
NMD takes two PIN domains to tango
- Department of Chemistry, Biochemistry and Pharmaceutical Sciences, University of Bern, CH-3012 Bern, Switzerland
- Corresponding author: oliver.muehlemann{at}unibe.ch
Abstract
Three new studies (Arpa et al., J Mol Biol 169734 [2026]; Kurscheidt et al., Nat Commun 17: 1934 [2026]; Modena et al., RNA, this issue [2026]) found that the PIN domains of SMG5 and SMG6, two proteins involved in nonsense-mediated mRNA decay (NMD), interact and show that the physiological endonucleolytic activity initiating the degradation of the nonsense mRNA is exerted by this composite catalytic site.
Keywords
Nonsense-mediated mRNA decay (NMD) is a central translation-coupled RNA surveillance pathway that protects cells from aberrant transcripts and shapes gene expression programs (Karousis and Muhlemann 2022; Monaghan et al. 2023). It has been known for more than 20 years that degradation of mRNAs targeted by NMD is initiated by an endonucleolytic cleavage near the NMD-triggering termination codon in Drosophila melanogaster cells (Gatfield and Izaurralde 2004). This was confirmed in human cells (Eberle et al. 2009) and recently in Caenorhabditis elegans (Viscardi et al. 2025). Intriguingly, two of the metazoan NMD factors—SMG5 and SMG6—contain PIN (PilT N-terminal) domains, which possess ribonuclease activity in bacterial toxin-antitoxin systems (Arcus et al. 2011) and structurally resemble FLAP family endonucleases, such as RNase H (Glavan et al. 2006). However, only the PIN domain of SMG6 harbors the conserved triad of acidic amino acids (D1251, D1353, and D1392 in human SMG6), which are necessary for the catalytic activity of other PIN domains (Glavan et al. 2006; Senissar et al. 2017). In contrast, the PIN domain of SMG5, while adopting a very similar overall structure, contains only one of these three acidic residues (Glavan et al. 2006). Accordingly, SMG6 was identified as the endonuclease required for initiating mRNA decay in NMD (Huntzinger et al. 2008; Eberle et al. 2009). Mutation of any of the three conserved aspartates in the SMG6 PIN domain inhibited the degradation of NMD-sensitive transcripts (Huntzinger et al. 2008; Eberle et al. 2009). While the SMG5 PIN domain, as expected, showed no nuclease activity in vitro (Glavan et al. 2006), recombinant SMG6-PIN could cleave circular RNAs in vitro (Eberle et al. 2009), demonstrating its endonuclease activity. However, 25 pmol of SMG6-PIN were required in this assay to digest 0.01 pmol RNA in 2 h, begging the question of why this enzymatic reaction was so inefficient. What was missing in this assay?
It took 17 years and the development of AlphaFold to answer this question: Three recent studies now demonstrate that the missing piece was the PIN domain of SMG5 (Arpa et al. 2026; Kurscheidt et al. 2026; Modena et al. 2026). These studies independently originated from AlphaFold models, which predicted with high confidence an interaction between the PIN domains of SMG5 and SMG6 that is conserved across humans, flies, and worms (Fig. 1A). The structural models indicated that the ribonuclease center is jointly formed by the SMG6 PIN domain, including its N-terminal α-helical extension and a stretch located between the 14-3-3-like and the α-helical domains (amino acids 831–874 in human SMG6), and the SMG5 PIN domain with its own N-terminal α-helical extension. This composite interface is fully accessible for RNA binding, and the three conserved aspartates of the SMG6 PIN together with a conserved aspartate of the SMG5 PIN (D893 in human SMG5) are ideally positioned to coordinate two bivalent cations required for the nucleophilic attack on the RNA (Mn2+ in in vitro assays).
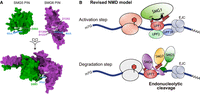
(A) Surface rendering of the SMG5 (green) and SMG6 (purple) PIN domains. The interaction surfaces (upper part) and the complex (lower part) of both PIN domains are depicted. The catalytic center of the composite endonuclease is indicated by the positions of the RNA, the conserved aspartates, and the two Mn2+ cations (courtesy of Stefanie Jonas). (B) Revised working model for metazoan NMD (adapted from Karousis and Muhlemann 2022). Translation termination at a premature termination codon (PTC) triggers SMG1-mediated hyperphosphorylation of UPF1, facilitated by UPF2, UPF3B, and the EJC (activation step). Hyperphosphorylated UPF1 recruits SMG6 and SMG5:SMG7 to the RNA, allowing assembly of the composite SMG5–SMG6 PIN endonuclease, which cleaves the target mRNA.
The structural model of SMG5 and SMG6 forming a composite active site was subsequently verified biochemically: in vitro RNA cleavage assays confirmed that the N-terminally extended SMG5 PIN domain substantially increased the ribonuclease activity of SMG6, dependent on SMG5-D893 (Arpa et al. 2026; Kurscheidt et al. 2026). Moreover, mutations predicted to weaken or abolish the PIN–PIN interaction inhibited the ribonuclease activity, and compensatory mutations on the opposite side of the interaction interface restored activity in these assays (Arpa et al. 2026; Kurscheidt et al. 2026).
The findings were also confirmed in vivo in C. elegans (Modena et al. 2026) and in human cells (Kurscheidt et al. 2026). Consistent with earlier evidence that the SMG5 PIN is required for SMG6 activity in human cells (Boehm et al. 2021), Modena and colleagues demonstrated that the PIN domain of SMG5 is also indispensable for NMD in C. elegans (Modena et al. 2026). Paralleling the results of the in vitro analyses, structure-guided mutagenesis in C. elegans and human cells revealed that disrupting the interface between the SMG5 and SMG6 PIN domains abolishes NMD, whereas compensatory mutations that restore the predicted PIN–PIN interaction rescue NMD activity (Kurscheidt et al. 2026; Modena et al. 2026). Overall, these findings document the requirement of assembling the composite PIN endonuclease for functional NMD in metazoans.
Together, these studies overturn the long-standing notion that SMG5 and SMG6 act in parallel decay pathways (Muhlemann and Lykke-Andersen 2010). Instead, they uncovered that NMD relies on a split catalytic center assembled from the PIN domains of SMG5 and SMG6. This discovery represents a major leap in elucidating the molecular mechanism of NMD in metazoans. It reveals an elegant regulatory principle that ensures tight spatial and temporal control of the endonuclease activity during NMD. Cleavage of a UPF1-bound mRNA occurs only when SMG5 and SMG6 are properly engaged. SMG5 forms a heterodimer with SMG7 (Jonas et al. 2013) that binds phospho-epitopes in the C-terminal region of UPF1 (Ohnishi et al. 2003; Fukuhara et al. 2005; Jonas et al. 2013), whereas SMG6 interacts with an N-terminal phospho-epitope and the helicase stalk region of UPF1 (Fig. 1B; Okada-Katsuhata et al. 2012; Chakrabarti et al. 2014; Nicholson et al. 2014). The PIN domains of SMG5 and SMG6 form the composite catalytic site only in such correctly assembled RNA-bound phospho-UPF1-SMG6-SMG5:SMG7 complexes, ensuring specificity in NMD. Beyond resolving a central mechanistic puzzle in mRNA surveillance, the three new studies represent an efficient strategy for controlling potentially dangerous enzymatic activities: the catalytic domain is split across two proteins and activated only when they engage in the correct molecular partnership.
ACKNOWLEDGMENTS
We are grateful to Stefanie Jonas (ETH Zürich, Switzerland) for her help with Figure 1A. The research of the authors’ laboratory is supported by the National Center of Competence in Research (NCCR) on RNA & Disease, funded by the Swiss National Science Foundation (SNSF; grant 51NF40-205601), by the SNSF project grant 3200-0-239917, by the NOMIS Foundation, and by the Canton of Bern (University intramural funding).
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.081027.126.
-
Freely available online through the RNA Open Access option.
This article, published in RNA, is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








