
Identification of Up47 in three thermophilic archaea, one mesophilic archaeon, and one hyperthermophilic bacterium
- 1Architecture et Réactivité de l'ARN, Institut de Biologie Moléculaire et Cellulaire du CNRS, Université de Strasbourg, F-67084 Strasbourg, France
- 2Laboratoire de Chimie Biologique, Université Libre de Bruxelles, Institut Labiris, Anderlecht B-1070, Belgium
- Corresponding authors: e.westhof{at}ibmc-cnrs.unistra.fr, p.wolff{at}ibmc-cnrs.unistra.fr
Abstract
Analysis of the profile of the tRNA modifications in several Archaea allowed us to observe a novel modified uridine in the V-loop of several tRNAs from two species: Pyrococcus furiosus and Sulfolobus acidocaldarius. Recently, Ohira and colleagues characterized 2′-phosphouridine (Up) at position 47 in tRNAs of thermophilic Sulfurisphaera tokodaii, as well as in several other archaea and thermophilic bacteria. From the presence of the gene arkI corresponding to the RNA kinase responsible for Up47 formation, they also concluded that Up47 should be present in tRNAs of other thermophilic Archaea. Reanalysis of our earlier data confirms that the unidentified residue in tRNAs of both P. furiosus and S. acidocaldarius is indeed 2′-phosphouridine followed by m5C48. Moreover, we find this modification in several tRNAs of other Archaea and of the hyperthermophilic bacterium Aquifex aeolicus.
Keywords
INTRODUCTION
In a previous work, we have established the profile of the tRNA modifications of several Archaea (Wolff et al. 2020). We used mass spectrometry to localize and characterize the tRNA modifications. We discovered some modifications that we did not manage to fully identify chemically. One of them was attributed to position U47 in some tRNAs from Pyrococcus furiosus (a hyperthermophilic archaeon belonging to the phylum Thermococcales) and from the thermoacidophile Sulfolobus acidocaldarius (belonging to the phylum Sulfolobales) with an assumed nucleoside mass of 338 kDa (noted as xU47). The recent identification by mass spectrometry of 2′-phosphouridine (Up47, nucleoside mass of 324 kDa) at position 47 of tRNAs of thermoacidophilic crenarcheon Sulfurisphaera tokodaii, and by extension in tRNAs from several other thermophilic Archaea (Ohira et al. 2022), prompted us to reanalyze our data.
We now find that the putative modified uridine (xU47) in P. furiosus and S. acidocaldarius corresponds indeed to a 2′-phosphouridine (Up) that is followed by 5-methylated C48 (accounting for the 14 kDa difference between the observed nucleoside masses, 338 and 324, respectively) as in S. tokodaii (Ohira et al. 2022). Moreover, we also find that Up is present at position 47 of the mesophilic archaeon Methanococcus maripaludis, in the hyperthermophilic and acidophilic archaeon Saccharolobus shibatae and interestingly also in the hyperthermophilic bacterium Aquifex aeolicus.
RESULTS AND DISCUSSION
MSMS data
The interpretation of the first MSMS data of the oligonucleotides obtained by RNase T1 of tRNAs from several archaea (the fragment of interest always started with 5′-U47C48) allowed us to conclude on the presence of xU47C48. Indeed, the expected ions corresponding to the fragmentation between U47 and C48 (fragments c1 and y5 in the example [see Fig. 1]) were missing in the spectra. Instead, we only found a fragment of 704.1 kDa corresponding to the dinucleotide xU47C48. On the other hand, RNase A analysis of the same tRNAs revealed the presence of the fragment ion carrying the same unknown modification, with a dinucleotide fragment of 722.1 kDa (corresponding to xU47C48-3′p). This allows us to conclude that the modification carried by the U47 prevents RNase A cleavage. Moreover, because C48 was cleaved by the RNase A, it suggests also that C48 was likely unmodified. This leads us to conclude that the fragment generated by RNase A corresponds to xU47C48.

Deconvoluted MS/MS sequencing spectrum (Wolff et al. 2020) (UP)(m5C)(m5C)C(Um)Gp of Ini-tRNA-CAU from S. acidocaldarius after T1 digestion (m/z = 1005.2, z = −2). No ion fragment of Up is detected, probably due to the protection conferred by 2′-phosphate and insufficient CID energy, but ions c2 (704) and y4 (1306) corresponding to the dinucleotide (Wolff et al. 2020) were detected. See Table 2 for localization of the fragment.
After publication of the first known internal post-transcriptional phosphorylation of uridine at position 47 (Ohira et al. 2022), we performed complementary experiments to determine whether this phosphoryl adduct is also present in our tRNAs studied previously (Wolff et al. 2020). To confirm the presence of Up47, the RNase T1 fragment oligonucleotides obtained from purified initiator tRNAMet from S. acidocaldarius was treated with bacterial alkaline phosphatase (BAP) followed by MS analysis. BAP can hydrolyze 5′- and 3′-phosphate but also 2′-phosphate (Ohira et al. 2022). As a matter of fact, based on the CID sequencing spectrum, the RNA fragment (UP)(m5C)(m5C)C(Um)Gp (Fig. 1), detected without BAP treatment, was converted to U(m5C)(m5C)C(Um)G (Fig. 2).
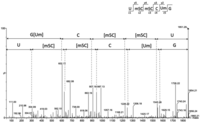
Deconvoluted MS/MS sequencing spectrum U(m5C)(m5C)C(Um)G of Ini-tRNA-CAU from S. acidocaldarius after T1 and BAP digestion (m/z = 925.13, z = −2). The spectrum clearly shows the ion fragments corresponding to U47 without phosphate (c1 = 305 and y5 = 1545). See Table 2 for localization of the fragment.
The data provided by these additional experiments allow us to conclude that the previously found xU47 was in fact Up followed by m5C. The mass of Up (324 kDa) added to the mass of the methyl group of m5C (14 Da) corresponds to the mass of the reported xU (338 kDa, see above). The missing CID fragmentation ions resulting from cleavage between U47 and C48, mentioned above (Fig. 3), could be explained by the presence of 2′-O-phosphate in U47 (Fig. 3A). We hypothesize that the presence of a 2′-phosphate group may protect the phosphodiester link against CID fragmentation, as it is the case with 2′-O-methyl (Smith and Brodbelt 2011). Hence, the fragmentation is less favorable resulting in weaker or suppressed MSMS peaks (Fig. 3B). In the case of Ohira et al. (2022), the presence of Up47 was also detected in S. acidocaldarius after digestion by nuclease P1 and LC/MS analysis (see also Table 2 in this paper).
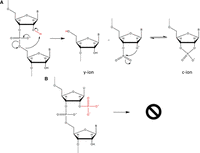
(A) Mechanism of formation of c-ions and y-ions by CID showing the involvement of 2′-OH during fragmentation (see also Tromp and Schurch 2005). (B) Scheme showing that CID formation of c-ions and y-ions is less favorable due to O2′-phosphate of Up47.
Presence of Up47 in various tRNAs
In Sulfurisphaera tokodaii, Up47 was observed in Lys-tRNA-UUU, Thr-tRNA-UGU, Gly-tRNA-UCC, Arg-tRNA-UCG, Pro-tRNA-GGG, Ile2-tRNA-CAU, and Phe-tRNA-GAA and was not observed in Gln-tRNA-UGG, Cys-tRNA-GCA, Leu-tRNA-UAG, nor Ser-tRNA-GGA (Ohira et al. 2022). Here, we find Up47 in Initiator Met-tRNA-CAU of all four archaea that we analyzed, as well as in Met-tRNA-CAU of M. maripaludis and P. furiosus (Tables 1, 2). We further find Up47 in Arg-tRNA-UCG, Asn-tRNA-GUC, Thr-tRNA-GGU, Val-tRNA-GAC, Gly-tRNA-GCC (see Supplemental Figs. S1–S4 for selected annotated spectra). As noticed (Ohira et al. 2022), all tRNAs with Up47 have 5 nt in the V-loop and Up47 is always followed by C48. Except in the archaeon M. maripaludis and bacterium A. aeolicus where it is unmodified, C48 is methylated at position 5 (m5C48). Further, in A. aeolicus, position 46 is m7G46. In five tRNAs, position 49 is also methylated at position 5, resulting in m5C49. Furthermore, in S. shibatae Initiator Met-tRNA-CAU, ac4C is detected at position 50. Expected modifications typical of Archaea are also detected (Table 2; see also Pang et al. 1982; Gupta 1984; Edmonds et al. 1991; Watanabe et al. 1997; Urbonavicius et al. 2006; Grosjean et al. 2008; Tomikawa et al. 2013; Hirata et al. 2019; Yu et al. 2019; Rose et al. 2020; Sas-Chen et al. 2020; Wolff et al. 2020; Ohira et al. 2022).
List of tRNAs containing Up47 identified by LC-MSMS with relevant LC/MS data and sequences
Compilation of modified tRNA sequences from M. maripaludis, P. furiosus, S. acidocaldarius, S. shibatae, and A. aeolicus
Conclusions
To date the presence of post-transcriptional internal phosphorylation of a nucleotide in tRNA as Up47 was reported in hyperthermophilic Archaea and in Nitrososphaera viennensis, a mesophilic archaeon (Ohira et al. 2022). Here, we show that this modification is also present in the mesophilic archaeon M. maripaludis. Most interestingly we find this modification in A. aeolicus, a hyperthermophilic bacterium. The presence of Up47 in several thermophilic archaea and A. aeolicus was only suspected from the presence in the genomes of gene arkI homologs coding for specific U47 kinase. The recombinant form of this newly identified modification enzyme from T. kodakarensis can catalyze base phosphorylation of U47 in vitro in the presence of ATP (Ohira et al. 2022). In Table 1, potential arkI homologs, annotated as Ser/Thr kinases, in the various archaeal species studied are indicated (see also Ohira et al. 2022). In A. aeolicus, the gene (aq_578) is in a region of the chromosome containing tRNA genes with, as direct neighbor, the polypeptide deformylase gene. In sum, these results show that this internal nucleotide phosphorylation is a modification not only present in thermophilic but also in mesophilic archaea (as in the mesophilic bacterium Nautilia profundicola where ArkI has Up-modifying activity in vitro, Ohira et al. 2022) and in hyperthermophilic Bacteria. The present note completes our previous report on tRNA modifications in Archaea (Wolff et al. 2020).
MATERIALS AND METHODS
tRNA isolation by 2D-gel electrophoresis
tRNA isoacceptors were isolated using two-dimensional gel electrophoresis as previously described (Wolff et al. 2020). Briefly, the total tRNA of each organism was briefly heated at 90°C. Then, samples were separated in a first-dimension gel under denaturing conditions using 12.5% polyacrylamide gel, 1× TBE and 8 M urea, followed by a second dimension under semidenaturing conditions using 20% polyacrylamide gel, TBE 1× and 4 M urea at room temperature. Gels were stained with an ethidium bromide solution (10 µg/L−1) for about 10 min. Finally, spots containing tRNAs were visualized and excised under UV light (302 nm).
In-gel RNase digestion of tRNAs
Gel spots containing tRNAs were first desalted by several washes (at least eight) with 100 µL 200 mM NH4AcO and gel spots were dried under vacuum. For RNase T1 and RNase A hydrolyzes, gel pieces were rehydrated by 20 µL of 0.1 U/µL−1 of RNase T1 or by 20 µL of 0.01 U/µL−1 of RNase A (Thermo Fisher Scientific) in 100 mM NH4AcO (pH is not adjusted). For RNase U2 digestion, spots were digested by using 50 µL of RNase U2 (homemade) at 0.3 ng/µL−1 in 50 mM NH4AcO (pH 5.3) and incubated for 45 min at 65°C. After digestion, supernatants were dried under vacuum. Bacterial alkaline phosphatase (BAP) dephosphorylation was performed by adding 10U of BAP to the RNase mixture and incubating for 3 h at 37°C.
NanoLC-MS/MS of RNA oligonucleotides
Pellets containing RNase digestion products were resuspended in 3 µL of milli-Q water and separated on an Acquity peptide BEH C18 column (130 Å, 1.7 µm, 75 µm × 200 mm) using a nanoAcquity system (Waters). The analyses were performed with an injection volume of 3 µL. The column was equilibrated in buffer A containing 7.5 mM TEAA (Triethylammonium acetate), 7.0 mM TEA (Triethylammonium), and 200 mM HFIP (Hexafluoroisopropanol) at a flow rate of 300 nL/min−1. Oligonucleotides were eluted using a gradient from 15% to 35% of buffer B (100% methanol) for 2 min followed by an increase of buffer B to 50% in 20 min. MS and MS/MS analyses were performed using SYNAPT G2-S (hybrid quadrupole time-of-flight mass spectrometer) from Waters Corporation. All experiments were performed in negative mode with a capillary voltage set at 2.6 kV and a sample cone voltage set at 30 V. Source was heated to 130°C. The samples were analyzed over an m/z range from 550 to 1600 for the full scan, followed by fast data direct acquisition scan (Fast DDA). Collision-induced dissociation (CID) experiments were achieved using Ar.
Data analysis
All MSMS spectra were deconvoluted using MassLynx software from Waters and manually sequenced by following the y and/or c series (w ions were also useful when sequencing was difficult or to confirm a sequence). tRNA identification was done by comparisons with the genomic sequences obtained from GtRNAdb (http://gtrnadb.ucsc.edu/) (Chan and Lowe 2016). Data about nucleoside modification were obtained from Modomics database (https://iimcb.genesilico.pl/modomics/modifications) (Boccaletto et al. 2022).
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
The project was supported by the French National Program Investissement d'Avenir (Labex NetRNA) administered by the Agence Nationale de la Recherche (ANR-10-LABX-0036_NETRNA). This work of the Interdisciplinary Thematic Institute IMCBio, as part of the ITI 2021-2028 program of the University of Strasbourg, CNRS and Inserm, was supported by IdEx Unistra (ANR-10-IDEX-0002), and by SFRI-STRAT'US project (ANR 20-SFRI-0012) and EUR IMCBio (ANR-17-EURE-0023) under the framework of the French Investments for the Future Program. Anthony Lechner was supported by Fonds Régional de Coopération pour la Recherche (Region Grand Est, EpiRNA).
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079546.122.
-
Freely available online through the RNA Open Access option.
- Received December 6, 2022.
- Accepted January 19, 2023.
This article, published in RNA, is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








