
Circuit logic: interdependent RNA modifications shape mRNA and noncoding RNA structure and function
- Stem Cell Program and Division of Hematology/Oncology, Boston Children's Hospital, Boston, Massachusetts 02215, USA
- Department of Stem Cell and Regenerative Biology, Harvard University, Cambridge, Massachusetts 02138, USA
- Corresponding author: jennifer.porat{at}childrens.harvard.edu
Abstract
Continued advances in high-throughput detection of posttranscriptional RNA modifications have enabled large-scale, mechanistic studies into the importance of RNA modifications in regulating the structure, function, and stability of coding and noncoding RNAs. More recently, this has expanded beyond investigations of independent single modifications, revealing the breadth of modification complexities in single transcripts and the biogenesis pathways involved that lead to coordinately modified RNA species. This has resulted in the concept of modification circuits, where one modification can promote or inhibit the subsequent installation of other modifications, or when modifications are coordinated across different RNA species. These circuits play important roles in the biogenesis of multistepped posttranscriptional modifications, modulate ribonucleoprotein complex formation and conformational switches, and mediate codon-biased translation through the coordination of mRNA and tRNA modifications. Here, I review evidence of complex modification circuits in mRNA and noncoding RNA and highlight open questions concerning the molecular mechanisms giving rise to modification circuits and their importance in the context of RNA processing and maturation.
Keywords
RNA-BINDING PROTEINS COORDINATE AND CARRY OUT RNA MODIFICATIONS
RNAs undergo an extensive maturation process that can involve 5′ capping, 3′ end processing or trimming, splicing, and critically, the addition of chemical modifications. The past decade has seen a tremendous surge in our collective understanding of the breadth and importance of these posttranscriptional RNA modifications, brought about in large part by the combination of classic RNA biochemistry and high-throughput sequencing (Wiener and Schwartz 2020). Posttranscriptional RNA modifications, which are as numerous as they are chemically diverse, occur on mRNA and noncoding RNAs, where they have roles ranging from modulating RNA structure to serving as recognition sites for RNA-binding proteins (RBPs). More than 170 RNA modifications have been described (Machnicka et al. 2013), ranging from chemically simple methylations to the addition of more complex functional groups that require several enzymatic steps for their synthesis (Helm and Alfonzo 2014).
While the molecular and chemical mechanisms of each processing step vary greatly depending on the nature of each reaction, a unifying facet is the involvement of RBPs, which can provide target specificity, mediate recruitment to certain transcripts, position the target base for modification, and catalyze said modification. Over 1000 RBPs have been identified and validated in humans alone (Gerstberger et al. 2014), and the number has only continued to climb with recent proteome-wide studies (Hentze et al. 2018). Among these are RNA modification enzymes, which often recognize specific sequences or structural motifs on their targets. Well-studied examples of this are the pseudouridine synthase Pus7, which recognizes a UGUAG motif (the underlined uridine being the target of modification) (Carlile et al. 2014; Safra et al. 2017) and Pus1, which recognizes the target uracil at the 5′ end of a bulged stem–loop (Carlile et al. 2019). That these sequence and structural elements are present in mRNA and tRNA provides an explanation as to how a single modification enzyme can target a diverse array of RNA substrates. In contrast, select RNA modification enzymes possess catalytic activity only and must rely on RBP cofactors to recognize their targets. tRNA and mRNA acetylation is carried out by the acetyltransferase Kre33/Nat10 (nomenclature refers to the yeast and human homologs, respectively) and the noncatalytic cofactor Tan1/THUMPD1, which serves as an adaptor for RNA binding (Sharma et al. 2015). Similarly, m7G modification of tRNA is catalyzed by the methyltransferase METTL1, while the RBP WDR4 acts as a scaffold for METTL1 and the T arm of the tRNA (Li et al. 2023; Ruiz-Arroyo et al. 2023).
But while RBPs and RNA modification enzymes often cooperate to ensure RNA structure and functionality, the installation of multiple modifications in a single RNA molecule requires precise temporal and spatial coordination. tRNAs are perhaps the best-studied example of multimodified RNA, with each transcript containing an average of eight and 13 posttranscriptional modifications in bacteria and eukaryotes, respectively (for review, see Zhang et al. 2022). However, the use of modification-specific detection methods, such as chemical derivatization techniques that can only detect a single type of modification, has made it challenging to assess how multiple modifications are regulated in a coordinated manner. Despite this, continuous improvements in sequencing technologies have enabled the simultaneous detection of multiple posttranscriptional modifications (for review, see Begik et al. 2022; White and Hesselberth 2022). Such studies have validated and uncovered new modification “circuits,” in which the deposition of a given modification is a prerequisite for the installation of a subsequent modification, or in some cases, inhibitory to others. Recent evidence suggests that such modification circuits are widespread among diverse RNA species, both mRNA and noncoding RNA (Guy et al. 2012; Guy and Phizicky 2015; Arimbasseri et al. 2016; Barraud et al. 2019; Behrens et al. 2021; Watkins et al. 2022; Hernandez-Alias et al. 2023; Yared et al. 2023; Huang et al. 2024; Lucas et al. 2024; Shaw et al. 2024; Hwang et al. 2025; Zhang et al. 2025).
In this review, I discuss recent technological advances enabling profiling of multiple modifications in a single transcript, mechanistic studies revealing how multiple modifications are coordinated, how these modification circuits regulate RNA processing activities and ordered RBP binding, and how they are in turn regulated by ribonucleoprotein (RNP) complex assembly.
RNA MODIFICATION DETECTION METHODS
Insights into RNA modification circuits were brought about in large part due to advances in mass spectrometry (Wein et al. 2020; Jones et al. 2023) and Illumina-based (Behrens et al. 2021; Watkins et al. 2022; Hernandez-Alias et al. 2023) and nanopore-based (Thomas et al. 2021; Lucas et al. 2024; Shaw et al. 2024; White et al. 2024) RNA sequencing, which use mutational signatures from reverse transcription or base-calling errors, respectively, to identify and locate multiple modifications in a single read. To date, no single method can detect the full repertoire of RNA modifications. Mass spectrometry can detect multiple modifications in a single run, provided that a mass standard exists, the modification is sufficiently abundant for detection, and the modified nucleoside possesses a unique mass that can be differentiated from the unmodified or differentially modified nucleoside (Wetzel and Limbach 2016). In the absence of additional chemical treatments, Illumina-based RNA sequencing only reports on Watson–Crick face methylations (m1A, m1G, m22G, and m3C) (Watkins et al. 2022) and the degree to which modification cross talk can be assessed is dependent on the sequencing method, as the reverse transcriptases TGIRT (Behrens et al. 2021) and SuperScript IV (Watkins et al. 2022) display different quantitative mutation signatures for different modifications (Hernandez-Alias et al. 2023). Further, current measurements by nanopore direct RNA sequencing are not quantitative for tRNA, as they report mismatch probability values rather than an absolute fraction of modified molecules (Shaw et al. 2024), necessitating the use of orthogonal modification mapping methods to fully understand the breadth and extent of modification circuits. One way to achieve fully quantitative nanopore measurements involves the use of synthesized standards containing a given modification in different stoichiometries and sequence contexts—including sequence contexts containing other modifications—to train models to recognize specific modification combinations (Shaw et al. 2024), as has been done for certain mRNA modifications (Huang et al. 2024).
MODIFICATION CIRCUITS IN tRNA
Modification circuits in the tRNA anticodon loop
Prokaryotic and eukaryotic tRNAs are heavily posttranscriptionally modified, although modifications tend to cluster in the anticodon loop, where they can affect mRNA decoding by influencing codon–anticodon base-pairing (for review, see Tuorto and Lyko 2016). With such a high density of modified bases, it is unsurprising that the presence of a particular modification can influence the deposition or exclusion of another. Well-studied examples include 2′-O-methylation at C32 and G34, which promote wybutosine formation at G37 in phenylalanine tRNAs (Guy et al. 2012; Guy and Phizicky 2015), and the modifications N6-isopentyladenosine (i6A) or N6-threonylcarbamoyladenosine (t6A) at position 37, which stimulate m3C32 modification in serine and threonine tRNAs (Fig. 1; Arimbasseri et al. 2016; Han et al. 2017). It has been hypothesized that these modification circuits act as a recognition mechanism to differentiate tRNAs that have highly similar anticodon loop sequences (for review, see Han and Phizicky 2018). In support of this idea, the wobble uridine (U34) in yeast tRNA can remain unmodified, methylated, pseudouridylated, deaminated, or modified to methoxycarbonylmethyl-2-thiouridine (mcm5s2). Therefore, a recognition mechanism that relies on preexisting modifications, rather than sequence alone, might be required to generate modification specificity (Han and Phizicky 2018). A nonmutually exclusive explanation for modification circuits in the anticodon loop is the idea that an earlier-occurring modification may alter the structure of the anticodon loop, such that another base is more accessible for a subsequent modification (Han and Phizicky 2018).
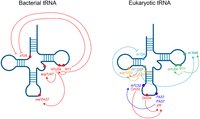
Modification circuits in bacterial and eukaryotic tRNAs. Modification sites and associated positive and inhibitory cross talk in tRNA. Modifications that are linked by a circuit are highlighted in the same color. Figure created in BioRender.
Modification circuits in the tRNA body
While much work has been focused on uncovering modification circuits in the anticodon loop, recent evidence suggests that modifications in the tRNA body, which primarily affect tRNA structure and stability, abide by similar logic. The largest number of evolutionarily conserved tRNA modifications is found in the T-loop and as such, T-loop modification circuits are likewise seen across diverse species. The bacterial pseudouridine synthase TruB, responsible for the nearly universally modified Ψ55, preferentially binds and modifies unmodified tRNA, while the methyltransferase TrmA, which catalyzes m5U54, exhibits tighter binding to tRNA already modified with Ψ55, suggesting that prior modification by TruB promotes subsequent modification by TrmA (Schultz and Kothe 2020). In line with the model that TrmA and TruB act early in the tRNA biogenesis pathway and influence downstream modifications, Illumina RNA sequencing, mass spectrometry, and northern blotting revealed that knockout of TrmA results in a loss of the tRNA modifications acp3U47 and ms2i6A37 (Jones et al. 2024; Schultz et al. 2024), and knockout of TrmA or TruB leads to an increase in s4U8 (Fig. 1; Schultz et al. 2024). Further, tRNAs that display altered acp3U47 or ms2i6A37 decode at least one codon differently in TrmA knockout cells, pointing to the importance of body modification circuits in modulating codon-specific translation (Schultz et al. 2024). Similar circuits have been observed in yeast using time-resolved nuclear magnetic resonance (NMR) and nanopore direct RNA sequencing, where Ψ55 occurs first, promoting the subsequent modifications m5U54 (Barraud et al. 2019; Yared et al. 2023; Lucas et al. 2024) and m1A58 (Fig. 1; Barraud et al. 2019; Lucas et al. 2024; Shaw et al. 2024).
Beyond the T-loop, several other modification circuits within the tRNA body have been described, with a given modification either promoting or inhibiting the installation of subsequent modifications. Illumina-based sequencing methods reported several examples of modification cross talk involving m22G26, which is catalyzed by the methyltransferase Trm1/TRMT1, including stimulating m1A58 (Behrens et al. 2021; Hernandez-Alias et al. 2023) and inhibiting m1G9 (Behrens et al. 2021). Notably, only tRNAs with U7:A66, rather than G7:C66 at the base of the acceptor stem, exhibit inhibition of m1G9 by m22G26, suggesting that this circuit may modulate tRNA structure. Additional cross talk involves the TRMT1-like protein TRMT1L, which catalyzes m22G27 and promotes the deposition of an aminocarboxypropyl (acp3U) modification on tRNA TyrGUA (Hwang et al. 2025; Zhang et al. 2025). tRNA TyrGUA is the only m22G27-containing tRNA in humans, although loss of TRMT1L also results in decreased acp3U levels on the m22G27-lacking tRNA CysGCA and tRNA AlaCGC, which can be restored with expression of a catalytic-null TRMT1L mutant (Fig. 1; Hwang et al. 2025; Zhang et al. 2025). Notably, TRMT1L still interacts with CysGCA (Hwang et al. 2025), suggesting that TRMT1L itself, and not the m22G27 modification, is important in promoting acp3U.
It is interesting to note that enzymes installing modifications that occur earlier in tRNA biogenesis (Barraud et al. 2019; Schultz and Kothe 2020), and thus have the capacity to influence the deposition of subsequent modifications, have also been demonstrated to possess catalytic-independent RNA folding activities (Keffer-Wilkes et al. 2016, 2020). It remains to be found whether catalytic or catalytic-independent activities (or a combination of both) are required for modification circuits initiated by Ψ55 (TruB), m5U54 (TrmA), and m22G26 (Trm1), all of which have established roles in RNA folding that are independent from their catalytic activity (Keffer-Wilkes et al. 2016, 2020; Porat et al. 2023b). While it is possible that the local structural rearrangements mediated by these dual-function modification enzymes (Keffer-Wilkes et al. 2016) may better position certain bases for subsequent modifications in the circuit, this has not yet been tested.
MODIFICATION CIRCUITS AT A SINGLE SITE
While there is an increasing appreciation for the interplay of RNA modifications within and between transcripts, perhaps the most dramatic form of modification circuitry involves stepwise posttranscriptional modifications on a single base. One of the first examples of a posttranscriptional modification that relies on a prior modification of the same base was the discovery of the modified base queuosine (Q) in vertebrate tRNAs (Kasai et al. 1975; Kuchino et al. 1976; Okada et al. 1977; Zhao et al. 2023). Although Q modification, where guanine at position 34 undergoes a transglycosylation reaction (Chen et al. 2010), is widespread across evolution (Katze et al. 1982), only vertebrate tRNAs contain hypermodified Q. The tRNA glycosylases QTMAN and QTGAL catalyze the addition of a mannose to Q on tRNA Asp and galactose to Q on tRNA Tyr, respectively (Thumbs et al. 2020; Hillmeier et al. 2021; Zhao et al. 2023), where the bulky modification serves to slow ribosomes at cognate codons and can suppress stop codon readthrough (Zhao et al. 2023). The biogenesis of wybutosine (yW), a posttranscriptional modification found at position 37 of eukaryotic and archaeal phenylalanine tRNAs (Blobstein et al. 1973), is perhaps even more complex and intricate. Guanine at position 37 is first methylated to m1G by the methyltransferase Trm10, followed by a multistepped cascade catalyzed by the Tyw (tRNA yW-synthesizing protein) family of enzymes where (1) the tricyclic core is formed to generate the yW-187 intermediate (numbers in wybutosine intermediate names refer to the difference in mass between yW and intermediates), (2) an aminocarboxypropyl (acp) group is added to form yW-86, (3) methylation yields yW-14, (4) another methylation produces yW-58, and (5) methoxycarbonylation produces the final wybutosine (Noma et al. 2006). Wybutosine can also be further modified to hydroxywybutosine (Noma et al. 2010). Additional tRNA hypermodifications have been observed in Trypanosoma brucei tRNA ThrAGU, where the methyltransferase TRM140 catalyzes m3C modification at position 32, which is a prerequisite for subsequent C to U deamination by the deaminase ADAT2/3 (Rubio et al. 2017).
Complex hypermodifications are not unique to tRNAs, as yeast and human 18S rRNA (position 1191 in yeast and 1248 in humans, both of which are positioned next to the P-site tRNA) undergoes snoRNA-guided pseudouridylation, further methylation to N1-methylpseudouridine, and finally, Tsr3-catalyzed addition of an acp group to give rise to m1acp3Ψ (Meyer et al. 2016; Babaian et al. 2020). The DTWD family of acp transferases is also involved in RNA hypermodifications, with DTWD2-catalyzed modification at position 20 in human asparagine and isoleucine tRNAs (Takakura et al. 2019) serving as the site for N-linked glycosylation to generate glycoRNA (Flynn et al. 2021; Xie et al. 2024). It is likely that other RNA modifications are required to template RNA glycosylation, which will be an area of future research in the expanding field of glycoRNA.
CO-OCCURRENCE OF MODIFICATIONS IN mRNA
Discovery of the breadth and complexity of mRNA modifications prompted the idea that a single mRNA transcript may contain multiple modifications. While this has not yet been explored for most possible mRNA modification combinations, the simultaneous detection of multiple modifications by nanopore direct RNA sequencing, combined with machine learning, has revealed positive and inhibitory modification pairs in a single transcript. Nanopore simultaneous investigation for pseudouridine and m6A (NanoSPA) can detect pseudouridine and m6A at the same time, and when performed on wild-type and modification enzyme knockdown cell lines, revealed modification interdependencies (Huang et al. 2024). Transcripts possessing more pseudouridine contained less m6A-modified sites, while knockdown of the m6A writer METTL3 led to an increase in pseudouridine in transcripts with more m6A sites, indicating an inhibitory relationship between m6A and pseudouridine. Perhaps surprisingly, knockdown of the pseudouridine synthase TRUB1 resulted in decreased m6A levels, raising the possibility that TRUB1-modified pseudouridines promote m6A deposition, while the 12 other human pseudouridine synthases install pseudouridines that may inhibit m6A (Huang et al. 2024). Additionally, m6A and m3C occur in the same transcript in mRNA from HEK293 and HeLa human cancer cell lines, mouse cortex tissue, and rice (Acera Mateos et al. 2024; Wu et al. 2024), although it remains unknown whether one influences the other.
Still, it remains difficult to assess any causalities contributing to modification co-occurrence, such as whether either modification influences the installation of the other. Part of this difficulty arises from differential modification levels at individual sites and the idea that even in a single transcript, the deposition of each m6A occurs independently, as has been shown for m6A sites in β-actin mRNA (Leger et al. 2021). Thus, the degree to which one modification influences another may not be generalizable, but rather governed by modification penetrance, sequence specificity, or other factors unique to a single transcript. mRNA modifications, and whether they form circuits like tRNAs, are still poorly understood at both the mechanistic and functional levels. Continued developments in direct RNA sequencing to allow the detection of more modifications with higher confidence, such as the use of fully unmodified in vitro-transcribed transcriptomes to prevent false positive modification site calling (Tavakoli et al. 2023; Acera Mateos et al. 2024; McCormick et al. 2024), will lead to further insights into the possible role of co-occurring mRNA modifications.
MODIFICATIONS IN 7SK AND U6 snRNA AFFECT RNA FUNCTION AND RNP ASSEMBLY
Aside from structural alterations imparted by modifications, often through altering RNA secondary and tertiary structure, many of the functions assigned to modifications arise from the recruitment of RBPs to certain modifications. Given that some multimodified RNA species exist in dynamic RNP complexes, it stands to reason that modulating RNA modifications—perhaps through modification circuits or co-occurring modifications—may alter the equilibrium of various RNP complex states. This has been well studied for the 7SK small nuclear RNA (snRNA), which acts as a scaffold for RNP formation to control transcriptional regulation by sequestering positive transcription elongation factor b (P-TEFb) (for review, see Hasler et al. 2021). 7SK is present in the cell in several structurally, compositionally, and functionally distinct RNPs, including promoter-associated 7SK bound to HEXIM1/2, which is involved in P-TEFb sequestration and subsequent transcriptional downregulation (Nguyen et al. 2001; He et al. 2008; Markert et al. 2008); 7SK bound to hnRNP A1 and A2/B1 or hnRNP Q and R, which form upon transcription-dependent dissociation of the 7SK-P-TEFb-HEXIM complex (Barrandon et al. 2007; Van Herreweghe et al. 2007), and enhancer-associated 7SK bound to the ATP-dependent nucleosome remodeling complex BAF, which serves to repress enhancer RNA transcription (Fig. 2; Flynn et al. 2016).
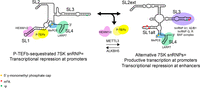
7SK snRNP complexes and conformations are associated with distinct RBPs and posttranscriptional modifications. Schematic of P-TEFb-sequestering and alternative 7SK snRNP complexes. The P-TEFb-sequestering 7SK snRNP is characterized by low levels of m6A, while hnRNP- and BAF-bound 7SK is methylated by METTL3 and adopts a different conformation. 7SK secondary structure adapted from Olson et al. (2022).
7SK contains several posttranscriptional modifications, including a single pseudouridylation site, Ψ250, catalyzed by Dyskerin (Zhao et al. 2016; Leger et al. 2021), and several m6A sites installed by the methyltransferase METTL3 and erased by ALKBH5 (Leger et al. 2021; Perez-Pepe et al. 2023; Wang et al. 2023). In line with the hypothesis that alternate modification states may underlie the conformational changes and RNP dynamics that modulate transcription, modified sites identified by both methods overlap with previously identified RBP-binding sites, including m6A at sites in stem–loop 1 (SLI) that interact with HEXIM1/2, and pseudouridylation and m6A sites in SLIII that overlap with hnRNP R, hnRNP A1, and hnRNP A2/B1 sites (Briese et al. 2018; Leger et al. 2021; Luo et al. 2021; Perez-Pepe et al. 2023; Wang et al. 2023). Overexpression of ALKBH5 targeted to 7SK with dCasRx, leading to a decrease in 7SK m6A, led to a reduction in nascent transcription and serine 2 phosphorylation in the C-terminal domain of RNA polymerase II, supporting a role for m6A in stimulating P-TEFb release and productive transcription elongation (Wang et al. 2023). m6A modifications in SLI, in particular, drive structural changes in 7SK, where unmethylated 7SK favors a P-TEFb-sequestering conformation that is associated with stronger 7SK-HEXIM1/2 interactions, and methylated 7SK promotes P-TEFb release, upon which 7SK can assemble into a new RNP with hnRNP proteins (Fig. 2; Perez-Pepe et al. 2023; Wang et al. 2023). Supporting the idea that m6A influences RBP binding, particularly for hnRNP family proteins, m6A in a hairpin of the long noncoding RNA MALAT1 disrupts hairpin structure to increase accessibility of an hnRNP C and hnRNP G-binding site (Liu et al. 2015, 2017). The structure of BAF-associated 7SK is distinct from HEXIM-bound 7SK and instead most closely resembles 7SK found in hnRNP-containing RNPs (Flynn et al. 2016), but the degree to which m6A may play a role in the BAF-7SK RNP remains unknown.
In contrast, the methyl phosphate capping enzyme (MePCE) and La-related protein 7 (LARP7), which are associated with 7SK snRNP assembly (Krueger et al. 2008) and are found in the P-TEFb-sequestered and -released RNPs, associate with 7SK independent of methylation status (Wang et al. 2023). Pseudouridylation is similarly found in both RNPs, as pseudouridylation is required for efficient formation of the P-TEFb- and hnRNP-containing RNPs and thus found in most cellular 7SK (Zhao et al. 2016). The degree of cross talk between pseudouridylation and m6A, and whether this influences the equilibrium of 7SK RNPs, remains to be found but represents a promising area of future research. But while methylation status correlates with distinct RNPs, questions remain as to whether m6A influences RBP binding and RNP assembly, or whether RBP-guided conformational changes in 7SK promote or inhibit m6A. In this sense, it is also interesting to consider the alternate hypothesis that modification circuits may exist as a function of RNP assembly pathways, where the ordered binding of a series of RBPs, with or without additional changes in subcellular localization, dictates the order of modification installation.
MePCE and LARP7 are also present in U6-containing snRNPs, where they influence a cascade of posttranscriptional modifications. MePCE installs a 5′ γ-monomethyl phosphate on U6 and 7SK (Singh and Reddy 1989; Jeronimo et al. 2007), which is thought to occur upstream of LARP7 binding, since LARP7 binds the catalytic core of MePCE to inhibit its capping function (Xue et al. 2009). Although the exact timing of posttranscriptional modifications with respect to RBP binding has not yet been worked out for U6 (for review, see Didychuk et al. 2018), METTL16-catalyzed installation of m6A—occurring in the central adenosine of the conserved ACAGAGA sequence that base-pairs with the 5′ splice site of pre-mRNA—occurs early in the U6 biogenesis pathway, before U6 has undergone 3′ end processing to form a 2′, 3′-cyclic phosphate in humans and fission yeast, or a terminal 3′ phosphate in budding yeast (Didychuk et al. 2017; Pendleton et al. 2017; Warda et al. 2017). U6 also undergoes LARP7- and MePCE-guided 2′-O-methylation (Hasler et al. 2020; Wang et al. 2020; Porat et al. 2023a), resulting in a network of RBP-influenced U6 modifications that ultimately coordinate and fine-tune splicing (Hasler et al. 2020; Wang et al. 2020; Ishigami et al. 2021; Porat et al. 2023a). Still, the interdependence of these modifications and processing events remains unknown, as are the precise roles MePCE and LARP7 play in coordinating the timing of such distinct modifications.
COORDINATION OF MODIFICATIONS ON MULTIPLE RNA SPECIES
In addition to modification cross talk existing within a single transcript, recent work revealed cases where modifications on several classes of transcripts are coordinated or coregulated. An example of this comes from modification enzymes that target different types of transcripts, such as pseudouridine synthases that modify tRNA and mRNA (Carlile et al. 2014,2019), the demethylase FTO which binds and regulates modifications in mRNA, tRNA, snRNA, snoRNA, and rRNA (Wei et al. 2018), and the 2′-O-methyltransferase Fibrillarin, which uses snorD101 to modify proline and glutamine tRNAs and rRNA (Zhang et al. 2023). The ability of a single enzyme or complex to carry out modifications on different classes of targets—particularly when both classes are involved in a common process like translation—suggests the possibility of modification coordination across RNA species. This has been demonstrated for the methyltransferase TRMT10A, which catalyzes m1G9 on tRNA and regulates m6A in mRNA through an interaction with FTO (Ontiveros et al. 2020). TRMT10A and FTO bind an overlapping set of mRNAs and TRMT10 enhances FTO demethylase activity, leading to selective demethylation of transcripts encoding proteins involved in mRNA processing and stability. Further, TRMT10-dependent m6A-containing mRNAs are enriched in codons decoded by m1G9-modified tRNAs, suggesting that TRMT10 coordinates mRNA and tRNA modifications to affect codon-biased translation, where particular codons are translated more efficiently as a function of the structure or abundance of the cognate tRNA (Ontiveros et al. 2020). Additional modification coordination in the context of translation regulation has been observed for rRNA and proline and glutamine tRNAs, which all interact with the 2′-O-methylation-guiding SNORD101, suggesting that components of the translation machinery may undergo coordinated modification regulation to fine-tune translation (Zhang et al. 2023). Similarly, dihydrouridine has been observed in tRNA, mRNA, and snoRNA, and notably, dihydrouridine was found in the C/D and H/ACA boxes of rRNA-modifying snoRNA (Draycott et al. 2022), raising the question of whether snoRNA modifications can affect downstream snoRNA-guided modifications in a regulatory cascade, thereby expanding the idea of modification circuits.
CONCLUSIONS AND FUTURE OUTLOOK
There exists an intricate network of RNA processing factors, including RBPs, RNA modification enzymes, and even other RNAs, that coordinate and compete to ultimately influence the fate and function of diverse classes of RNA. While the contribution of posttranscriptional modifications to RNA structure and function has been explored in great detail (for review, see Wiener and Schwartz 2020), the interplay between multiple modifications in a single molecule—or at a single site—has only recently begun to be explored in mechanistic detail. As RNA modifications rarely occur in isolation—which is particularly true for small noncoding RNA—these modification circuits act as intricate regulatory networks that guide the order and efficiency of RNA processing.
As we work toward improved methods capable of faithfully detecting multiple posttranscriptional modifications simultaneously (Alfonzo et al. 2021), several open questions remain concerning the mechanisms and functions of RNA modification circuits. To date, modification circuits have been characterized for the select group of modifications involved in the circuit, although it remains unknown how the disruption of a single modification circuit may also influence other circuits in the same molecule. This is particularly important for tRNA, where circuits exist in the D-loop, T-loop, and anticodon loop, in addition to the interactions between loops in the tertiary structure of a tRNA. Moreover, the extent to which mRNA modifications may regulate each other is still ambiguous, as is the extent to which modifications and their circuits affect other aspects of mRNA processing, including splicing, polyadenylation, and nuclear export. Finally, the molecular mechanisms underlying modification circuits have yet to be fully elucidated. One can speculate that earlier-occurring modifications induce structural changes that facilitate or inhibit downstream modifications, or that early modifications influence RBP binding that likewise promotes structural changes in the RNA.
Decades of research have begun to uncover the importance of RNA modifications, as has continued progress in the ability to quantitatively detect multiple modifications at once. With recent calls to promote the study of RNA modifications and development of tools to detect them (National Academies of Sciences, Engineering, and Medicine 2024), we can expect that discoveries to come over the next few years will continue to revolutionize our understanding of the complexities and interconnected nature of RNA modifications.
ACKNOWLEDGMENTS
I would like to thank Ryan Flynn, Mark Bayfield, Richard Maraia, Stephen Rader, Mohammadaref Talebi, and Tom Cech for critical reading of the manuscript, Jacob Fine for helpful discussions, and Christopher Watkins for the title suggestion.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080421.125.
-
Freely available online through the RNA Open Access option.
This article, published in RNA, is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








