–translation was set up in 5 µL volumes using either a PURExpress ΔRF123 Kit or a PURExpress Δ(aa, tRNA) Kit (NEB), with the addition of 20 ng of DNA template (M, MF, MF2, MF4, MF6, or ML2) containing a T7 promoter upstream of the coding sequence. In addition, 0.2 µM BPY-Met-tRNAfMet was added to each reaction. The reactions were carried out for 30 min at 37°C and then divided into two parts. To the first part of the split sample, 2 µL of 1 M NaHCO3 was added, subsequently followed by incubation at 37°C for 20 min. The reactions were stopped by adding standard formamide dye without xylene cyanol (95% deionized formamide, 0.025% [w/v] bromophenol blue, 0.5 mM EDTA). Samples were preheated for 3 min at 70°C and then loaded to a 10% denaturing PAGE (RNA gel) (19:1 AA:bisAA; 1× TBE buffer; 7M urea).
For antibiotic activity assay, ribosomes were used as a limiting component. BPY-Met-tRNAfMet at 0.1 µM concentration was diluted with 0.33 µM fMet-tRNAfMet so that a ribosome should cycle several times to convert all BPY-Met-tRNAfMet into the products of translation. All antibiotics were dissolved in DMSO; the concentration of all antibiotic stock solutions was 50 µM, except for erythromycin and etamycin, which were used at 70 µM concentration. Chemically synthesized peptides MF2 or MF4 were loaded onto the gel at 0.1 µM concentration each as positive controls where indicated.
In vitro translation system on the basis of individually purified components was assembled in the buffer containing 50 mM Tris-HCl (pH 7.5), 70 mM NH4Cl, 30 mM KCl, and 7 mM MgCl2, as described previously (Paleskava et al. 2021). MF and MF6 mRNAs were obtained by T7 transcription followed by purification on a HiTrap Q HP anion exchange column (Cytiva). mRNA sequences are listed in Supplemental Table 2. Initiation complexes were assembled with BPY-Met-tRNAfMet in the P site and either of two mRNAs. Ten picomoles of initiation complexes were mixed with 15 pmol of ternary complexes EF-Tu⋅GTP⋅[14C]Phe-tRNAPhe (1067 dpm/pmol) in the case of MF and 60 pmol of ternary complexes and 2 μM EF-G in the case of MF6, followed by incubation for 2 min at 37°C. The amount of [14C]Phe bound to the ribosomal complexes was determined by filtration through nitrocellulose filters (0.45 μm, Sartorius). The fraction of 14C-Phe bound to the ribosomes on the filters was measured using the Tri-Carb liquid scintillation counter upon addition of toluene scintillation mixture containing 0.4% PPO and 0.04% POPOP. The efficiency of BPY-Met-Phe formation was determined by reversed-phase HPLC on RP8 (Merck) with an acetonitrile gradient in 0.1% trifluoroacetic acid. Beforehand, samples were quenched with 1/10 volume of 5 M KOH, hydrolyzed for 30 min at 37°C, and neutralized with 1/10 volume of glacial acetic acid. The percentage of synthesized peptide was determined by incorporation of the radioactive label as a ratio of peptide formed to the amount of the 70S ribosomes in the reaction mixture. Evaluation of BPY-Met-Phe6 formation by peptide analysis was not possible due to its precipitation upon sample treatment.
Detection of peptides in a gel
Gels were scanned by a Typhoon FLA 9500 Biomolecular Imager (GE Healthcare) in the Cy2 channel with excitation Peak (492 nm) and emission Peak (510 nm). Results were visualized and intensities calculated using the Image J program. The data were normalized to the background.
Synthesis of chemical compounds
Reagents
1,1,1,3,3,3-hexafluoroisopropanol (HFIP) from Acros Organics, Fmoc-amino acids and 2-chlorotrityl chloride resin for solid phase peptide synthesis (2CTC Resin) from Iris Biotech, O-(benzotriazol-1-yl)-N,N,N′,N′-tetramethyluronium hexafluorophosphate (HBTU), diisopropylethylamine (DIPEA) from Sigma-Aldrich, succinimide ester of BODIPY-fluorescent dye (BODIPY-C3-SE) from Lumiprobe (Russian Federation), anhydrous solvents such as methanol from LiChrosolv and isopropanol, dioxane, pyridine, N,N-dimethylformamide (DMF), CH2Cl2, acetonitrile from PanReac AppliChem, and BODIPY FL sulfonated succinimidyl ester from Thermo Fisher Scientific were used in this work.
The sodium salt of BODIPY sulfosuccinimidyl ester (Gießler et al. 2010), BODIPY-Met-(Phe)2 (BODIPY-MF2), BODIPY-Met-(Phe)4 (BODIPY-MF4), and BPY-tRNAfMet were synthesized as described in the Supplemental Material.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
This work was supported by the Russian Science Foundation (grant number 21-64-00006 to O.A.D. for assay development), the Russian Science Foundation (grant number 22-14-00278 to A.L.K., for RNA labeling, reconstituted translation system). The authors thank the MSU development program for access to SeqStudio for sequencing of genetic constructs.
Author contributions: V.I.M.: investigation, methodology, writing—original draft; M.B.: investigation, methodology; A.G.T.: resources; D.O.: investigation, resources; V.E.S.: investigation; N.V.S.: resources; V.N.T.: investigation; A.S.F.: investigation; P.K.: investigation; O.T.: investigation; T.P.M.: investigation; A.P.: investigation; V.I.P.: investigation; I.A.O.: methodology; O.A.D.: supervision; A.L.K.: supervision, writing—review and editing, funding acquisition; P.V.S.: supervision, conceptualization, writing—review and editing, funding acquisition.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079766.123.
- Received July 5, 2023.
- Accepted November 22, 2023.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHORS


Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Valeriya I. Marina and Medina Bidzhieva are co-first authors of this paper, “An easy tool to monitor the elemental steps of in vitro translation via gel electrophoresis of fluorescently labeled small peptides.” Valeriya is a PhD student at Lomonosov Moscow State University focusing on the development of methods for studying the effect of antibiotics on the translation mechanism of the bacterial ribosome. Medina is a master's student at Peter the Great St. Petersburg Polytechnic University.
What are the major results described in your paper and how do they impact this branch of the field?
Thanks to the fact that we discovered unusual activity of the BODIPY tag in PAGE UREA, we were able to develop a method that has the amazing ability to resolve very short peptides, down to a single amino acid. Moreover, this method allows one to monitor the steps of translation initiation, peptide transfer, translocation, and termination as well as their inhibition at an unprecedented resolution of a single amino acid.
What led you to study RNA or this aspect of RNA science?
V.I.M.: RNA is one of the most important classes of compounds in the cell, thanks to which it becomes possible to study crucial processes in the cell, in particular new, unknown aspects of protein biosynthesis.
M.B.: One of the key steps in gene expression is protein biosynthesis, and I was lucky to work on such an important topic with a friendly team and in a great laboratory.
During the course of these experiments, were there any surprising results or particular difficulties that altered your thinking and subsequent focus?
V.I.M.: In many ways, we were greatly influenced by the discovery of the unusual mobility of BODIPY-labeled peptides in RNA gels. This is what motivated us to develop the method so that it can be used endlessly for various purposes.
M.B.: In addition to what my colleague said, I was surprised by the sensitivity of the described method.
What are some of the landmark moments that provoked your interest in science or your development as a scientist?
V.I.M.: I believe this is an opportunity to get an education at the best university in the country and work side by side with leading scientific minds, to feel their support and not be afraid to make mistakes, to search, fail, not give up and keep moving forward to new discoveries.
M.B.: I studied at a physical-technical high school opened by a Nobel laureate in physics, Zhores I. Alferov. There I fell in love with physics and chemistry, so I decided to devote my life to science.
If you were able to give one piece of advice to your younger self, what would that be?
V.I.M.: I would advise myself not to be afraid to ask questions, for it is impossible to know everything in the world.
M.B.: I would advise myself to feel free to consult and discuss experiments and mistakes with senior colleagues.
Are there specific individuals or groups who have influenced your philosophy or approach to science?
V.I.M.: Absolutely! I was inspired by my two supervisors, Professor Petr V. Sergiev and Professor Ilya A. Osterman.
M.B.: I was most influenced by my supervisor Andrey L. Konevega and colleagues from our research group.
What are your subsequent near- or long-term career plans?
V.I.M.: I want to get my PhD in the next couple of years and then build my own research group to continue studying bacterial translation.
M.B.: I'm still just at the beginning of my journey, so there are definitely a lot of plans in the long term. But in the near future, I would like to get a master's degree and study for a PhD.
What were the strongest aspects of your collaboration as co-first authors?
V.I.M.: Our groups have been working for a long time, but it is the first time that we are working together (before this, cooperation took place between our supervisors, Petr V. Sergiev and Andrey L. Konevega). I am very grateful that we had the opportunity to work together.
M.B.: When I was offered to work with colleagues from Moscow, I was very excited. It was an amazing experience that allowed us to achieve excellent results.








