
The small noncoding RNA Vaultrc5 is dispensable to mouse development
- Laboratory of Biochemistry and Molecular Biology, National Cancer Institute, The National Institutes of Health, Bethesda, Maryland 20892, USA
- Corresponding author: joana.vidigal{at}nih.gov
-
Handling editor: Fatima Gebauer
Abstract
Vault RNAs (vtRNAs) are evolutionarily conserved small noncoding RNAs transcribed by RNA polymerase III. Vault RNAs were initially described as components of the vault particle, but have since been assigned multiple vault-independent functions, including regulation of PKR activity, apoptosis, autophagy, lysosome biogenesis, and viral particle trafficking. The full-length transcript has also been described as a noncanonical source of miRNAs, which are processed in a DICER-dependent manner. As central molecules in vault-dependent and independent processes, vtRNAs have been attributed numerous biological roles, including regulation of cell proliferation and survival, response to viral infections, drug resistance, and animal development. Yet, their impact to mammalian physiology remains largely unexplored. To study vault RNAs in vivo, we generated a mouse line with a conditional Vaultrc5 loss-of-function allele. Because Vaultrc5 is the sole murine vtRNA, this allele enables the characterization of the physiological requirements of this conserved class of small regulatory RNAs in mammals. Using this strain, we show that mice constitutively null for Vaultrc5 are viable and histologically normal but have a slight reduction in platelet counts, pointing to a potential role for vtRNAs in hematopoiesis. This work paves the way for further in vivo characterizations of this abundant but mysterious RNA molecule. Specifically, it enables the study of the biological consequences of constitutive or lineage-specific Vaultrc5 deletion and of the physiological requirements for an intact Vaultrc5 during normal hematopoiesis or in response to cellular stresses such as oncogene expression, viral infection, or drug treatment.
Keywords
INTRODUCTION
Vault RNAs (vtRNAs) are small noncoding RNAs initially identified in rat livers (Kedersha and Rome 1986) through their association with the vault complex, the largest naturally occurring cellular particle described to date (Kedersha et al. 1986, 1991; Kedersha and Rome 1990; Kickhoefer et al. 1993). Both vaults and vtRNAs are evolutionarily conserved, and the particle itself is present at high copy numbers in many cellular contexts suggesting it has important functions in eukaryotes, though these remain to date poorly defined (van Zon et al. 2003; Hahne et al. 2021).
Although initially identified via its association with the vault, only a minority of the vtRNA is typically bound to that complex (Kickhoefer et al. 1998), and many alternative functions for this RNA have been proposed. In humans, vtRNAs are expressed from four related loci (vtRNA1-1, vtRNA1-2, vtRNA1-3, and vtRNA2-1) and are thought to serve as noncanonical precursors for miRNAs, whose processing is DICER-dependent but DROSHA-independent (Persson et al. 2009; Lee et al. 2011; Hussain et al. 2013; Sajini et al. 2019). Processing of vtRNAs into these miRNAs (also known as small-vaultRNAs; svtRNAs) seems to be a regulated process (Hussain et al. 2013; Sajini et al. 2019) suggesting they may act in a context-dependent manner. Studies in cell culture have suggested these svtRNAs are important for proper cell differentiation (Sajini et al. 2019), required for the regulation of cell cycle progression and apoptosis (Fort et al. 2018,2020), implicated in the regulation of drug metabolism (Persson et al. 2009), and dysregulated in both cancer (Xiong et al. 2011; Yu et al. 2014; Shen et al. 2018) and neural diseases (Somel et al. 2010). Unprocessed vtRNAs from both species have also been implicated in the regulation of autophagy (Horos et al. 2019; Buscher et al. 2022), PKR activity (Lee et al. 2011), apoptosis (Amort et al. 2015), lysosome biogenesis (Ferro et al. 2022), and viral particle trafficking (Saruuldalai et al. 2022). Surprisingly, despite the mounting evidence implicating both full-length and processed vtRNAs in the regulation of fundamental biological processes, their physiological requirements are not known.
In contrast to humans, mice have a single vtRNA (Vaultrc5 or Mvg1) (Stadler et al. 2009). The existence of a sole vtRNA gene in murine genomes represents an invaluable opportunity to define the physiological requirements of an abundant but mysterious class of noncoding RNA molecules in mammals. With this purpose, we have generated a new mouse model carrying a conditional loss-of-functional allele for Vaultrc5 and performed the initial characterization of Vaultrc5-null animals. We show that constitutive loss of Vaultrc5 in vivo is compatible with animal development and survival. We also find that Vaultrc5-null animals are histologically indistinguishable from their wild-type littermates with no detectable gene expression changes in either livers or brains, two organs where vtRNAs have been proposed to play important functions. Nevertheless, we have observed a minor reduction in platelet counts in the absence of Vaultrc5, suggesting vtRNAs may have important roles in hematopoiesis. Our work represents the first step toward defining the functions of mammalian vtRNAs in vivo. This mouse model will enable more detailed studies using targeted and acute deletions of this conditional allele to further define the physiological roles of vtRNAs during tissue development and homeostasis or in response to viral infections, oncogenic insults, or other cellular stresses.
RESULTS
Analysis of noncanonical miRNAs derived from the murine Vaultrc5 locus
Mouse genomes encode a sole vtRNA gene, Vaultrc5 (also known as Mvg1) (Kickhoefer et al. 2003). Vaultrc5 is transcribed as a single exon-transcript from Chromosome 18 (18qB2) from a locus located immediately downstream from Zmat2 (Supplemental Fig. S1A), and that is in a syntenic region conserved across mammals (Stadler et al. 2009). All vtRNAs are characterized by their ability to associate with the vault particle, are often transcribed from analogous genomic locations, and have been described in eukaryotes ranging from amoebas to humans. However, the Vaultrc5 locus shows relatively low sequence conservation to other vertebrates or placental mammals when compared to that of protein-coding genes (Supplemental Fig. S1A). Nevertheless, a few features of the locus show some conservation. First, like vtRNAs of other species, the regulatory elements in its polymerase III (pol III) promoter are well defined (Supplemental Fig. S1B). These include a proximal element and a TATA-like box (Kickhoefer et al. 2003). Both of these are also found in the promoters of human vtRNA1 genes (vtRNA1-1, vtRNA1-2, and vtRNA1-3) (Kickhoefer et al. 2003), which like Vaultrc5 are located downstream from Zmat2. Moreover, at least the proximal element shows broad conservation among placental mammals and vertebrates as does the polymerase III termination sequence (Supplemental Fig. S1B) (see also Stadler et al. 2009). Within the gene body itself, Vaultrc5 contains three regulatory sequences in the form of the A and B boxes (Kickhoefer et al. 1993), which characterize pol III type 2 promoters such as those found in tRNA genes (Dieci et al. 2007). Only two of these (A box and B2 box) are broadly conserved, as are the sequences that surround them (Supplemental Fig. S1B). In contrast, the central region of vtRNAs varies substantially between species (Stadler et al. 2009). In the case of Vaultrc5, it contains a third pol III internal sequence known as B1 box, with low overall conservation with other species (Supplemental Fig. S1B) with the notable exception of rats (Kickhoefer et al. 2003).
Despite these differences, the 3′ and 5′ ends of Vaultrc5, are predicted to form a double-stranded region that is identical to that predicted for human vtRNA1 genes, as exemplified here by vtRNA1-1 (Fig. 1A). Importantly, although the sequences predicted to fold into this double-stranded structure are highly conserved between vtRNAs of the two species (Supplemental Fig. S1C), they share only 83%–89% of nucleotide identity. Yet, every position at which the sequence of the mouse Vaultrc5 differs from that of one or more vtRNA1 genes, the change either has little impact on base-pairing (e.g., G·C pairing between bases 3 and 136 of Vaultrc5 vs. G·U wobble pairing between bases 3 and 92 in vtRNA1-1) or is compensated by changes in the complementary RNA sequence (Fig. 1A; Supplemental Fig. S1C). This suggests that the secondary structure of the stem may play important roles for the functions of vtRNAs, and that those are conserved between mice and humans. Given that for human vtRNA1-1 this stem is processed by DICER into noncanonical miRNAs (Fig. 1B; Persson et al. 2009), we thought it was likely that identical molecules are produced from Vaultrc5 as well (Supplemental Fig. S2A). To investigate this possibility, we analyzed small RNA sequencing data sets from a variety of adult mouse tissues (Kern et al. 2020). We found that reads matching the predicted location of the mature miRNA sequences were relatively low abundant (Fig. 1C,D; Supplemental Fig. S2B-D). Of all four predicted noncanonical miRNAs derived from this locus, svtRNAa* had the highest number of reads. Nonetheless, they were much less abundant than those matching canonical miRNAs. As an example, miR-19a-3p a miRNA with functions in hematopoietic tissues reached about 9000 RPM in the bone marrow and spleen samples, with a mean of around 2600 RPM across all tissues (Fig. 1D). In contrast, reads for the predicted vaultRNA-derived miRNA-like fragments had an abundance similar to that of miR-19a-5p, the passenger strand of miR-19a, which is degraded following loading of the mature miRNA into an Argonaute protein (miR-19a-5p max = 39.3 RPM, mean = 6.3 RPM; svtRNAa max = 5.2 RPM, mean = 0.7 RPM; svtRNAa* max = 130.8 RPM, mean = 20.7; svtRNAb max = 3.8 RPM, mean = 0.5 RPM; svtRNAb* max = 3.8 RPM, mean = 0.5 RPM). Low counts were observed even in the lung, where the levels of vaultRNA-derived miRNAs were reported to be high in humans (mean counts svtRNAa = 0.4 RPM, svtRNAa* = 25.5 RPM, svtRNAb = 0.2 RPM, svtRNAb* = 0.2 RPM) (Persson et al. 2009). Given that the strength of repression imparted by a miRNA depends strongly on its abundance (Mukherji et al. 2011) and that even well-expressed miRNAs like miR-19a have typically only modest effects on target gene expression, these data suggest that under homeostatic conditions, Vaultrc5 is not a source of noncanonical miRNAs with physiologically relevant functions in the tissues we have analyzed.
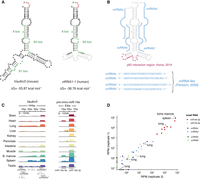
Structure and conservation of the murine Vaultrc5 locus. (A) Minimum free energy secondary structure prediction for Vaultrc5 and vtRNA1-1 transcripts. The position of the internal regulatory elements is highlighted. The sequences that are not conserved between the mouse and human transcripts are shown in gray in the vtRNA1-1 structure. (B) Position of known functional elements on vtRNA1-1, including the noncanonical miRNAs (blue, sequences shown on the bottom) and p62 interaction region (magenta). (C) Genome browser view of sequencing reads over Vaultrc5 and pre-mmu-miR-19a loci in multiple adult mouse tissues. (D) Expression levels of the predicted Vaultrc5-derived miRNAs compared to those of miR-19a (miR-19a-3p) and its passenger strand (mir-19a-5p) across two sequencing replicates.
Previous work has suggested that in humans, svtRNAb plays a role in drug resistance through the regulation of target genes such as CYP3A4 (Persson et al. 2009). Based on this, it is possible that vtRNA-derived miRNAs perform conserved regulatory functions only under stress conditions. We think that is unlikely to be the case. First, miRNAs recognize their targets primarily through base-pair complementarity via their extended seed regions (miRNA nt 2–8) (Bartel 2009). Although the sequence variations between the vtRNA-1 and Vaultrc5 genes preserve the secondary structure of the vaultRNA stem, they do not preserve the seed of all predicted svtRNAs, specifically svtRNAa and svtRNAb* (Supplemental Fig. S2A). Second, although the predicted seed sequences of svtRNAb and svtRNAa* is shared between the two species, there is poor overlap between their predicted targets, based on our own implementation of the targetscan algorithm (Supplemental Table S1; Lewis et al. 2005).
Generation of a Vautrc5 conditional loss-of-function allele
Despite the lack of evidence that Vaultrc5 functions as a noncanonical precursor for murine miRNAs, the conserved secondary structure of its stem across species suggests this noncoding RNA plays important functions in vivo. In fact, vtRNAs have been functionally implicated among other processes in cell survival (Amort et al. 2015; Bracher et al. 2020), proliferation (Ferro et al. 2022), and differentiation (Skreka et al. 2012; Sajini et al. 2019), all of which are essential for successful embryonic development. The expression of full-length vtRNAs also seems to be regulated during cellular differentiation (Skreka et al. 2012). Together, these reports suggest that full-length vtRNAs may be essential during mammalian embryogenesis. To help define what those functions might be, we generated a conditional loss-of-function allele that would allow both constitutive as well as spatiotemporally controlled deletion of the locus, enabling the dissection of the physiological requirements for Vaultrc5 expression in mice. We designed a construct in which loxP sites were inserted directly upstream and downstream from the annotated promoter and termination elements, respectively, without disrupting them (Fig. 2A; Supplemental Fig. S1B). Both loxP sequences were inserted at genomic sites with no sequence conservation among vertebrates or placental mammals, suggesting they do not unintentionally disrupt regulatory elements of neighboring genes. Moreover, no conserved sequences aside from those of Vaultrc5 are predicted to be affected by Cre-mediated recombination of the loxP sites. Of note, putative enhancer elements as well as CTCF-binding sites are found in the vicinity of the Vaultrc5 but, as above, none are affected by our targeting or by the subsequent CRE-mediated genomic deletion. To facilitate the genotyping of the Vaultrc5 locus following targeting, we included an XmnI restriction site next to each loxP sequence (Supplemental Fig. S1D).
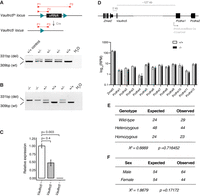
A conditional loss-of-function allele for Vaultrc5. (A) Schematic representation of conditional Vaultrc5 locus (Vaultrc5flx) and its recombination after Cre expression to generate a null allele. The position of the three primers used for monitoring the recombination is highlighted. When used in combination, these result in amplicons of 309 base pairs (bp) for wild-type alleles (P1/P2) and 331 bp for recombined alleles (P1/P3). An example gel is shown on the bottom, showing the recovery of heterozygous null animals (+/−) following β-actin-Cre expression. (B) Genotyping PCR showing the recovery of wild-type (+/+), heterozygous (+/−), and Vaultrc5-null (−/−) animals from Vaultrc5+/− intercrosses. (C) Relative levels of Vaultrc5 in mouse embryonic fibroblasts isolated from wild-type, heterozygous, and homozygous null animals. (D, top) Schematic representation of the Vaultrc5 locus and its distance to its conserved upstream and downstream neighbors. (Bottom) Expression of Vaultrc5 flanking genes in brains from wild-type and null animals. (E) Observed and Expected numbers for wild-type (+/+), heterozygous (+/−), and Vaultrc5-null (−/−) animals obtained from Vaultrc5+/− intercrosses. P-value was calculated with a χ2 test. (F) As in E but for animal sex.
Constitutive loss of Vautrc5 is compatible with mouse development
To test the broad requirement for vtRNA expression during mouse development, we crossed Vaultrc5flx/+ mice to the general β-actin-Cre deleter line (Lewandoski et al. 1997). This led to widespread recombination of the floxed alleles and to offspring that were heterozygous for a Vaultrc5 null allele (Vaultrc5−/+; Fig. 2A). Subsequent intercrossing of heterozygous Vaultrc5−/+ mice generated full Vaultrc5 loss-of-function animals, carrying the null allele in homozygosity (Vaultrc5−/−) (Fig. 2B). Quantification of Vaultrc5 in cells from these animals by RT-qPCR confirmed a genotype-dependent reduction of Vaultrc5 RNA levels with no detectable transcripts in Vaultrc5−/− cells (Fig. 2C). In contrast, the expression of Zmat2 and Pcdha1 was not altered in these samples (Fig. 2D), confirming that our targeting strategy did not disrupt their regulation.
The recovery of Vaultrc5−/− animals at weaning suggests that complete loss of vtRNA expression in mice is compatible with animal development. Nevertheless, many genes that have well-established essential functions during embryogenesis—including those in the canonical miRNA pathway—lead to lethal phenotypes with incomplete penetrance when disrupted (Wang et al. 2008; Van Stry et al. 2012; Vidigal and Ventura 2015; Sala et al. 2020). We found no evidence of that being the case for Vaultrc5. Specifically, data collected over multiple litters showed that Vaultrc5−/− animals were recovered at the expected ratio at weaning (Fig. 2E). Similarly, we found no deviation from the expected ratios between sexes (Fig. 2F). Together, these data indicate that mice are able to successfully complete development and survive in the absence of vtRNAs.
Vaultrc5-null mice are histologically normal
Aside from being viable, Vaultrc5−/− animals were morphologically indistinguishable from littermate controls, suggesting that the constitutive absence of vtRNAs does not lead to gross phenotypic abnormalities in mice. Furthermore, we found no weight differences—a common indicator of suboptimal animal health—between genotypes either in male (median weights of 18.25 g, 18.05 g, 18.55 g for wild-type, heterozygous, and homozygous, respectively) or female animals (median weights of 15.75 g, 15.9 g, 15.8 g for wild-type, heterozygous, and homozygous, respectively) (Fig. 3A).
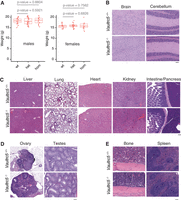
Vaultrc5-null mice are morphologically and histologically indistinguishable from wild-type littermates. (A) Weight of male (left) and female (right) animals of the indicated genotypes at weaning, showing no difference between Vaultrc5-null mice and littermate controls. Each dot represents an animal. P-values were calculated with a two-tailed t-test. (B–E) Hematoxylin and Eosin-stained histology sections from organs collected from Vautrc5+/+ and Vautrc5−/− sex-matched littermates at 8 weeks of age.
To test if the loss of Vaultrc5 caused more subtle phenotypes, we collected tissues from Vaultrc5−/− and Vaultrc5+/+ littermates of both sexes at 8 weeks of age and subjected them to a histopathological evaluation (Figs. 3B–3E; Supplemental Note S1). We were particularly interested in brain tissues as previous studies had implicated vtRNAs in neuronal differentiation (Wakatsuki et al. 2021a,b). In line with a potential role for vtRNAs in the brain, their dysregulation has also been implicated in neurodegenerative diseases (Minones-Moyano et al. 2013). Despite these reports, we found that in the absence of Vaultrc5, the neural tissues were seemingly uncompromised. Specifically, cortex, medulla, hippocampus, brain stem, corpus callosum, and cerebellum all showed no histological differences between Vaultrc5−/− and Vaultrc5+/+ animals (Fig. 3B). Meninges were present on the cortex, and the parenchyma of the gray and white matter consisted of fine capillaries, glial cells, neurons, and abundant neuropil. The cerebellum of mutant animals was equally unremarkable and characterized by a very cellular granular layer and a less cellular molecular layer, with the prominent Purkinje cells at the interface of the two layers. Finally, the delicate pia mater lined the cerebellum along the molecular layer in animals of both genotypes. In sum, we found no evidence that constitutive loss of vtRNA impacts neural tissues in mice, with the histologic findings in both Vaultrc5-null and wild-type controls being within normal limits and consistent with normal brain development and tissue differentiation.
In addition to neural tissues, Vaultrc5 may also impact liver biology since vtRNAs were initially identified as components of the vault particle from rat liver extracts (Kedersha and Rome 1986), an organ in which the murine Vaultrc5 is well expressed (Kickhoefer et al. 2003). Moreover, this organ is characterized by high levels of autophagy (Ueno and Komatsu 2017), a process that seems to be regulated by vtRNAs through their interaction with p62 via the central region (Fig. 1B; Horos et al. 2019). Although the predicted structure of this region has limited similarity between vtRNA1-1 and Vaultrc5 (Fig. 1A), the binding of p62 to vtRNAs seems to be conserved between the two species (Horos et al. 2019). Yet, despite the importance of autophagy to hepatic functions (Ueno and Komatsu 2017), histopathological analysis showed no evidence that the development or structure of the liver was compromised in the absence of Vaultrc5. Specifically, the liver in animals of both genotypes was encapsulated and composed of hepatocytes with abundant granular pink cytoplasm. In both cases, the organ was organized as cords of cells with a zonal arrangement around both portal triads and central veins. Macrophages and scattered mononuclear cells were also identified throughout the parenchyma (Fig. 3C). Similarly, in our analysis of other major organs, including the lung, heart, kidney, spleen, intestine, pancreas (Fig. 3C) as well as ovaries and testis (Fig. 3D), we found no evidence of histological differences between Vaultrc5−/− and Vaultrc5+/+ animals. A detailed description of the histological findings in these organs can be found in Supplemental Note S1.
Finally, we analyzed the histology of bone and spleen sections in our mice as these are major hematopoietic and lymphoid organs, and vtRNAs have been previously implicated in immune cell functions (Mrazek et al. 2007; Amort et al. 2015; Li et al. 2015). We found no histological differences between the spleens of Vaultrc5 wild-type and Vaultrc5 loss-of-function animals (Fig. 3E). Similarly, bones of wild-type and mutant animals were histologically identical (Fig. 3E). In both cases, bone marrows showed high cellularity and tri-lineage hematopoiesis. Within this cell population, megakaryocytes were the most abundant cells with erythroid and granulocytic precursors easily observed at higher magnifications. In all cases, these cells showed no morphological abnormalities in the absence of Vaultrc5.
As part of our pathology analysis, we also performed a complete blood count. We found that measurements for Vaultrc5−/− animals as well as for Vaultrc5+/+ littermate controls were within the normal physiological ranges expected for the C57Bl/6 strain (Mouse Phenome Database, www.jax.org/phenome). They were also for the most part identical between wild-type and mutant mice with two notable exceptions (Supplemental Fig. S3). First, although we found no statistical difference in white and red blood cell counts (Supplemental Fig. S3A,B), red blood cell size (Supplemental Fig. S3E,), or hemoglobin levels (Supplemental Fig. S3G-I) between Vaultrc5−/− and Vaultrc5+/+ animals, hematocrit values—the percentage of red blood cells in the blood—were slightly reduced in mutant mice (Supplemental Fig. S3D). This suggests that Vaultrc5 may have essential albeit subtle roles in erythropoiesis. Second, the platelet counts were reduced in the absence of Vaultrc5 to ∼78% of the values in wild-type animals (Supplemental Fig. S3C). This may point to a potential role for Vaultrc5 in platelet development from megakaryocyte precursors in the bone marrow or in the regulation of platelet survival. Nevertheless, platelet values remained well within the normal physiological ranges expected for the C57Bl/6 strain (Vaultrc5+/+ = 977–997 × 103 platelets/μL; vautrc5−/− = 745–792 × 103 platelets/μL; C57Bl/6 = 562–2159 × 103 platelets/μL).
RNA sequencing profiles of wild-type and Vaultrc5−/− animals
We reasoned that even in the absence of major histological changes, loss of Vaultrc5 could result in molecular phenotypes that would be reflected in transcriptome dysregulation. To determine if that was the case, we performed RNA sequencing to the brain and liver of vautrc5−/− animals, alongside those of age- and sex-matched wild-type littermate controls (Fig. 4). As before, we chose to focus on these organs because previous studies pointed to a role for vtRNA in their development or homeostasis. For example, in vitro studies of both human vtRNA1-1 (Wakatsuki et al. 2021a) and mouse Vaultrc5 (Wakatsuki et al. 2021b) promote synapse formation by modulating MAPK signaling and deregulation of the human vtRNA2-1 was proposed to induce neural dysfunction in humans (Minones-Moyano et al. 2013). Despite this, we found no evidence of gene dysregulation caused by the absence of vtRNA in mouse brains (Fig. 4A). Similarly, human vtRNA1-1 was shown to promote cell proliferation and tumorigenesis in human liver cell lines (Ferro et al. 2022), which together with the abundant expression of vtRNAs in the liver (Kickhoefer et al. 2003) suggests a potential role for vtRNAs in this organ. Yet, we again found no gene significantly dysregulated in the Vaultrc5−/− samples (Fig. 4B). Finally, vtRNA loss did not significantly affect the expression of Mvp, Vparp, or Tep1, the other components of the vault particle in the two tissues we have analyzed. Together with the histological analysis of these organs, these data suggest that in the mouse, vtRNA has no significant impact to the development or homeostasis of the brain or liver.
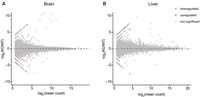
RNA sequencing profiles of Vautrc+/+ and Vautrc5−/− animals. MA plots showing lack of differentially expressed genes between brains (A) and livers (B) of wild-type and Vaultrc5-null animals. Differentially expressed genes were defined as those with log2 fold-change > [2] and adjusted P-value < 0.1.
DISCUSSION
We describe here the generation of Vaultrc5 conditional knockout mice and the initial characterization of animals with constitutive deletion of this locus. Given that mice—like most mammals—have a single vtRNA1 gene and have in addition lost vtRNA2 (Stadler et al. 2009), Vaultrc5−/− animals are null for vtRNAs. This is to our knowledge the first characterization of the physiological requirements for this evolutionarily conserved but poorly understood class of small RNAs in vertebrates.
Despite the myriad of roles assigned to vtRNA genes in humans, ranging from regulation of cell proliferation, survival, and differentiation, we found that animals null for Vaultrc5 are viable and histologically normal. This suggests that vtRNAs may not be essential in mice under unchallenged conditions. This is similar to what has been described for the other components of the vault particle. Indeed, mice null for the Major vault protein (Mvp−/−), the telomerase-associated protein 1 (Tep1−/−), or the vault poly-(adenosine diphosphate-ribose) polymerase (Vparp−/−) are all seemingly phenotypically normal (Kickhoefer et al. 2001; Mossink et al. 2002; Liu et al. 2004) even when multiple genes are deleted (Liu et al. 2004). Yet, when challenged with specific pathogens Mvp−/− animals show increased resistance to infection (Kowalski et al. 2007). Similarly, Vparp−/− animals show increased susceptibility to carcinogen-induced tumorigenesis (Raval-Fernandes et al. 2005). Thus, as is the case for many other genes, it may be that vtRNA expression only becomes indispensable under particular conditions such as aging, oncogenic stress, or viral infections all of which have been previously linked to vtRNA functions. It is also important to note that human genomes are unique in that they express four vtRNA paralogs, unlike the majority of other mammals, which like mouse have a single vtRNA gene. Given this, it is formally possible that vtRNAs have acquired unique functions in human cells, which are absent in the mouse.
Still, hematopoiesis may be particularly sensitive to loss of vtRNAs since we have observed a small but significant reduction in the platelet counts in our animals compared to littermate controls. Follow-up studies breeding the conditional Vaultrc5fx/flx animals with lineage-restricted deleters such as Vav-Cre (leading to recombination of the locus in all cells of the hematopoietic system) (Stadtfeld and Graf 2005) or Gp1ba-Cre (leading to recombination in megakaryocytes) (Nagy et al. 2019) combined with both immune challenges and a more detailed characterization of the immune system will be essential to fully characterize these phenotypes, define the extent to which Vautrc5 is required for hematopoiesis, and understand the predominant mechanism through which this evolutionarily conserved small noncoding RNA regulates mammalian physiology. To facilitate those studies, we are making our mice freely available to the community through The Jackson Laboratory mouse strain repository.
MATERIALS AND METHODS
Mouse husbandry and transgenic lines
Mouse β-actin-Cre line has been previously described (Lewandoski et al. 1997) and was obtained from the Jackson Laboratory (strain 019099). Vaultrc5flx mice (carrying loxP sites around Vaultrc5 gene) will be made available through the Jackson Laboratory as JAX Stock No. 037602. These animals were generated by zygotic injection of a single-stranded donor DNA template ordered from IDT and in vitro assembled Cas9-gRNA ribonucleoprotein complexes. Reagents were generated and tested by the Genome Modification Core at Frederick National Laboratory for Cancer Research and used in targeting experiments by the Mouse Modeling & Cryopreservation Core. Super-ovulated C57Bl6NCr female mice were used as embryo donors. Animals were genotyped using PCR followed by Restriction Fragment Length Polymorphism (RFLP). The 3′ loxP site or the 5′ loxP were amplified using (3′loxP, 5′-GAATCCGCGGAACTTTGG-3′, 5′-AATGCATACACAGGAGAGTTTCA-3′; 5′loxP, 5′-AGGCAACCCATCTCTTATT-3′, 5′-GAGATGACAGACCAATCGG-3′), which amplify a 1140-bp band from (3′ loxP) or a 1113-bp band (5′ loxP). PCRs were cleaned up and digested with Xmnl for 1 h. The resulting fragments were visualized by agarose gel electrophoresis. Correct integration in the founder male was confirmed by amplifying genomic DNA from tail clippings using primers flanking the targeting construct (5′-AGGCAACCCATCTCTTATT-3′, 5′-TGCATGTTAAAAACCCTCAGAAC-3′), cloning the resulting amplicon into the TOPO vector, followed by Sanger sequencing of multiple clones. A colony for this mouse strain was established by crossing the founder to C57Bl6 females obtained from The Jackson Laboratory (Stock No: 000664). All animal procedures were conducted according to the NIH Guide for the Care and Use of Laboratory Animals, under Animal Study Proposal no. 390923 approved by relevant National Institutes of Health Animal Care and Use Committees.
Conservation tracks
Vertebrate and placental mammal basewise conservation tracks were generated by PhyloP, downloaded from the UCSC Genome Browser, and visualized on IGV.
RNA secondary structure predictions
Minimum free energy structure predictions were computed with RNAfold v2.4.18 (Mathews et al. 2004) using the parameters “RNAfold -p -d2 ‐‐noLP < sequence1.fa > sequence1.out.”
TargetScan predictions
Data sets and Perl scripts were used from the 7.2 release of TargetScanMouse and TargetScanHuman (Lewis et al. 2005). First, the conserved miRNA targets and nonconserved sites were identified using a custom set of data available on TargetScan for both mouse and human (Garcia et al. 2011; Agarwal et al. 2015). Second, conserved branch length (sum of phylogenetic branch lengths between species that contain a site) and PCT (probability of preferentially conserved targeting) for each predicted target in a custom set of data was calculated (Friedman et al. 2009). Third, the context++ scores for a set of predicted miRNA sites in a custom set of data were calculated. The context++ score for a specific site is the sum of the contribution of 14 features described in Nam et al. (2014). A list of targets with predicted miRNA binding sites in their 3′ UTRs was generated based on conserved branching length, PCT and context++ score. The entire analysis was done on Homo sapiens, Mus musculus, Macaca mulatta, Pan troglodytes, Bos taurus, Rattus norvegicus, and Monodelphis domestica, which were available on TargetScan.
Histology and blood collection
Eosin-hematoxylin staining was performed on 5 μm sections of tissues fixed in 10% neutral buffered formalin and embedded in paraffin. Blood was collected by cardiac puncture in anesthetized animals. Animals were euthanized immediately upon completion of blood collection in K2-EDTA collection tubes (Thermo Scientific). Complete blood counts as well as histopathology analysis were performed at MD Biosciences.
Cell lines and cell culture
Mouse embryonic fibroblasts (MEFs) were derived using standard protocols. Briefly, embryos were isolated from Vaultrc5+/− intercrosses at embryonic day (E) 13.5. After removal of internal organs, embryo carcasses were minced and digested with trypsin at 37°C before enzyme inactivation with DMEM Media (GIBCO) supplemented with 10% FBS, penicillin/streptomycin (100 U/mL), and L-glutamine (2 mM). For each embryo, the resulting cell suspension was plated on a 10 cm dish. Once confluent, these primary cells were frozen as passage 1. To generate immortalized cell lines, early passage primary MEFs from all genotypes were infected in parallel with the SV40 large T antigen (Addgene:13970) (Zhao et al., 2003). MEFs were maintained at 37°C (5% CO2) in DMEM media (GIBCO) supplemented with 10% FBS, penicillin/streptomycin (100 U/mL), and L-glutamine (2 mM).
RNA isolation and quantitative RT-PCR
Quantitative real-time RT-PCR (qPCR) was performed on RNA isolated using TRIzol reagent (Invitrogen), using standard methods. RNA was treated with ezDNase (Thermo Fisher Scientific) and reverse transcribed using SuperScript IV Reverse Transcriptase (Thermo Fisher Scientific) using random hexamer primers. Relative expression of Vaultrc5 and Gapdh was quantified using SYBR Green (Thermo Fisher Scientific) and the following primer sets: Vaultrc5, 5′-AGCTCAGCGGTTACTTCGAC-3′, 5′-TCGAACCAAACACTCACGGG-3′; Gapdh 5′-AGGTCGGTGTGAACGGATTTG, 5′-TGTAGACCATGTAGTTGAGGTCA-3′.
Small RNA sequencing analysis
Small RNA-seq data for different mouse tissues were downloaded from GEO DataSets (GSE119661) (Kern et al. 2020). Adapter sequence TGGAATTCTCGGGTGCCAAGG was trimmed using parameters; -e 0.1 -n 1 -O 3 -q 20 -m 15 -M 25. Trimmed reads were mapped to customed noncoding RNA genome (Mus_musculus.GRCm39.ncrna) using STAR aligner (2.7.11b) with ‐‐outFilterMismatchNmax 1 \ ‐‐outFilterMismatchNoverLmax 1 \ ‐‐outFilterMismatchNoverReadLmax 1 \ ‐‐outFilterMatchNmin 16 \ ‐‐outFilterMatchNminOverLread 0 \ ‐‐outFilterScoreMinOverLread 0 \ ‐‐outFilterMultimapNmax 5000 \ ‐‐winAnchorMultimapNmax 5000 \ ‐‐seedSearchStartLmax 30 \ ‐‐alignTranscriptsPerReadNmax 30000 \ ‐‐alignWindowsPerReadNmax 30000 \ ‐‐alignTranscriptsPerWindowNmax 300 \ ‐‐seedPerReadNmax 3000 \ ‐‐seedPerWindowNmax 300 \ ‐‐seedNoneLociPerWindow 1000 \ ‐‐outFilterMultimapScoreRange 0 \ ‐‐alignIntronMax 1 \ ‐‐alignSJDBoverhangMin 999999999999. Bam and Bigwig files were generated using samtools (v 1.19) and exported to Integrative Genomics Viewer (2.16.2) to visualize reads to the Vaultrc5 and mmu-miR-19a loci. Expression of svtRNAs and mmu-miR-19a were represented in reads per million (RPM).
Total RNA sequencing
RNA from brains and livers from Vaultrc5+/+ and Vaultrc5−/− mice was isolated using TRIzol reagent (Invitrogen) following the manufacturers’ protocol. RNA was then treated with DNase I using Zymo RNA Clean & Concentrator kit. Libraries were constructed using the Illumina Stranded Total RNA Prep, ligation with Ribo-Zero Plus, and sequenced with stranded sequencing on NextSeq2000. Reads were trimmed using cutadapt (v 4.7) for all Illumina adapters with parameters; ‐‐nextseq-trim = 20 ‐‐trim-n -n 5 -e 0.1 -O 3 -q 20,20 -m 25, then trimmed reads were mapped to Mus_musculus. GRCm39 genome with Ensemble Genes 112 using ‐‐outFilterMultimapNmax 25 \ ‐‐winAnchorMultimapNmax 50 \ ‐‐alignTranscriptsPerReadNmax 10000 \ ‐‐alignWindowsPerReadNmax 10000 \ ‐‐alignTranscriptsPerWindowNmax 100 \ ‐‐seedPerReadNmax 1000 \ ‐‐seedPerWindowNmax 50 \ ‐‐seedNoneLociPerWindow 10 \ ‐‐outFilterMultimapScoreRange 1 \ ‐‐outFilterMismatchNoverLmax 0.3 \ ‐‐sjdbScore 2 using STAR aligner (2.7.11b). Read alignments were counted using Subread featureCounts (v 2.0.3) with -s 0. Differentially expressed genes were analyzed by using DEseq2 (R 3.6.0).
Statistical analysis
Statistics were done using R v4.1 (R Core Team 2018).
DATA DEPOSITION
RNA sequencing data sets have been deposited to GEO and are available under GSE269048.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
We thank all members of the Vidigal and Batista labs for discussions and comments on this work. We thank Raj Chari and Parirokh Awasthi for the generation of the Vaultrc5 allele. This work was supported by the Intramural Research Program of the National Institutes of Health through the Center for Cancer Research, National Cancer Institute, project 1ZIABC011810 (J.A.V.).
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080161.124.
-
Freely available online through the RNA Open Access option.
- Received June 25, 2024.
- Accepted August 5, 2024.
This is a work of the US Government.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Mahendra Prajapat is the first author of this paper, “The small noncoding RNA Vaultrc5 is dispensable to mouse development.” Mahendra obtained his PhD in Systems Biology from the Department of Chemical Engineering, IIT Bombay, India in 2015, where he studied gene expression dynamics using computational tools and mathematical models. Thereafter, he continued his research as a short-term postdoctoral researcher at the Indian Institute of Science, Bangalore, India and Tampere University, Finland. In November 2018, he joined the National Cancer Institute, National Institutes of Health USA. He had computational expertise and was determined to learn wet-lab techniques as well as mouse genetics. He has now generated four different mouse models, one of which is described in the current article. These genetic models support his current research interests of understanding the role of noncoding RNAs in gene regulation and how it alters phenotypic outcomes.
What are the major results described in your paper and how do they impact this branch of the field?
Several studies have shown vaultRNA's role in regulating a number of critical cellular processes using human and mouse cell lines. Interestingly, some observations even suggested that these RNAs could be involved in riboregulation of autophagy, regulation of drug metabolism during cancer treatment or serve as precursors of noncanonical miRNAs. Therefore, I decided to knock out the vaultRNA gene in mouse and dissect how its absence impacts animal physiology in vivo. Surprisingly, I found that the loss of vaultRNA does not affect an animal's development or survival, that mice null for this abundant and conserved RNA are morphologically and histologically normal, and that they show no sign of any perturbation in the transcriptome. Yet, we found a small but significant depletion of platelets in these animals, pointing to a potential role in hematopoiesis.
What led you to study RNA or this aspect of RNA science?
From my time as an undergrad, I have always been fascinated in understanding how gene expression and its dynamics dictate cellular decision-making in order to appropriately respond to surrounding perturbations. To begin my research journey, I joined a lab as a PhD student where I studied gene regulation using computational tools and mathematical models of gene expression. Later, I met Dr. Joana Vidigal, my current mentor, who leads an RNA biology lab where she investigates similar questions in the context of pathways that involve noncoding RNAs in vivo. I became fascinated about the ongoing research in Dr. Vidigal's lab and accepted a position in her group to begin investigating another layer of gene regulation—by noncoding RNAs—using mouse genetics.
During the course of these experiments, were there any surprising results or particular difficulties that altered your thinking and subsequent focus?
The data surprised me because I expected to see some extent of perturbations upon the loss of vaultRNA, at least at the level of RNA profiles, since several cell line-based studies have described important functions for this conserved RNA in the past. Importantly, I chose liver and brain for RNA profiles, because cell lines derived from these tissues were predominantly used to perform vaultRNA studies, but these tissues were morphologically and histologically normal and had no sign of difference at the transcriptome level upon loss of vaultRNA. This is surprising but still in line with studies where the loss of one or multiple components of the Vault particle does not produce phenotypic differences in mice. I now hypothesize that the loss of vaultRNA may only impact animal physiology when mice are challenged with stresses such as viral infection, tumorigenesis, or other cellular stresses.
What are some of the landmark moments that provoked your interest in science or your development as a scientist?
There are two major moments that provoked my interest in science: in my school days when I was first introduced to human anatomy and how different organs need to properly coordinate to make us healthy; and in my undergrad studies, when I learned that biological processes are biochemical reactions. I learned to write these down as mathematical equations, and to make models to understand how output changes upon varying input. There are two major events that developed my scientific temper: when I met Dr. Supreet Saini, who gave me the opportunity to play around with my ideas of developing mathematical models of gene expression; and when I met Dr. Joana Vidigal, who taught me mouse genetics and allowed me to explore and validate my ideas experimentally.
Are there specific individuals or groups who have influenced your philosophy or approach to science?
My PhD supervisor Dr. Supreet Saini, IIT Bombay, India, who trained me in Systems Biology and taught computational and mathematical tools, and my current mentor, Dr. Joana Vidigal, National Cancer Institute USA, who trained me in mouse genetics and taught me the techniques and tools used in wet lab.








