
Sod1-deficient cells are impaired in formation of the modified nucleosides mcm5s2U and yW in tRNA
- 1Department of Molecular Biology, Umeå University, 901 87 Umeå, Sweden
- 2Department of Experimental Medical Science, Lund University, 221 00 Lund, Sweden
- Corresponding authors: marcus.johansson{at}med.lu.se, anders.bystrom{at}umu.se, fu.xu{at}dkfz-heidelberg.de
-
Handling editor: Eric Phizicky
Abstract
Uridine residues present at the wobble position of eukaryotic cytosolic tRNAs often carry a 5-carbamoylmethyl (ncm5), 5-methoxycarbonylmethyl (mcm5), or 5-methoxycarbonylhydroxymethyl (mchm5) side-chain. The presence of these side-chains allows proper pairing with cognate codons, and they are particularly important in tRNA species where the U34 residue is also modified with a 2-thio (s2) group. The first step in the synthesis of the ncm5, mcm5, and mchm5 side-chains is dependent on the six-subunit Elongator complex, whereas the thiolation of the 2-position is catalyzed by the Ncs6/Ncs2 complex. In both yeast and metazoans, allelic variants of Elongator subunit genes show genetic interactions with mutant alleles of SOD1, which encodes the cytosolic Cu, Zn-superoxide dismutase. However, the cause of these genetic interactions remains unclear. Here, we show that yeast sod1 null mutants are impaired in the formation of 2-thio-modified U34 residues. In addition, the lack of Sod1 induces a defect in the biosynthesis of wybutosine, which is a modified nucleoside found at position 37 of tRNAPhe. Our results suggest that these tRNA modification defects are caused by superoxide-induced inhibition of the iron-sulfur cluster-containing Ncs6/Ncs2 and Tyw1 enzymes. Since mutations in Elongator subunit genes generate strong negative genetic interactions with mutant ncs6 and ncs2 alleles, our findings at least partially explain why the activity of Elongator can modulate the phenotypic consequences of SOD1/sod1 alleles. Collectively, our results imply that tRNA hypomodification may contribute to impaired proteostasis in Sod1-deficient cells.
Keywords
INTRODUCTION
A general feature of tRNA molecules is that they contain posttranscriptionally modified nucleosides. The presence of these modified nucleosides influences the structure and function of tRNAs, and they, thereby, provide an additional layer of regulation to gene expression (Phizicky and Hopper 2023). The nucleosides in the anticodon region of tRNAs, especially positions 34 (the wobble nucleoside) and 37, are often modified, and modifications at these positions influence anticodon–codon interactions and consequently the efficiency and/or fidelity of translation (Björk and Hagervall 2014; Agris et al. 2017).
Uridines present at position 34 of eukaryotic cytosolic tRNAs are usually modified to a nucleoside of the xm5U type, where the xm5 side-chain is a 5-carbamoylmethyl (ncm5), 5-methoxycarbonylmethyl (mcm5), or 5-methoxycarbonylhydroxymethyl (mchm5) group (Boccaletto et al. 2022). The xm5U nucleosides sometime also harbor an additional 2-thio (xm5s2U) or 2′-O-methyl (xm5Um) group. The first step in the synthesis of the xm5 side-chains is dependent on the Elongator complex, which is composed of six Elp proteins (Elp1-Elp6) (Winkler et al. 2001; Huang et al. 2005; Karlsborn et al. 2014; Kolaj-Robin and Seraphin 2017). Elongator likely catalyzes the formation of cm5U, which is then converted into ncm5U, mcm5U, or mchm5U by other enzymes (Kalhor and Clarke 2003; Huang et al. 2005; van den Born et al. 2011; Karlsborn et al. 2014; Kolaj-Robin and Seraphin 2017). In tRNAs harboring an xm5s2U34 residue, the sulfur is transferred to the 2-position by a conserved thiouridylase complex—designated Ncs6/Ncs2 (or Tuc1/Tuc2) in Saccharomyces cerevisiae and CTU1/CTU2 in humans (Björk et al. 2007; Huang et al. 2008; Nakai et al. 2008; Leidel et al. 2009; Noma et al. 2009). The catalytically active Ncs6 (CTU1) subunit of the 2-thiouridylase complex is an iron-sulfur [3Fe-4S] cluster-containing protein (Liu et al. 2016). The presence of an xm5U residue or a derivative thereof at the wobble position promotes decoding of cognate A- and G-ending codons (Lim 1994; Murphy et al. 2004; Johansson et al. 2008; Kurata et al. 2008), and the inability to form the xm5 and/or s2 groups induces a diverse array of phenotypes (Karlsborn et al. 2014; Kolaj-Robin and Seraphin 2017). Moreover, in both yeast and human cells, the lack of the wobble xm5 and/or s2 groups is associated with increased levels of aggregated endogenous proteins (Nedialkova and Leidel 2015; Bento-Abreu et al. 2018; Rapino et al. 2018).
Wybutosine (yW) and its derivatives are nucleosides found at position 37 of eukaryotic and archaeal phenylalanine tRNA 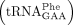 (Boccaletto et al. 2022). The yW nucleoside is derived from a guanosine, and in yeast, the formation of yW requires the activity of five distinct
enzymes: Trm5, Tyw1, Tyw2, Tyw3, and Tyw5, which all have human homologs (Björk et al. 2001; Waas et al. 2005; Noma et al. 2006). The first step in the biosynthesis of yW is the formation of N1-methylguanosine (m1G) by the methyltransferase Trm5 (Droogmans and Grosjean 1987; Björk et al. 2001). Tyw1, which is a [4Fe-4S]-cluster-containing radical-SAM enzyme, then converts m1G into 4-demethylwyosine (imG-14) (Waas et al. 2005; Noma et al. 2006). The imG-14 residue is subsequently converted into yW by additional steps catalyzed by Tyw2, Tyw3, and Tyw4 (Noma et al. 2006). The presence of yW37 in
(Boccaletto et al. 2022). The yW nucleoside is derived from a guanosine, and in yeast, the formation of yW requires the activity of five distinct
enzymes: Trm5, Tyw1, Tyw2, Tyw3, and Tyw5, which all have human homologs (Björk et al. 2001; Waas et al. 2005; Noma et al. 2006). The first step in the biosynthesis of yW is the formation of N1-methylguanosine (m1G) by the methyltransferase Trm5 (Droogmans and Grosjean 1987; Björk et al. 2001). Tyw1, which is a [4Fe-4S]-cluster-containing radical-SAM enzyme, then converts m1G into 4-demethylwyosine (imG-14) (Waas et al. 2005; Noma et al. 2006). The imG-14 residue is subsequently converted into yW by additional steps catalyzed by Tyw2, Tyw3, and Tyw4 (Noma et al. 2006). The presence of yW37 in 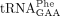 influences its interaction with the cognate codons and the modification is important for maintaining the reading frame during
translation (Carlson et al. 2001; Konevega et al. 2004; Waas et al. 2007).
influences its interaction with the cognate codons and the modification is important for maintaining the reading frame during
translation (Carlson et al. 2001; Konevega et al. 2004; Waas et al. 2007).
Allelic variants of genes encoding Elongator subunits have been linked to several neurological disorders, including amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD) (Simpson et al. 2009; van Blitterswijk et al. 2014; Bento-Abreu et al. 2018; Suzuki 2021). A hallmark of both ALS and FTD is the accumulation of protein aggregates in the affected neurons. The first gene found to be associated with ALS was SOD1, which encodes the cytosolic Cu, Zn-superoxide dismutase, and mutations in SOD1 are now known to account for 15%–30% of familial and around 1% of sporadic cases (Rosen et al. 1993; Zou et al. 2017). The function of superoxide dismutases is to protect cells from reactive oxygen species by catalyzing the conversion of superoxide (O2-) to hydrogen peroxide and molecular oxygen (McCord and Fridovich 1969; Wang et al. 2018). Most ALS-inducing SOD1 alleles are considered to harbor gain-of-function mutations and the mutations usually induce misfolding and aggregation of the SOD1 protein (Saccon et al. 2013; Taylor et al. 2016). Although the toxic properties of the mutant SOD1 proteins are thought to be causative for ALS, the reduction in SOD1 activity may also influence the disease (Saccon et al. 2013). The SOD1 protein is highly conserved, and the phenotypes of an S. cerevisiae sod1Δ strain can be complemented by expression of the human SOD1 protein (Rabizadeh et al. 1995; Corson et al. 1998). In addition to increased sensitivity to oxidative stress, S. cerevisiae sod1Δ mutants are, in the presence of oxygen, auxotrophic for methionine and lysine, and show a leaky leucine auxotrophy (Bilinski et al. 1985; Wallace et al. 2004). Although the precise mechanism underlying these phenotypes remains uncertain, the lysine and leucine auxotrophies were suggested to be a consequence of superoxide-induced damage to the [4Fe-4S] clusters in the biosynthetic Lys4 and Leu1 enzymes (Wallace et al. 2004).
Genetic variants of ELP3 are associated with increased susceptibility for ALS, and the levels of ELP3 protein are reduced in brain tissue from humans carrying risk-associated ELP3 alleles (Simpson et al. 2009). Further, the motor cortex of sporadic ALS patients was found to have reduced ELP3 levels, and the reduction correlates with decreased levels of mcm5s2U-modified tRNAs (Bento-Abreu et al. 2018). The depletion of ELP3 in a motor neuron cell line led to reduced levels of mcm5s2U in tRNA, and with respect to SOD1, the reduction was associated with increased aggregation of mutant but not wild-type variants of the human SOD1 protein (Bento-Abreu et al. 2018). In zebrafish, the expression of human ELP3 reduced the axonopathy associated with expression of the mutant SOD1A4V protein. Similarly, the overexpression of human ELP3 in SOD1G93A mice counteracted the denervation and extended their survival (Bento-Abreu et al. 2018).
In this study, we used S. cerevisiae as a model to investigate the interconnection between Elongator and Sod1. We show that the inactivation of SOD1 leads to reduced levels of the modified nucleosides mcm5s2U and yW in tRNA and that these tRNA modification defects are likely caused by superoxide-induced inhibition of the Ncs6/Ncs2 and Tyw1 enzymes.
RESULTS
Yeast strains deleted for SOD1 or CCS1 show alterations in tRNA modification
The findings that the overexpression of human ELP3 protein is protective in the zebrafish SOD1A4V and mouse SOD1G93A models indicate a genetic link between Elongator and SOD1 (Bento-Abreu et al. 2018). Further, high-throughput studies in S. cerevisiae have revealed negative genetic interactions between a sod1Δ mutation and null alleles of any of several ELP genes (Collins et al. 2007; Lin et al. 2008). To confirm the negative genetic interaction, we compared the growth of isogenic wild-type, sod1Δ, elp3Δ, and sod1Δ elp3Δ yeast strains. These analyses showed that the double mutant indeed grows slower than the respective single mutant at both
30°C and 37°C (Fig. 1A). Since mutations in the yeast ELP3 gene show negative genetic interactions with mutant alleles of genes encoding other tRNA modifying factors, including NCS2 and NCS6 (Björk et al. 2007; Klassen et al. 2015, 2016; Xu et al. 2019), we hypothesized that the lack of Sod1 may be associated with a tRNA modification defect. To investigate this possibility,
we isolated total tRNA from wild-type and sod1Δ strains and analyzed the nucleoside composition by HPLC. We also analyzed total tRNA from a strain deleted for CCS1 (CCS in humans), which encodes the copper chaperone for Sod1 (Culotta et al. 1997). Note that a limitation of using HPLC analyses of total tRNA digests is the difficulty in detecting changes in modified
nucleosides that are present at more than one position in tRNA and catalyzed by different enzymes. The analyses revealed that
the relative abundance of mcm5s2U is mildly reduced in total tRNA from the sod1Δ and ccs1Δ strains (Fig. 1B,D; Supplemental Table S1). Both strains also showed an increased abundance of mcm5U, whereas the levels of ncm5U were unaffected (Fig. 1B,D). In addition to influencing the levels of mcm5-modified uridines, the sod1Δ and ccs1Δ alleles induced reduced levels of wybutosine (yW) (Fig. 1C,E). In the analyses of total tRNA, yW is detected both as a yW nucleoside and a yWpA dinucleotide; the dinucleotide is present
in total tRNA digests due to inefficient hydrolysis of the phosphodiester bond between yW37 and A38 in 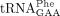 (Björk et al. 2001). We conclude that the lack of Sod1 or Ccs1 induces changes to the tRNA modification profiles.
(Björk et al. 2001). We conclude that the lack of Sod1 or Ccs1 induces changes to the tRNA modification profiles.
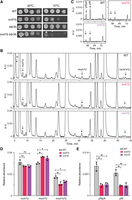
Yeast strains deleted for SOD1 or CCS1 show defects in tRNA modification. (A) Growth of wild-type (MJY1150), sod1Δ (MJY1152), elp3Δ (UMY4563), and sod1Δ elp3Δ (MJY1205) strains. The strains were grown overnight in liquid SC medium at 30°C, 10-fold serially diluted, spotted on SC plates, and incubated at 30°C or 37°C for 3 days. (B,C) HPLC analyses of nucleosides in total tRNA isolated from wild-type (MJY1150), sod1Δ (MJY1152), and ccs1Δ (MJY1163) cells. The y-axes show absorbance units (A.U.) at 254 (B) or 240 nm (C). (D,E) Effects of sod1Δ and ccs1Δ alleles on the levels of ncm5U, mcm5U, mcm5s2U, yWpA, and yW. The peak-area for the relevant nucleoside/dinucleotide (at 254 nm) was divided by the peak area for pseudouridine (Ψ), which serves as the loading control. The values represent the mean from three independent experiments. The standard deviation is indicated. Supplemental Table S1 includes values for other modified nucleosides. (*) P < 0.05, (**) P < 0.01, ns indicates not significant (P>0.05).
Sod1-deficient cells show defects in thiolation of U34 residues and in the formation of yW37
Although our analyses of total tRNA showed that the lack of Sod1 or Ccs1 leads to altered levels of mcm5s2U, mcm5U, and yW, it remained unclear if the phenotype is caused by a tRNA modification defect or if it reflects a change in the
relative abundance of the tRNAs harboring these nucleosides. To directly assess if the lack of Sod1 induces a tRNA modification
defect, we purified the mcm5s2U-containing 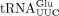 , mcm5U-containing
, mcm5U-containing 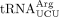 , and yW-containing
, and yW-containing 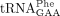 from the wild-type and sod1Δ strains. As a control, we purified the ncm5U-containing
from the wild-type and sod1Δ strains. As a control, we purified the ncm5U-containing 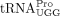 . Analyses of the nucleoside composition of the purified tRNAs, revealed that the levels of mcm5U in
. Analyses of the nucleoside composition of the purified tRNAs, revealed that the levels of mcm5U in 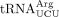 and ncm5U in
and ncm5U in 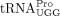 are unaffected by the sod1Δ allele (Supplemental Fig. S1). However, the analyses of
are unaffected by the sod1Δ allele (Supplemental Fig. S1). However, the analyses of 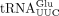 revealed that the lack of Sod1 causes a reduction in the levels of mcm2s2U and that the reduction is associated with a concomitant increase in the levels of mcm5U (Fig. 2A). These observations suggest that Sod1 influences the formation of the s2 group in mcm5s2U34-containing tRNA species. The analyses of
revealed that the lack of Sod1 causes a reduction in the levels of mcm2s2U and that the reduction is associated with a concomitant increase in the levels of mcm5U (Fig. 2A). These observations suggest that Sod1 influences the formation of the s2 group in mcm5s2U34-containing tRNA species. The analyses of 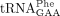 not only showed that the sod1Δ allele causes a reduction in the levels of yW, but they also revealed that the tRNA contained increased levels of m1G (Fig. 2B). The increased m1G levels suggest that the sod1Δ mutant is defective in the second step of yW synthesis, i.e., the conversion of m1G to 4-demethylwyosine (imG-14). Thus, the lack of Sod1 leads to reduced thiolation of U34 residues and a defect in the biosynthesis yW37.
not only showed that the sod1Δ allele causes a reduction in the levels of yW, but they also revealed that the tRNA contained increased levels of m1G (Fig. 2B). The increased m1G levels suggest that the sod1Δ mutant is defective in the second step of yW synthesis, i.e., the conversion of m1G to 4-demethylwyosine (imG-14). Thus, the lack of Sod1 leads to reduced thiolation of U34 residues and a defect in the biosynthesis yW37.

Sod1 deficiency is associated with distinct tRNA modification defects. (A,B). HPLC analyses of nucleosides in 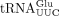 (A) and
(A) and 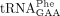 (B) isolated from wild-type (MJY1150) and sod1Δ (MJY1152) cells. The absorbance units (A.U.) were measured at 254 nm or 240 nm (yW). The chromatograms represent one of two
independently performed experiments, which showed comparable results.
(B) isolated from wild-type (MJY1150) and sod1Δ (MJY1152) cells. The absorbance units (A.U.) were measured at 254 nm or 240 nm (yW). The chromatograms represent one of two
independently performed experiments, which showed comparable results.
The tRNA modification defects in sod1Δ cells are partially suppressed by increased Ncs6/Ncs2 and Tyw1 expression
Since there is no evidence to suggest that Sod1 is directly involved in tRNA modification, it seemed likely that the tRNA modification defects in sod1Δ and ccs1Δ cells are a consequence of the reduced ability to detoxify superoxide. The formation of the s2 group as well as the conversion of m1G to imG-14 is dependent on proteins (Ncs6 and Tyw1) harboring iron-sulfur clusters (Noma et al. 2006; Liu et al. 2016). Since iron-sulfur clusters are sensitive to oxidation by superoxide (Imlay 2006), it seemed possible that the tRNA hypomodification in sod1Δ cells could be caused by reduced levels of active Ncs6/Ncs2 and Tyw1 enzymes. This model suggests that it may be possible to counteract the respective tRNA modification defect by increasing the expression of the Ncs6/Ncs2 and Tyw1 enzymes. Accordingly, the introduction of a high-copy plasmid harboring the NCS6 and NCS2 genes into sod1Δ cells was able to partially suppress the defect in mcm5s2U biosynthesis (Fig. 3A,C). Similarly, increased TYW1 dosage elevated the levels of yW/yWpA (Fig. 3B,D). Thus, the tRNA hypomodification in sod1Δ (and ccs1Δ) cells may be caused by reduced levels of active Ncs6/Ncs2 and Tyw1 enzymes.
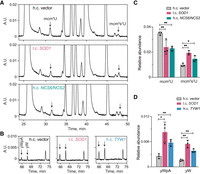
The tRNA modification defects in sod1Δ cells are partially suppressed by increased NCS6/NCS2 or TYW1 expression. (A,B) HPLC analyses of nucleosides in total tRNAs from sod1Δ cells (MJY1152) carrying the indicated high-copy (h.c.) or low-copy (l.c.) URA3 plasmid. The absorbance units (A.U.) were measured at 254 (A) or 240 nm (B). (C,D) Effects of the SOD1, NCS6/NCS2, and TYW1 plasmids on the abundance of indicated nucleosides/dinucleotide. The relative abundance was calculated by dividing the peak area of the relevant nucleoside/dinucleotide (at 254 nm) by the peak area for pseudouridine. The values represent the mean from three independent experiments. The standard deviation is indicated. (*) P < 0.05, (**) P < 0.01.
The tRNA modification defects of sod1Δ cells can be mimicked by exposing wild-type cells to paraquat
If the tRNA modification defects in sod1Δ cells are caused by superoxide-induced damage to the iron-sulfur clusters in Ncs6 and Tyw1, then a similar phenotype should be observed in wild-type cells grown in medium supplemented with the superoxide-generating chemical paraquat (PQ). To test this prediction, we cultured wild-type cells in the absence (control) or presence (0.25 or 2.5 mM) of PQ and analyzed the nucleoside composition of the total tRNA pool. We also analyzed the nucleoside composition of total tRNA isolated from ncs6Δ, tyw1Δ, and elp3Δ strains grown in the absence of PQ (Supplemental Fig. S2). The analyses revealed that wild-type cells grown in medium containing 0.25 mM PQ showed a tRNA modification pattern similar to the sod1Δ mutant, i.e., the tRNA contained elevated levels of mcm5U and reduced levels of mcm5s2U and yW/yWpA (Fig. 4; Supplemental Table S2). An even stronger phenotype was observed when the cells were grown in medium containing 2.5 mM PQ, which included a further reduction in the levels of mcm5s2U and a complete lack of yW/yWpA (Fig. 4; Supplemental Table S2). The presence of 2.5 mM PQ also caused significantly reduced levels of mcm5U and ncm5U, implying that the activity of the Elongator complex is reduced at the higher PQ concentration. Consistent with this notion, the Elp3 subunit of Elongator contains a radical-SAM domain and consequently a [4Fe-4S] cluster (Chinenov 2002; Dauden et al. 2019). Collectively, our results suggest that the tRNA modification defect of sod1Δ cells is caused by the inability to detoxify superoxide, and they imply that the phenotypes are caused by oxidative damage to iron-sulfur cluster-containing tRNA modifying enzymes.
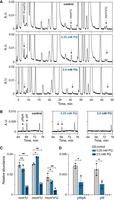
Effects of PQ on tRNA modification in wild-type cells. (A,B) HPLC chromatography of nucleosides from total tRNA isolated from wild-type (MJY1150) cells grown in the absence (control) or presence (0.25 or 2.5 mM) of PQ. The absorbance units (A.U.) were measured at 254 (A) or 240 nm (B). (C,D). Effects of PQ on the levels of indicated nucleosides/dinucleotide. The relative abundance was calculated by dividing the peak area of the relevant nucleoside/dinucleotide (at 254 nm) by the peak area for pseudouridine. The values represent the mean from three independent experiments. The standard deviation is indicated. Supplemental Table S2 includes values for other modified nucleosides. (*) P < 0.05, (**) P < 0.01, ns indicates not significant (P>0.05).
DISCUSSION
In both yeast and metazoans, sod1/SOD1 alleles have been reported to genetically interact with allelic variants of Elongator-subunit genes (Collins et al. 2007; Lin et al. 2008; Bento-Abreu et al. 2018). In this study, we show that Sod1-deficient yeast cells are partially defective in the formation of the s2 group in mcm5s2U34-containing tRNAs and in the biosynthesis of yW in 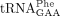 . Since null alleles of NCS6 or NCS2 show negative genetic interactions with mutations in genes for Elongator subunits (Björk et al. 2007; Nedialkova and Leidel 2015), the effect of the sod1Δ allele on the formation the s2 group can at least partially explain the genetic interaction between sod1 and elp alleles. The importance of the mcm5s2U34 and yW37 residues for translational efficiency and fidelity suggests that the tRNA modification defects may, on their own, contribute
to impaired proteostasis in Sod1-deficient cells.
. Since null alleles of NCS6 or NCS2 show negative genetic interactions with mutations in genes for Elongator subunits (Björk et al. 2007; Nedialkova and Leidel 2015), the effect of the sod1Δ allele on the formation the s2 group can at least partially explain the genetic interaction between sod1 and elp alleles. The importance of the mcm5s2U34 and yW37 residues for translational efficiency and fidelity suggests that the tRNA modification defects may, on their own, contribute
to impaired proteostasis in Sod1-deficient cells.
Inactivation of the yeast SOD1 gene generates auxotrophies for methionine and lysine, as well as a leaky leucine auxotrophy. While the methionine auxotrophy is likely caused by effects on cellular NADPH levels (Slekar et al. 1996), the lysine and leucine auxotrophies have been suggested to be the consequence of oxidative damage to the [4Fe-4S] clusters in the Lys4 and Leu1 biosynthetic enzymes (Wallace et al. 2004). Since Ncs6 and Tyw1 harbor iron-sulfur clusters and the tRNA hypomodification can be induced in wild-type cells by supplementing the medium with PQ, the tRNA modification defects in Sod1-deficient cells may well be caused by a superoxide-induced damage to the respective tRNA modifying enzyme. However, the lack of Sod1 induces a variety of metabolic changes (Montllor-Albalate et al. 2022), and it cannot be excluded that the effects on tRNA modification are more indirect. Although Fe/S clusters are generally sensitive to oxidative damage (Imlay 2006), the activity of the [4Fe-4S] cluster-containing Elongator complex is largely unaffected in Sod1-deficient cells. The activity of Elongator is, however, reduced in wild-type cells grown in medium supplemented with 2.5 mM PQ. It remains unclear whether the lack of an effect of the sod1Δ allele on the formation of xm5 groups is caused by a less solvent-exposed Fe/S cluster in Elongator or other features of the Elongator-dependent modification pathway.
In S. cerevisiae, the levels of 2-thiolated tRNAs are influenced by the growth conditions (Laxman et al. 2013; Alings et al. 2015), and it has been proposed that this modulation may play a role in regulating translation in response to environmental changes (Laxman et al. 2013). With respect to the availability of sulfur-containing amino acids, prototrophic S. cerevisiae cells exhibit lower levels of thiolated U34 residues when grown in minimal medium compared to when grown in minimal medium supplemented with methionine and cysteine (Laxman et al. 2013). Since cysteine is the sulfur donor for both the s2 group and Fe/S clusters, the precise mechanism by which the availability of sulfur-containing amino acids influences tRNA modification remains uncertain.
In both yeast and mammalian cells, the translation defects induced by the lack of wobble mcm5 or s2 groups have been linked to increased misfolding and aggregation of endogenous proteins (Nedialkova and Leidel 2015; Bento-Abreu et al. 2018; Rapino et al. 2018). In yeast, the lack of both groups has been found to aggravate the proteostasis defect (Nedialkova and Leidel 2015). ALS is characterized by protein aggregation in motor neurons and the importance of the mcm5s2U34 residue for proteostasis has provided a possible explanation to the increased ALS susceptibility of individuals with reduced ELP3 expression (Bento-Abreu et al. 2018). Cells with lower ELP3 levels and consequently lower mcm5s2U levels are expected to have intrinsically higher levels of protein misfolding and aggregation. Although this model may explain why ELP3 is a genetic modifier of ALS, our finding that Sod1 deficiency leads to reduced thiolation of U34 residues in tRNAs implies an additional level of complexity for at least some ALS-associated SOD1 alleles. Mutations in SOD1 that result in reduced superoxidase activity are expected to cause decreased thiolation of wobble uridines, which would, in turn, affect translation and proteostasis. The impact on translation and proteostasis would then be aggravated by reduced ELP3 levels. It should also be noted that high levels of oxidative stress impair the formation of Elongator-dependent xm5 groups. Further studies are needed to determine the combinatorial effects of variant SOD1 and ELP3 alleles on translation, proteostasis, and neurological dysfunction.
MATERIALS AND METHODS
Yeast strains, media, plasmids, and genetic procedures
Yeast strains used in this study are listed in Supplemental Table S3, and oligonucleotides in Supplemental Table S4. Synthetic complete medium was prepared as previously described (Amberg et al. 2005; Johansson 2017). The medium was, when required, supplemented with PQ (Sigma, #36541).
Strains deleted for SOD1 were constructed by transforming the diploid strain MJY1143 with a sod1::KanMX6 DNA fragment, which was PCR-amplified from pFA6a-KanMX6 (Longtine et al. 1998), with appropriate homologies. Following PCR confirmation, the generated heterozygous diploid was allowed to sporulate, and the wild-type (MJY1150 and MJY1151) and sod1Δ (MJY1152 and MJY1153) strains were obtained from tetrads. The ncs6::KanMX6 strain (MJY1191) was generated in a similar manner with the difference that the DNA fragment was transformed into MJY1191, which is a diploid formed between MJY1150 and MJY1151. The ccs1Δ (MJY1163), elp3Δ (UMY4563), and tyw1Δ (UMY4600) strains were also derived from heterozygous diploids, and to construct these strains the kanMX4 gene deletion cassette was PCR-amplified from the relevant strain of the haploid yeast deletion collection (Research Genetics). The sod1Δ elp3Δ double mutant (MJY1205) was derived from a cross between MJY1153 and UMY4563.
The pRS316-SOD1 and pRS426-TYW1 plasmids were constructed by amplifying the wild-type SOD1 and TYW1 genes from genomic DNA isolated from strain W303-1A, using the primer pairs OAB3538/OAB3539 and oMJ579/oMJ580, respectively. The SOD1 gene was cloned between the XhoI and XbaI sites of pRS316 (Sikorski and Hieter 1989), and TYW1 between the XhoI and EcoRI sites of pRS426 (Christianson et al. 1992). To construct the pRS426-NCS6/NCS2 plasmid, we first amplified the NCS6 gene using OAB1429 and OAB3558, and cloned it between the SalI and NotI sites of pRS426. The NCS2 gene was subsequently cloned into the SalI site of pRS426-NCS6 using a DNA fragment amplified with OAB1425/OAB1426. All plasmids were confirmed by DNA sequencing of the inserts.
HPLC analyses of tRNA-derived nucleosides
Cells were grown in freshly prepared SC medium at 30°C, and total tRNA was isolated as previously described from 100 OD600 units of exponentially growing cells (Chen et al. 2011). Typically, 50 µg tRNA was digested with nuclease P1 (N8630, Sigma-Aldrich) and bacterial alkaline phosphatase (P4252, Sigma-Aldrich), and the hydrolysate was analyzed by HPLC (Gehrke and Kuo 1990; Xu et al. 2018). The relative abundance of a modified nucleoside was calculated using pseudouridine (Ψ) as the loading control. Means between groups were compared using the unpaired two-tailed t-test in GraphPad Prism. In Supplemental Table S1, multiple testing correction was applied using the Benjamini–Hochberg procedure to obtain false discovery rate (FDR) values. Purification and analyses of single tRNA species were performed as described previously (Björk et al. 2001; Huang et al. 2005).
ACKNOWLEDGMENTS
We thank Artyom Egorov for valuable discussions. This work was supported by the Swedish Research Council (621-2016-03949 to A.S.B.), Karin and Harald Silvanders Foundation/Insamlingsstiftelsen Umeå universitet (FS 2.1.6-1870-16 to A.S.B.), Åke Wibergs Foundation (M14-0207 to M.J.O.J.), and Magnus Bergvalls Foundation (2017-02098 to M.J.O.J.).
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080181.124.
-
Freely available online through the RNA Open Access option.
- Received July 9, 2024.
- Accepted September 13, 2024.
This article, published in RNA, is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Fu Xu is the first author of this paper, “Sod1-deficient cells are impaired in formation of the modified nucleosides mcm5s2U and yW in tRNA.” Fu obtained his PhD degree under the supervision of Professor Anders Byström at Umeå University in Sweden, where he studied tRNA biogenesis in yeast. He is currently a postdoctoral fellow in the group of Professor Michaela Frye at the German Cancer Research Center (DKFZ) in Heidelberg, Germany, where he is investigating the role of RNA modifications in skin cancer and tissue regeneration.
What are the major results described in your paper and how do they impact this branch of the field?
In this paper, we demonstrate that the tRNA-modifying enzymes Nsc6/NSC2 and Tyw1, which contain iron-sulfur clusters, are inhibited in cells lacking the superoxide dismutase Sod1. Consequently, we show that the formation of mcm5s2U and yW in tRNAs is impaired in Sod1-depleted cells. SOD1 mutations are known to be associated with impaired proteostasis and ALS disease. Our findings reveal a novel role of Sod1 in tRNA biogenesis.
What led you to study RNA or this aspect of RNA science?
I developed an interest in RNA modifications while exploring the regulation of gene expression. Gene expression is controlled dynamically at multiple levels, from DNA replication to RNA turnover, with RNA modifications playing a vital role in this process.
If you were able to give one piece of advice to your younger self, what would that be?
I would say it is important to approach research with both realism and optimism. Don't get discouraged by failed experiments or unexpected results—they may lead to fascinating discoveries.
Are there specific individuals or groups who have influenced your philosophy or approach to science?
Yes, my approach to science has been greatly influenced by my PhD supervisor, Professor Anders Byström, and co-supervisor, Dr. Marcus Johansson, who have served as role models in research. Their passion for science and curiosity about understanding the molecular mechanisms behind various observations have shaped my own scientific mindset. I'm also deeply inspired by the optimism and efficiency of my postdoctoral supervisor, Professor Michaela Frye, who encourages me to take on challenging scientific questions.
What are your subsequent near- or long-term career plans?
My short-term career goal is to complete my postdoctoral research in Professor Michaela Frye's lab, where I am studying the role of RNA modifications in skin cancer and tissue regeneration. In the long term, I aim to establish my own research group focused on investigating the molecular mechanisms of human diseases, drawing on my expertise in RNA biology.








