
Methylation guide RNAs without box C/D motifs
- Jiayin Wang1,2,
- Zuxiao Yang1,3 and
- Keqiong Ye1,2
- 1Key Laboratory of RNA Biology, CAS Center for Excellence in Biomacromolecules, Institute of Biophysics, Chinese Academy of Sciences, Beijing 100101, China
- 2University of Chinese Academy of Sciences, Beijing 100049, China
- 3Institute of Chinese Integrative Medicine, Hebei Medical University, Shijiazhuang 050017, Hebei, China
- Corresponding author: yekeqiong{at}ibp.ac.cn
Abstract
Box C/D RNAs guide site-specific 2′-O-methylation of RNAs in archaea and eukaryotes. The defining feature of methylation guide RNAs is two sets of box C and D motifs that form kink-turn structures specifically recognized by L7Ae family proteins. Here, we engineered a new type of methylation guide that lacks C/D motifs and requires no L7Ae for assembly and function. We determined a crystal structure of a bipartite C/D-free guide RNA in complex with Nop5, fibrillarin and substrate in the active form at 2.2 Å resolution. The stems of new guide RNAs functionally replace C/D motifs in Nop5 binding, precisely placing the substrate for site-specific modification. We also found that the bipartite architecture and association of L7Ae with C/D motifs enhance modification when association of guide RNAs or substrates is weak. Our study provides insights into the variations, robustness and possible evolutionary path of methylation guide RNAs.
Keywords
INTRODUCTION
Guide RNAs recognize nucleic acid substrates by sequence complementarity and instruct protein enzymes or effectors to act on the substrates. Major classes of guide RNA include siRNA, miRNA, crRNA, box C/D RNA, and box H/ACA RNA. C/D RNA and H/ACA RNA are among the first discovered guide RNAs and direct 2′-O-methylation and pseudouridylation of RNAs, respectively (Kiss 2001; Watkins and Bohnsack 2011; Yu and Meier 2014). They are ubiquitously present in eukaryotes and archaea, but absent in bacteria. In eukaryotes, they are located in the nucleolus and Cajal body, known as small nucleolar and small Cajal body RNAs (snoRNAs and scaRNAs), and primarily guide modification of rRNAs and snRNAs, respectively. In archaea, their substrates include rRNAs and tRNAs.
C/D RNAs are named after the conserved motifs box C (RUGAUGA, R = purine) and box D (CUGA) located at the 5′ and 3′ end, respectively. C/D RNAs additionally contain related box C′ and D′ motifs in the internal region, which are connected to box C and D motifs by two spacers. The terminal C/D motifs and nearby sequences combine into a kink-turn (K-turn) structure, while the internal C′/D′ motifs often form a K-loop structure (Vidovic et al. 2000; Klein et al. 2001; Nolivos et al. 2005; Huang and Lilley 2018) (As the C/D and C′/D′ motifs are functionally equivalent in vitro, they are indistinguishably referred to as C/D motifs when their position and structural difference are irrelevant in description. Similarly, K-turn and K-loop are sometimes called K-turn.) One or both spacers can pair with substrate RNAs and select the nucleotide paired to the fifth position upstream of box D′ or D for modification (Cavaille et al. 1996; Kiss-Laszlo et al. 1996; Tycowski et al. 1996). Consequently, C/D RNAs own a bipartite structure with two sets of C/D motifs and spacers.
Archaeal C/D RNAs have a compact size (60–70 nt), ∼12 nt spacers and highly conserved box C′/D′ motifs (Omer et al. 2000). To form an enzymatically active RNA–protein complex (RNP), an archaeal C/D RNA is associated with two copies each of the scaffolding protein Nop5, the S-adenosyl methionine (SAM)-dependent methyltransferase fibrillarin and the RNA-binding protein L7Ae (Omer et al. 2002; Tran et al. 2003; Lin et al. 2011). Nop5 homodimerizes through its coiled-coil domain and recruits fibrillarin by its mobile amino-terminal domain (NTD) (Aittaleb et al. 2003). The K-turn structure formed by C/D motifs is specifically recognized by L7Ae and the binary complex is then associated to the carboxy-terminal domain (CTD) of Nop5, anchoring the guide RNA onto the enzyme (Moore et al. 2004; Liu et al. 2007; Ye et al. 2009; Lin et al. 2011). Eukaryotic C/D RNAs have variable sizes (60–130 nt), long spacers and frequently imperfect C′/D′ motifs. They associate with Nop56, Nop58, fibrillarin, and Snu13 (an L7Ae homolog) (Yang et al. 2020). Nop58 and Nop56 are paralogs of Nop5, form a heterodimer and bind the C/D and C′/D′ motifs, respectively (Cahill et al. 2002; Granneman et al. 2009).
The C/D motifs, not the C′/D′ motifs, are essential for processing and accumulation of eukaryotic C/D snoRNAs (Jarmolowski et al. 1990; Cavaille and Bachellerie 1996; Cavaille et al. 1996; Watkins et al. 1996; Kiss-Laszlo et al. 1998). But at the structural level, the C/D and C′/D′ motifs are functionally independent and specifically required to the function of coupled spacer upstream of box D/D′ (Tran et al. 2003; Qu et al. 2011; Yang et al. 2020).
In this study, we surprisingly found with biochemical and structural analysis that the C/D motifs in methylation guide RNAs can be functionally replaced by a simple duplex in archaeal enzymes. We created a novel type of methylation guide RNAs that lack any C/D motifs and do not depend on L7Ae for assembly and function. By studying various mutants, we gained new understanding about the contribution of bipartite architecture and association of L7Ae with C/D motifs to the robustness of methylation guide RNAs.
RESULTS
Methylation guide RNAs without C/D motifs
sR1c is a typical archaeal C/D RNA and has been subjected to extensive mutational analysis (Fig. 1A; Yang et al. 2020). We created a mutant of sR1c, termed bipartite methylation guide 1 (BMG1), by deleting its terminal C/D motifs while keeping a terminal stem of 4 bp. To measure RNA-guided methylation activities, guide RNAs were assembled with the archaeal Sulfolobus solfartaricus (Ss) proteins Nop5, Fib and L7Ae or the eukaryotic Chaetomium thermophilum (Ct) proteins Nop56, Nop58, Fib, and Snu13 and incubated with a cognate substrate paired to the D′ or D spacer (Fig. 1B; Ye et al. 2009; Lin et al. 2011; Yang et al. 2020). Substrates premethylated at target sites were also assayed to evaluate the site specificity of modification. As shown previously, sR1c guided the methylation of both the D′ and D substrates when loaded in both Ss RNPs and Ct RNPs (Yang et al. 2020). Because archaeal sR1c does not fit well into the Ct RNP, Ct RNP-mediated modification was inefficient for the D′ substrate and enhanced when the C/D motifs were removed (Yang et al. 2020). BMG1 efficiently guided the site-specific modification of the D′ substrate in Ss RNPs and Ct RNPs (Fig. 1B, lane 4). This was expected since the C/D and C′/D′ motifs are functional independent (Yang et al. 2020) and BMG1 still contains intact C′/D′ motifs. Surprisingly, the D substrate was also efficiently modified by the BMG1-loaded Ss RNP (Fig. 1B, lane 5). The activity was specific to archaeal Ss RNPs since the D substrate was not methylated by eukaryotic Ct RNPs (Fig. 1B, lane 5). As the results were apparently contradictory to the consensus that C/D motifs are essential for guiding function, we further analyzed the mechanism underlying the unusual activity of BMG1.
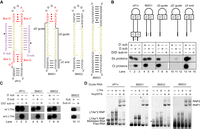
Activity and assembly of bipartite methylation guide RNAs without C/D motifs. (A) Secondary structure models of sR1c, BMG1, BMG2, and BMG3. Box C, D, C′, and D′ motifs are colored in red and spacers in yellow. The D′ and D substrates (sub) are shown in purple with modification sites marked by circles. The sequences deleted in BMG1 mutants are indicated with lines. (B) Methylation assay. Ss RNPs were assembled with guide RNAs (1 µM), Nop5/Fib complex (2 µM), and L7Ae (3 µM). Ct RNPs were assembled with guide RNAs (2 µM), Nop56/Nop58/Fib complex (∼2 µM), and Snu13 (6 µM). The assembled RNPs were incubated with the D or D′ substrates (30 µM) or mixed premethylated substrates (sub-m) in the presence of 30 µM cold SAM and trace amounts of [methyl-3H] SAM for 20 min at 70°C for Ss RNPs or at 50°C for Ct RNPs. RNAs were resolved in denaturing PAGE and visualized by 3H autoradiography. (C) Activity of Ss RNPs in the presence and absence of L7Ae. (D) EMSA of Ss RNPs. Each RNA (1 µM) was assembled with L7Ae (1 or 2 µM, indicated by a small or large plus sign) and Nop5/Fib complex (2 µM) in various combinations, resolved in 5% native PAGE and strained by SYBR Gold. L7Ae*1 and L7Ae*2 RNPs contain one and two copies of L7Ae. RNP1 and RNP2 refer to monomeric and dimeric species. Misfolded RNAs that migrated slower and bound poorly to L7Ae are marked with asterisk.
Deletion of the D spacer of BMG1 eliminated modification of the D substrate (Fig. 1B, lane 8), confirming that the activity was RNA-guided. Modification on the D substrate was greatly reduced when the terminal stem was disrupted by removing 18 nt at the 5′ end or 4 nt at the 3′ end of BMG1 (Fig. 1B, lanes 11 and 14). These suggest that the terminal stem is critical for the function of the D spacer and may replace the structural role of C/D motifs. In light of this idea, we further replaced the internal box C′/D′ motifs of BMG1 with a 4-bp stem, yielding a BMG2 RNA without any C/D motifs (Fig. 1A). Indeed, BMG2 guided site-specific modification of both the D and D′ substrates (Fig. 1C, lanes 7–9). For structural study, we designed a two-pieced, more symmetric C/D-free guide RNA, BMG3, which was composed of two 8-bp stems linked by two identical 13-nt sequences (Fig. 1A). BMG3 also turned out to be a functional methylation guide (Fig. 1C, lanes 10–11), indicating that the terminal stem can be longer than 4-bp.
Since L7Ae specifically binds the K-turn structure formed by C/D motifs (Vidovic et al. 2000; Moore et al. 2004), C/D-free methylation guide RNAs in principle do not require L7Ae for assembly and activity. Indeed, for BMG1, the activity of the D spacer coupled to the terminal stem did not depend on L7Ae, whereas the D′ spacer coupled to the internal C′/D′ motif still required L7Ae (Fig. 1C, lanes 4,5). BMG2 and BMG3 completely lacked C/D motifs and were able to guide specific modification of all substrates in the absence of L7Ae (Fig. 1C, lanes 7–11). sR1c depended on L7Ae for guiding modification of the D′ substrate (Fig. 1C, lane 1), but puzzlingly, it still guided modification of the D substrate in the absence of L7Ae (Fig. 1C, lane 2). Further studies on the unusual activity will be described later.
The assembly of Ss RNPs was examined with electrophoretic mobility shift assay (EMSA) (Fig. 1D). As expected for specific association of L7Ae with K-turn, sR1c with two sets of C/D motifs bound one or two copies of L7Ae, BMG1 with one set of C/D motifs bound only one copy of L7Ae, and BMG2 and BMG3 that contain no C/D motifs only associated nonspecifically with L7Ae. All these guide RNAs can assemble with the Nop5/Fib complex alone, accounting for their activity in the absence of L7Ae. Dimeric RNPs (RNP2) were observed for all RNAs due to self-association of monomeric RNPs (RNP1) (Yang et al. 2020). In conclusion, we have generated a new type of methylation guide that differs from natural C/D RNAs by lacking C/D motifs and requiring no L7Ae for assembly and function.
Structure of a C/D-free RNA-guided methylation RNP
To understand the structural basis of C/D-free methylation guide RNA activities, we determined a crystal structure of BMG3 RNA in complex with Nop5, Fib and substrate (Fig. 2A,B). The structure was solved by molecular replacement with the substrate-bound SL12 (also called CD45) C/D RNP structure minus L7Ae as a searching model (Lin et al. 2011) and refined to 2.2 Å resolution (Supplemental Table S1; Supplemental Fig. S1A). Despite the lack of L7Ae and C/D motifs, the BMG3 RNP structure closely resembles the SL12 RNP structure with a root mean square deviation (RMSD) of 0.709 Å over 1114 Cα atom pairs (Fig. 2C,D; Supplemental Fig. S1B). In the BMG3 RNP structure, Nop5 forms a homodimer by its coiled-coil domain, associates fibrillarin with its NTD and binds the terminal stems of BMG3 with its CTD. Each substrate forms a 10-bp duplex with the spacer of BMG3 and contacts fibrillarin. The active site of fibrillarin is loaded with a copurified S-adenosyl homocysteine (SAH) molecule, which differs from SAM by lacking the methyl donor group and an unmodified ribose at the target site (Supplemental Fig. S1A). These features indicate that the structure represents an active conformation prior to modification.
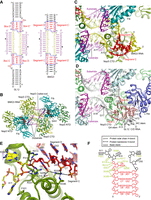
Crystal structure of substrate-bound BMG3 RNP. (A) Secondary structure diagrams of SL12 and BMG3 RNAs. C/D motifs in SL12 and segments C and D in BMG3 are colored in red, spacers in yellow and substrates in purple. Modification sites are marked by circles. The structurally equivalent Nop5-binding regions are encircled with blue boxes. Nucleotides in guide RNAs and substrates are separately numbered. Spacer sequences are additionally labeled by yellow numbers to indicate the distance to box D/D′ or segment D/D′. (B) Ribbon representation of structure of BMG3 in complex with Nop5, fibrillarin (Fib) and substrate. Two Nop5 molecules are colored in forest green and split pea, Fib in teal, guide RNA in yellow and substrates in purple. SAH bound in the active site of Fib is shown as red spheres. (C,D) RNA–protein interfaces in the BMG3 RNP (C) and SL12 RNP (PDB code: 3PLA) (D) structures. (E) Detailed interactions between BMG3 RNA and Nop5 at segments C/D. Hydrogen bonds are denoted by dashed lines. (F) Schematic of interactions between BMG3 RNA and Nop5.
In the conventional C/D RNP structure (Lin et al. 2011), the C/D RNA is assembled to the complex through two K-turns formed by the C/D motifs and their nearby sequences (Fig. 2A,D). A typical K-turn consists of a noncanonical stem containing tandem sheared GA pairs (GA stem), a 3-nt loop and a canonical stem composed of Watson–Crick pairs (WC stem) (Vidovic et al. 2000; Klein et al. 2001). The WC stem is minorly contacted by Nop5 and L7Ae and could be absent or replaced by a loop to form a K-loop. The K-turn structure is bound by L7Ae and the CTD of Nop5, two proteins that also interact with each other in C/D RNP structures. L7Ae specifically recognizes the GA stem and the internal loop, including a conserved projecting uridine. The CTD of Nop5 binds the GA stem, several adjacent spacer residues of C/D RNA and the loaded substrate. The α9′ helix of Nop5 is a key RNA-binding element that is located at the three-way junction of two guide-substrate duplexes and the GA-stem.
The terminal stem of BMG3 RNA lacks a K-turn and hence cannot bind L7Ae. Nevertheless, the terminal stem still contacts the CTD of Nop5 in a similar manner as the GA stem of C/D RNA does (Fig. 2C,D; Lin et al. 2011). Specifically, the proximal 4-bp duplex of the terminal stem is structurally equivalent to the GA stem of C/D RNA, which is composed of nucleotides 3–6 of box C (RUGAUGA) and the entire box D (CUGA). In addition, the unpaired base (G9 and G38 of BMG3) located at the proximal 3′ end of the terminal stem is structurally equivalent to the seventh base of box C (A16 and A56 of SL12). Both bases stack over the GA stem and bind Nop5. To show structural analogy with C/D RNA, we termed a 5-nt segment C or C′ in the C/D-free guide RNA that corresponds to nucleotides 3–7 of box C or C′ and a 4-nt segment D or D′ that corresponds to box D or D′ (Fig. 2A,C,D). Consequently, BMG3 contains two spacers with an effective length of 12 nt, just like the SL12 C/D RNA. At the side of segments C/D, Nop5 primarily binds the 3′ half of segment C and several spacer residues and barely contacts segment D (Fig. 2E,F), which is more distant to Nop5 compared to box D of C/D RNA (Supplemental Fig. S1B,C). Basic residues R313, K316, R331, K336, and R339 interact with the phosphate groups in segment C and spacers. In addition, residues Q296 and R339 make two hydrogen bonds with the O6 atom of unpaired G9. The protein–RNA interactions are similar at the side of segments C′/D′, but several interactions vary due to the different positions of protein side chains (Supplemental Fig. S1D).
In the BMG3 RNP structure, nucleotides 1 and 12 of the spacers (A10/U21 or A39/U50) do not bind substrates but contact helix α9′ of Nop5. Nucleotides 2–11 (A11–G20 or A40–G49) of the spacer pair with the substrate to form a 10-bp duplex. Lying over the dimerized coiled-coil domain of Nop5, the guide-substrate duplex is capped by two opposing CTDs of Nop5 at both ends and limited in size to 10 bp at the active state (Yang et al. 2016). Although U11/U31 of the substrate was designed to pair with nucleotide 12 of the spacer, it is unpaired in the structure and stacks over H327, as observed previously in a C/D RNP structure (Yang et al. 2016). The conformation of the guide-substrate duplex and its interactions with fibrillarin and Nop5 are nearly identical in both the structures of SL12 C/D RNP and BMG3 C/D-free RNP. Hence, the terminal stem of BMG3 can functionally replace the GA stem of C/D RNA in binding Nop5, ensuring the correct placement of the guide-substrate duplex and the site-specific modification.
Monopartite methylation guide RNAs without C/D motifs
The bipartite architecture has been shown to be dispensable for C/D RNA, and halfmer C/D RNAs with a single set of C/D motifs and a guide are also functional in vitro (Yang et al. 2020). We wondered if C/D-free guide RNAs can be also reduced to a monopartite form. We initially designed a monopartite methylation guide RNA (MMG1) containing a stem–loop and a 12-nt guide at the 5′ end, which has identical sequences as the D spacer of sR1c and BMG2 (Fig. 3A). The modification site was predicted to pair with position 5 of the guide as in bipartite methylation guides.
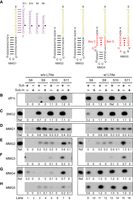
Activity of monopartite methylation guide RNAs. (A) Secondary structure diagrams of MMG1 to MMG5. These RNAs share the same guide sequence as the D guide of sR1c and BMG2. The pairing of MMG1 with substrates S8–S10 are shown. Modification targets are indicated by circles. (B–H) Activity and specificity of Ss RNPs assembled with sR1c (B), BMG2 (C), MMG1 (D), MMG2 (E), MMG3 (F), MMG4 (G), and MMG5 (H) and without (w/o) or with (w/) L7Ae. Substrates S8–S10 or their premethylated versions (sub-m) were modified at 70°C for 20 min. Reactions from a same gel are boxed. Relative modification levels were quantified against the reference reaction (Ref) present in each gel that showed modification of S10 by sR1c holo-RNP.
Four substrates S8–S11 that formed 8–11 bp duplexes with the guide were tested for modification by Ss RNPs assembled with or without L7Ae. Shorter substrates were commonly modified to less degrees and sometimes with less specificity. All substrates were efficiently modified at the target site by the fully assembled sR1c RNP (Fig. 3B, lanes 9–16). All substrates except for S8 were efficiently and specifically modified by the bipartite C/D-free BMG2 RNP in the L7Ae-independent manner (Fig. 3C). The monopartite MMG1 RNA poorly guided the methylation of these substrates (Fig. 3D). Moreover, the substrates premethylated at predicted target sites were modified to similar degrees as the unmethylated substrates, indicating that the modification occurred mainly at other sites.
In the BMG3 RNP structure, three nucleotides (G9, A10 and A11) in the 3′ overhang of the terminal stem interact extensively with the CTD of Nop5 (Fig. 2E). The three nucleotides contribute to anchoring the guide RNA onto Nop5, but are missing in MMG1. The stem of MMG1 might unwind on the top to create a 3′-overhang to bind Nop5, which would shift the stem's position on Nop5 and alter the specificity of modification. To fix this problem, the 3′ end of MMG1 was extended by AGA, generating an MMG2 RNA (Fig. 3A). In MMG2, the first adenosine of the 3′ overhang could potentially pair with a uridine at position 1 of the guide. To avoid this pairing, the uridine of the guide was further replaced with a cytidine to create an MMG3 RNA (Fig. 3A).
We found that MMG3 can guide modification of substrates S10 and S11 at the expected target but with poor activities (Fig. 3F). The targeting specificity of MMG2 fell between that of MMG1 and MMG3, suggesting that the terminal AU pair was partially formed in MMG2. All the tested monopartite C/D-free guides were much less active than bipartite C/D-free guides. Finally, L7Ae did not affect both the activity and specificity of all C/D-free methylation guides, as expected (Fig. 3C–F).
We also assessed the activity of monopartite C/D RNAs that contain a K-turn (MMG4) or a K-loop (MMG5) (Fig. 3A). In the absence of L7Ae, both RNAs displayed poor activities (Fig. 3G–H, lanes 1–8). MMG4 showed considerable specificity for substrates S10 and S11, but poor specificity for shorter substrates. MMG5 has worse guiding specificity than MMG4. When assembled with L7Ae into holoenzymes, both RNAs guided modification of all substrates with strict specificity and high efficiency (Fig. 3G,H, lanes 9–16).
In sum, monopartite C/D-free guide RNAs require a 3′-overhang for proper assembly and display rather poor activities. Their activity and specificity can be greatly improved by becoming bipartite (BMG2) or incorporating C/D motifs and L7Ae (MMG4 and MMG5).
L7Ae is dispensable for modification of strongly paired substrates in C/D RNP
L7Ae recognizes the K-turn and is considered to be essential for the assembly and function of C/D RNA. The activity assay of sR1c showed that the lack of L7Ae blocked modification of the D′ substrate, but did not affect modification of the D substrate (Fig. 1C, lanes 1–3 and Fig. 4B, lanes 1–3). The D′ and D guides were coupled to the internal C′/D′ motif and the terminal C/D motif, respectively (Fig. 4A). To test whether their positions mattered, the two guide sequences were exchanged in sR1c-ex RNA. The exchanged guide regions both worked in the fully assembled RNPs, but instructed modification of only the original D substrate in the absence of L7Ae (Fig. 4B, lanes 4–6). Hence, the position of these guides is not related to their different activities.
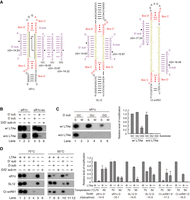
L7Ae is dispensable for modification of strongly associated substrates by C/D RNPs. (A) Substrate interactions of sR1c, SL12, and Ct snR61 RNAs. Gibbs free energy (ΔG) is indicated for each guide-substrate duplex. The D substrate of sR1c was mutated to change the GC pair at position 3 of the guide-substrate duplex to a GU or GG pair. (B) Activity of sR1c and sR1c-ex where the D and D′ guides were exchanged. The D or D′ substrates and their premethylated versions were modified at 70°C by Ss C/D RNPs assembled with (w/) or without (w/o) L7Ae. Lanes 1–3 are copied from Figure 1C. (C) The D substrate mutants (S) and their premethylated versions (M) were modified at 70°C by sR1c RNPs assembled with (w/) or without (w/o) L7Ae. The relative modification levels were normalized to modification of wild-type (GC) substrate by sR1c holo-RNP. Averages and standard deviations were calculated from three replications. (D) Modification at 70°C and 50°C. sR1c, SL12, and Ct snR61 RNPs were assembled with or without L7Ae and assayed on their cognate D and D′ substrates and mixed premethylated substrates. The relative modification levels were normalized to the reactions catalyzed by holo-RNPs at 70°C. Averages and standard deviations were calculated from two replications.
We noticed that sR1c formed a more stable duplex with the D substrate than the D′ substrate (ΔG = −18.05 vs −14.83 kcal/mol) and hypothesized that L7Ae may be dispensable for strongly paired substrates. To test this idea, the stability of the D guide-substrate duplex was reduced by replacing a GC pair at position 3 with a GU wobble pair (ΔG = −15.87 kcal/mol) or a GG mismatch (ΔG = −14.22 kcal/mol). The GU pair did not affect much modification by the holo-enzyme, but abolished modification by the L7Ae-lacking enzyme (Fig. 4C, lane 3). The GG mismatch greatly reduced modification by the holo-enzyme and totally abolished modification by the L7Ae-lacking enzyme (Fig. 4C, lane 5). Along the same line, short substrates S8 and S9, but not long substrates S10 and S11, showed L7Ae dependence for modification (Fig. 3B). We conclude that L7Ae could be dispensable for modification of strongly bound substrates.
To stabilize pairing interactions between substrates and guide RNAs, modification reactions were conducted at a lower temperature of 50°C (C/D RNPs from thermophilic Ss were commonly assayed at 70°C). Interestingly, the D′ substrate of sR1c was now modified to similar degrees in the presence and absence of L7Ae (Fig. 4D, row 1, lanes 7 vs 10). This indicates that L7Ae became dispensable when the substrate binding was strengthened at lower temperatures. Modification of the D substrate remained independent of L7Ae at 50°C (Fig. 4D, row 1, lanes 8 vs 11), yet was much worse than modification of the D′ substrate. Product release was probably inhibited for stably paired substrates at lower temperatures in the multiple-turnover reactions. We further analyzed how reaction temperature affects the activity of two other C/D RNAs: SL12 and Ct snR61 (Fig. 4D; Lin et al. 2011; Yang et al. 2020). The D′ substrate of SL12 and the D′ substrate of Ct snR61 cannot be modified by L7Ae-free RNPs at 70°C, but can at 50°C (Fig. 4D, rows 2 and 3, lanes 4 vs 10). These data show that low reaction temperature can enhance stability of guide-substrate duplexes and alleviate the requirement of L7Ae in C/D RNA-guided modification.
DISCUSSION
Natural methylation guide C/D RNAs are characterized by two sets of C/D motifs that are specifically recognized by L7Ae. We have engineered a new type of methylation guide RNAs that contain no C/D motifs and obviate L7Ae for assembly and function. The C/D-free methylation guide RNAs consist of two stems connected with single-stranded spacers which can bind substrates and select a target at position 5 upstream of the stem for modification. These RNAs assemble with archaeal Nop5 and fibrillarin into active enzymes that can modify substrates as efficiently and specifically as conventional C/D RNPs. Our structural analysis of the BMG3 RNP shows that the stem functionally replaces the C/D motifs in binding Nop5, placing the guide-substrate duplex in the same manner as in C/D RNP structures. The C/D-free RNAs are not active in eukaryotic Ct complexes, suggesting that Nop56 and Nop58 cannot accommodate the duplex substitution of C/D motifs.
We also created monopartite C/D-free guide RNAs that comprise a stem, a 3-nt overhang at the 3′ end and a guide at the 5′ end. The 3′ overhang is needed for registering the guide RNA onto Nop5 and ensuring modification specificity. However, these monopartite RNAs are poor guides and not practically useful. Bipartite structures are clearly advantageous over monopartite structures for C/D-free methylation guide RNAs. The advantage of being bipartite is unclear for conventional C/D RNAs, since halfmer C/D RNAs displayed similar activity and specificity as bipartite C/D RNAs in holo-RNPs (compare sR1c vs MMG4 and MMG5, Fig. 3B,G,H). Nevertheless, halfmer C/D RNAs were much worse than bipartite C/D RNAs in L7Ae-lacking RNPs. The gain of bipartite structure may be masked by the strong stabilizing effect of L7Ae association of C/D motifs in conventional C/D RNAs and surfaces only when the association of guide RNA is weakened in the absence of C/D motifs (Fig. 3) or L7Ae (Fig. 4). Hence, being bipartite makes C/D RNA a more robust guide.
We have found an interesting interplay between the binding strength of substrates and the assembly of C/D RNAs. When the assembly of C/D RNA is impaired in the absence of L7Ae, weakly bound substrates cannot be modified, but strongly bound substrates can still be modified (Fig. 4). This suggests that association of L7Ae with C/D motifs enhances the binding of substrates and is particularly important for modification of weakly associated substrates.
Our analyses on functional variants of C/D RNA provide new insights into its structural features. Although bipartite architecture, C/D motifs and L7Ae binding are not individually essential, they contribute to the robustness of methylation guide RNAs and become important when association of guide RNA is compromised or substrate binding is weak.
Our study demonstrates that methylation guide RNAs could be assembled in vitro into archaeal Ss RNPs via simple duplexes in the absence of L7Ae. Whether this kind of C/D-free guide RNA is present in archaea remains an open question. It is possible that some archaeal C/D RNAs might have only one of its C/D motifs replaced by a stem (like BMG1). The C/D-free guides with a simpler structure and less bound proteins might represent a primordial form of methylation guide.
MATERIALS AND METHODS
Methylation assay
Protein purification, RNA preparation, RNP assembly and activity measurement were conducted as previously described (Ye et al. 2009; Lin et al. 2011; Yang et al. 2020). Ss RNPs were assembled from guide RNAs (1 µM), Ss Nop5/Fib complex (∼2 µM), and L7Ae (3 µM). Ct RNPs were assembled from guide RNAs (2 µM), a copurified Nop56/Nop58/Fib complex (∼2 µM), and Snu13 (6 µM). The assembled RNPs were mixed with substrates (30 µM) complementary to the D or D′ spacers or mixed premethylated substrates in the presence of 30 µM cold SAM and trace amounts of [methyl-3H] SAM. The reactions were incubated for 20 min at 70°C for Ss RNPs or 50°C for Ct RNPs. RNAs were resolved in denaturing polyacrylamide gel electrophoresis (PAGE) and visualized by 3H autoradiography. The gels were exposed to X-ray films for 5–6 d.
Electrophoretic mobility shift assay
For EMSA,10 pmol of guide RNA was assembled with 0, 10 or 20 pmol of L7Ae, or 20 pmol of Nop5/Fib, or 20 pmol of L7Ae and 20 pmol of Nop5/Fib in 10 µL of binding buffer containing 25 mM HEPES-K pH 7.6, 300 mM NaCl, 0.01% NP-40, 10% glycerol, and 2 mM MgCl2. The RNPs were mixed with 1 µL of native loading dye containing 50% glycerol, 0.25% (w/v) bromophenol blue and 0.25% (w/v) xylene cyanol FF and separated by 5% native PAGE for 40 min running in Tris-glycine (pH 8.3) buffer with a power of 5 W. RNAs were stained with SYBR GOLD (Invitrogen) and visualized with a GelDoc XR+ imaging system (Bio-Rad).
Crystallization and structure determination
The BMG3 RNP (∼10 mg/mL) was prepared in 5 mM HEPES-Na (pH 7.6) buffer as described previously (Lin et al. 2011; Yang et al. 2016). The substrate RNA was added twice as much as the RNP before crystallization. Preliminary crystallization screen was performed with commercial kits using the hanging-drop vapor diffusion method at 18°C. Crystals were grown from 0.2 M sodium citrate tribasic dehydrate and 20% (w/v) polyethylene glycol 3350 (pH 8.3). Crystals were cryo-protected in 10% glycerol made in the well solution and flashed frozen in liquid nitrogen.
Diffraction data were collected in the Shanghai Synchrotron Research Facility (SSRF) BL17U1 beamline (Wang et al. 2018) and processed with the HKL-2000 program (Otwinowski and Minor 1997). The structure was determined by molecular replacement with the structure of SL12 RNP (PDB code: 3PLA) with L7Ae removed as a search model. The model was built in COOT (Emsley et al. 2010) and refined in PHENIX (Adams et al. 2010). Structural figures were prepared with PyMOL (Schrödinger).
DATA DEPOSITION
The atomic coordinates and structure factors have been deposited in the Protein Data Bank with access code 7XPL (PDB DOI: 10.2210/pdb7xpl/pdb).
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
We thank the staff at the SSRF for assistance in data collection, Hongjie Zhang for help in radioactivity experiments and Xiuling Gao for technical assistance. The study was supported by National Natural Science Foundation of China (91940302, 91540201, 31430024, 31325007), National Key R&D Program of China (2017YFA0504600), and Strategic Priority Research Program of Chinese Academy of Sciences (XDB37010201).
Author contributions: K.Y. conceived and supervised the work. J.W. conducted biochemical and structural studies. Z.Y. contributed to mutational analysis. J.W. and K.Y. prepared the manuscript.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079379.122.
- Received July 24, 2022.
- Accepted September 13, 2022.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








