
Evolutionary specialization of recoding: Frameshifting in the expression of S. cerevisiae antizyme mRNA is via an atypical antizyme shift site but is still +1
Abstract
An autoregulatory translational shift to the +1 frame is required for the expression of ornithine decarboxylase antizyme from fungi to mammals. In most eukaryotes, including all vertebrates and a majority of the studied fungi/yeast, the site on antizyme mRNA where the shift occurs is UCC-UGA. The mechanism of the frameshift on this sequence likely involves nearly universal aspects of the eukaryotic translational machinery. Nevertheless, a mammalian antizyme frameshift cassette yields predominantly −2 frameshift in Saccharomyces cerevisiae, instead of the +1 in mammals. The recently identified endogenous S. cerevisiae antizyme mRNA has an atypical shift site: UGC-GCG-UGA. It is shown here that endogenous S. cerevisiae antizyme frameshifting is +1 rather than −2. We discuss how antizyme frameshifting in budding yeasts exploits peculiarities of their tRNA balance, and relate this to prior studies on Ty frameshifting.
Keywords
Polyamines are small organic cations and many of their known functions are related to this physico-chemical property. Polyamines are involved in many aspects of cellular function, among them binding and stabilizing RNA and DNA, protein synthesis, cell proliferation, and programmed cell death (Childs et al. 2003; Wallace et al. 2003).
The rate limiting step in the biosynthesis of polyamines is the conversion of ornithine to putrescine catalyzed by the enzyme ornithine decarboxylase (ODC). In metazoan and fungal cells, the most important regulator of ODC is the protein ornithine decarboxylase antizyme (antizyme) (Hayashi et al. 1996). Antizyme genes have been identified in organisms ranging from yeast to mammals. Mammals have three paralogs of antizyme—antizymes 1, 2, and 3 (for review, see Ivanov et al. 2000a).
Antizyme binds to, and inhibits, ODC activity, targeting it for ubiquitin-independent proteolysis by the 26S proteosome (Murakami et al. 1992; Zhang et al. 2003). Antizyme also inhibits the import of polyamines into cells (Sakata et al. 2000; Belting et al. 2003). In contrast, antizyme stabilizes another protein, antizyme inhibitor, by inhibiting its ubiquitination (Bercovich and Kahana 2004). Further antizyme 1 is a component that binds to cyclin D1, at least in prostate cells, and targets it for degradation (Newman et al. 2004), Drosophila antizyme regulates nuclear entry and the overall levels of the sex determination master switch, sex lethal (Vied et al. 2003) and gametogenetin binding protein 1 interacts with mammalian antizyme 3 (Zhang et al. 2005), which is only expressed in particular stages of male germ cell development (Ivanov et al. 2000c; Tosaka et al. 2000).
The expression of antizyme mRNA is intriguing. Decoding initiates at the start of a short ORF and then switches to a long overlapping ORF in the +1 frame, which does not have independent initiation (Matsufuji et al. 1995). The efficiency of the required +1 translational frameshifting is stimulated by free polyamine levels in the cell. The involvement of polyamines completes an autoregulatory feedback loop where high levels of ODC activity lead to higher polyamine levels, which result in antizyme synthesis and the subsequent down-regulation of ODC and polyamines (for review, see Coffino 2001). Ribosomal frameshifting is the sensor and modulator of polyamine levels, and also (Le Roy et al. 2005) is responsive to interferon levels.
The translational frameshifting event occurs on the last ORF1 sense codon, which in vertebrates is invariably UCC. This is followed by a UGA-(U) stop codon. After decoding the UCC serine codon, the ribosomes that shift frame continue translation at the next available codon, (U)-GAU, in the +1 frame relative to ORF1. Several cis-acting RNA sequences are important for efficient stimulation of the frameshifting. Without the stimulatory elements, the frameshifting efficiency is reduced more than an order of magnitude. Some of these elements are located upstream (5′) and others are downstream (3′) of the frameshift site (Matsufuji et al. 1995; Ivanov et al. 1998a, 2000b; Howard et al. 2001; Petros et al. 2005). The upstream element in vertebrate orthologs of antizyme 1 consists of −50 nt adjacent to the frameshift site. Both phylogenetic and molecular analyses indicate that this 5′ stimulatory element is modular and has three distinct components (Ivanov et al. 2000a). These three modules have evolved gradually, with the module closest to the frameshift site evolving first and the distal one last. The frameshift stimulatory signal 3′ of the shift site in vertebrate antizyme 1 mRNA is a nearby pseudoknot. Its antizyme 2 counterpart is very similar. Different 3′ signals are present in invertebrates, including, in some, a different form of pseudoknot (Ivanov et al. 2004). A 3′ signal in Schizosaccharomyces pombe is poorly characterized but extends for up to 150 nt from the shift site (Ivanov et al. 2000b). On the basis of phylogenetic comparisons, other 3′ signals are suspected to exist in certain branches of nematodes and in fungi. The different 3′ elements apparently evolved independently.
Frameshifting sites of budding yeast antizyme mRNAs
The frameshift site in Saccharomyces cerevisiae antizyme mRNA is (GC)-GCG-UGA-C (Palanimurugan et al. 2004). A BLAST search in GenBank identified antizyme genes from 11 other yeast species that are closely related to S. cerevisiae (Fig. 1). In the following comparison, the Yarrowia lipolytica sequence is used as the outlier in the group. Y. lipolytica is the closest relative to S. cerevisiae (Cai et al. 1996), whose antizyme mRNA nevertheless has a UCC-UGA shift site, the main (higher) eukaryotic shift site. A number of other yeast/fungal antizyme genes are also known but are not considered here. Comparing antizyme genes of “Saccharomyces related” yeasts leads to several conclusions. Analysis of the S. cerevisiae frameshift site, which included utilization of computer RNA folding programs and comparison to nonbudding yeast antizyme frameshift sites, has suggested the presence of minimal cis-stimulatory sequences. This inference is further supported by comparing the S. cerevisiae frameshifting site to corresponding regions of antizyme genes from the 11 other related yeast antizymes. A total of only 8 nt are absolutely conserved in all 12 species in a region of 100 nt flanking the frameshift site. Seven of those eight are in the most conserved 12-nt region that is completely conserved between mammals and S. pombe. Additionally, 12 other nucleotides are conserved in at least 10 of 12 species examined here. Curiously, 6 of these 12 highly conserved nucleotides, and an additional one which is completely conserved, are also conserved in the antizyme genes of three fission yeasts—S. pombe, S. octosporus and S. japonicus. All 7 nt are located 5′ of the frameshift site (see Fig. 1). Although at this point speculative, it seams possible that these 7 nt play a role in the stimulation of frameshifting.
Two other highly conserved nucleotides also appear to be involved in stimulation of the frameshifting. These are the “CA” pair immediately 3′ of the UGA stop codon. A number of studies have shown that the nucleotide immediately 3′ of the stop codon is crucial for determining the efficiency of translation termination in both eukaryotes and prokaryotes (Tate et al. 1995). For most genes, efficient termination is advantageous and so the nucleotide least likely to promote efficient termination is the one most highly selected against in this position. In S. cerevisiae, and likely in other closely related yeasts, this nucleotide is “C” (Jacobs et al. 2002). Other experiments have shown that the sequence CA(A/G) located 3′ of a stop codon can induce a significant level of translational readthrough in S. cerevisiae (Namy et al. 2001). It is reasonable to suppose that the dinucleotide pair CA following the UGA stop codon decreases termination efficiency, thereby stimulating +1 frameshifting. Selective pressure on the 3′ nucleotide flanking the stop codon of antizyme ORF1, to maintain an inefficient termination context, is also apparent in other antizyme genes (Ivanov et al. 2000a).
While eight of 12 budding yeasts have GCG 5′ adjacent to the ORF1 UGA (giving the GCG UGA C frameshift site in S. cerevisiae), three have CCG at the corresponding position (and, as introduced above, one has UCC).
Shifting to the +1 frame can involve −2 or +1 frameshifting
Expression of a cassette with the mammalian antizyme 1 mRNA shift signals by mammalian translation components involves only +1 frameshifting. However, its heterologous expression in S. cerevisiae yields a high level of predominately −2 frameshifting at the same site, i.e., with an extra amino acid encoded at the shift site (Matsufuji et al. 1996). In contrast, expression of the same cassette in S. pombe yielded predominately +1 frameshifting (Ivanov et al. 1998b). The subsequent discovery of the endogenous S. pombe antizyme, which shares 12 nt encompassing the frameshift site with mammalian antizymes, seems likely to utilize +1 rather than −2 shift. The native S. cerevisiae antizyme gene, with its (GC)-GCG-UGA-C site, poses the question whether its expression involves −2 or +1 frameshifting (because of the heterologous expression result just mentioned and because a −2 shift would result in the new P-site codon being GCG, the same as the initial zero frame codon). −2 frameshifting is utilized in the expression of Mu and certain Mu-like phages (Xu et al. 2004). With the Mu sequence, CGG-GGG-CGA, repositioning of the ribosomal A-site tRNA (anticodon 3′GCI5′) from pairing with CGA to GGC seems to play an important role in the frameshifting being −2 (Xu et al. 2004; Baranov et al. 2006), in contrast to earlier studies with different types of model systems not involving inosine, where the slow-to-decode nature of the A-site was its important feature (Weiss et al. 1987, 1990).
The +1 frameshift site utilized in decoding the S. cerevisiae transposable element Ty3 is GCG-AGU-U (Farabaugh et al. 1993). In S. cerevisiae AGU is decoded by a rare arginine isoacceptor tRNA and involves a translational pause, thereby facilitating the frameshifting (Pande et al. 1995). In antizyme mRNA frameshifting, UGA performs the analogous stimulatory function. S. cerevisiae does not have a tRNA whose anticodon can form full Watson–Crick, or wobble, pairing with GCG (which is 5′ adjacent to the AGU). Instead the anticodon of the tRNAAla that decodes GCG, 3′CGI5′, uses an unusual G:I (purine:purine) apposition (Crick’s original, 1966, wobble pairing rules had I pairing with U, C, and A.). Involvement of poor pairing of this codon and anticodon is critical for frameshifting (Sundararajan et al. 1999; Stahl et al. 2001, 2004).
CCG is another codon that, at least in S. cerevisiae, lacks an exactly matching tRNA isoaccepter. It too has been shown to induce +1 frameshifting if followed by a rare codon (Vimaladithan and Farabaugh 1994; Sundararajan et al. 1999).
S. cerevisiae antizyme frameshifting is +1
The product derived from an S. cerevisiae antizyme frameshift cassette expressed in S. cerevisiae was investigated to determine whether the frameshifting at GC-GCG-UGA-C, is −2, +1, or some combination of the two. The last eight sense codons of ORF1 plus 60 nt downstream of the UGA stop codon were inserted between the BstEII and KpnI sites of the plasmid pGL107 (Matsufuji et al. 1996). An in-frame control was also generated, corresponding to the +1 frameshift product. After purification on a GST column, the frameshift protein product for the “wild-type” construct was further purified on a size-exclusion column to remove the protein product resulting from termination at the ORF1 stop codon. The protein products of the wild-type and +1 in-frame control were then analyzed by mass spectroscopy (Fig. 2). The mass of the in-frame +1 control, 144,950.7 Da, is different from the mass of the wild-type product, which is 144,961.5 Da, by a little over 10 Da (for an −145 kDa protein). A −2 frameshift product, in contrast is expected to be −174 Da heavier that the +1 control, the difference of the extra arginine encoded by CGU. The results provide strong evidence that the frameshifting required for expression of the endogenous antizyme is primarily +1. Although no traces of −2 frameshift product are apparent, the current analysis cannot exclude the possibility of very low levels of −2 frameshifting obscured by the limitations of mass spectrometry or of −2 frameshifting dependent on additional flanking antizyme sequences. Notwithstanding that caveat, the frameshifting detected is +1 and the reason for this directionality instead of what appears to be a good alternative −2 codon, anticodon pairing possibility, merits future investigation.
tRNA genes relevant to antizyme frameshifting in other budding yeasts
The data from finished, and the almost finished, yeast genome sequences were searched for tRNA genes using the “tRNAscan-SE 1.23” program (Lowe and Eddy 1997). Of the 12, only Kluyveromyces thermotolerans was judged to have insufficient sequence for this kind of search. Results of the analysis of the other 11 are shown in Table 1. As previously reported, no tRNAAla with an anticodon CGC exists in S. cerevisiae (which has a GCG alanine codon as its antizyme shift site). A gene for this tRNA species could not be detected also in S. paradoxus, S. bayanus, S. mikatae, S. kudriavzevii, C. glabrata, and S. castellii (all of which have a GCG alanine codon 5′ adjacent to their UGA). Correspondingly, in S. kluyveri and K. waltii (which have a CCG proline codon in place of GCG), no tRNAPro with an anticodon 3′GGC5′ was detectable. However, A. gossypii (with a GCG alanine codon 5′ adjacent to its UGA) has two genes for tRNAAla with the anticodon CGC. When tRNAAla with the anticodon CGC is artificially overexpressed in S. cerevisiae, this significantly reduces +1 frameshifting efficiency on the otherwise shift-prone sequence GCG-AGG-C (Sundararajan et al. 1999). If the presence of two genes for tRNAAla with the anticodon CGC in A. gossypii lead to somewhat reduced frameshifting, this is perhaps counteracted by other features of its antizyme gene. For example, its ORF2 has an unusually high G:C content—66% versus 32% in S. cerevisiae. For the two genomes as a whole the G:C content is 52% in A. gossypii and 38% in S. cerevisiae (Dietrich et al. 2004). Not surprisingly, a number of stable RNA structures can be predicted 3′ of the antizyme frameshift site of A. gossypii. Perhaps the GCG-UGA frameshift site there is not a strongly +1 shift-prone site on its own, but this is compensated by a stimulatory 3′ structure.
Y. lipolytica, like the fission yeast S. pombe, has a UCC serine codon 5′ adjacent to its ORF1 UGA. It is not surprising that it does not have a CGC or CCG shift site, as it has two genes for each of the alanine and proline tRNAs at issue. Y. lipolytica does not have the general budding yeast shift mechanism specialization but is more like fission yeast and vertebrates in this regard.
How does the frameshifting utilized by certain of the budding yeasts occur? If P-site tRNA re-pairing to mRNA at an overlapping codon is involved, what are the relative strengths of the original and frameshifted pairings? What is the relative efficiency of productive shifting to the +1 frame compared to shifting to the −1 frame? For instance, P-site tRNA re-pairing to give −1 frameshifting with the K. waltii sequence, UGC CCG UGA, would involve good first and second position pairing at CC. In contrast, important second position pairing at the +1 frame codon CGU is problematic. The same applies for the first position with any −2 frameshifting involving pairing to GCC. Understanding the native frameshifting in these organisms is relevant to the controversial issue as to whether shifting to the +1 frame always involves dissociation and repairing, or in a major way can occur without slippage by occlusion of the mRNA base 3′ adjacent to the zero-frame P-site codon (Stahl et al. 2001; Ivanov et al. 2003; Hansen et al. 2003; Baranov et al. 2004). This task will take time since tRNA modification and binding studies are needed in addition to protein product characterization. Irrespective of the outcome of that issue, the data presented here strongly support an idea implicit in the work of Sundararajan et al. (1999), namely, that low abundance or a complete absence of a particular elongator tRNA can be a predictor that the corresponding codon is prone to frameshift when present in the P-site of the ribosome. Similar conclusions can also be inferred from even earlier experiments (Atkins et al. 1979; Weiss and Gallant 1983). Thus, frameshifting in expression of native antizyme mRNA in S. cerevisiae utilizes an efficient +1 frameshift mechanism in that organism in distinction to the mechanism common from S. pombe to mammals.
It has been suggested that ciliates frequently utilize +1 frameshifting in the expression of their genes (Klobutcher and Farabaugh 2002; Karamysheva et al. 2003). It remains to be determined if these organisms will also exhibit frameshift evolutionary specialization based on tRNA availability.
Analysis for the presence or absence of genes for tRNAAla 3′CGC5′ and tRNAPro 3′GGC5′ in the yeast Saccharomyces cerevisiae, Saccharomyces paradoxus, Saccharomyces bayanus, Saccharomyces mikatae, Saccharomyces kudriavzevii, Candida glabrata, Saccharomyces castellii, Ashbya gossypii, Saccharomyces kluyveri, Kluyveromyces waltii, and Yarrowia lipolytica

Nucleotide sequence comparison of the frameshift site of S. cerevisiae-related antizyme mRNAs. Species names are abbreviated as follows: S.ce., Saccharomyces cerevisiae; S.pa., Saccharomyces paradoxus; S.ba., Saccharomyces bayanus; S.mi., Saccharomyces mikatae; S.ku., Saccharomyces kudriavzevii; C.gl., Candida glabrata; S.ca., Saccharomyces castellii; A.go., Ashbya gossypii; S.kl., Saccharomyces kluyveri; K.th., Kluyveromyces thermotolerans; K.wa., Kluyveromyces waltii; Y.li., Yarrowia lipolytica. Absolutely conserved nucleotides are highlighted in red. Highly conserved nucleotides are highlighted in blue. The highly conserved nucleotides also present in S. pombe, S. octosporus, and S. japonicus are indicated by an asterisk underneath. The frameshift site is underlined.
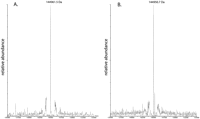
Mass-spectroscopic results. Increased mass is shown on the x axis and abundance of product on the y axis. (A) Molecular mass spectrum of “wild-type” antizyme frameshift cassette product fused between GST and β-gal. (B) Molecular mass spectrum of +1 in-frame control.
Acknowledgments
We thank Ken Wolfe from Trinity College Dublin for discussions on yeast phylogenetics and Barry Moore from the University of Utah for help with the tRNA gene analysis. This work was supported by an award from Science Foundation Ireland and NIH grants GM48152 to J.F.A. and GM71853 to R.F.G.
Footnotes
-
Article published online ahead of print. Article and publication date are at http://www.rnajournal.org/cgi/doi/10.1261/rna.2245906.
-
- Accepted November 28, 2005.
- Received September 30, 2005.
- Copyright 2006 by RNA Society








