
Principles, functions, and biological implications of m6A in plants
- Peizhe Song1,4,
- Zhihe Cai1,4 and
- Guifang Jia1,2,3
- 1Synthetic and Functional Biomolecules Center, Beijing National Laboratory for Molecular Sciences, Key Laboratory of Bioorganic Chemistry and Molecular Engineering of Ministry of Education, College of Chemistry and Molecular Engineering, Peking University, Beijing 100871, China
- 2PKU-Tsinghua Center for Life Sciences, Peking University, Beijing 100871, China
- 3Beijing Advanced Center of RNA Biology, Peking University, Beijing 100871, China
- Corresponding author: guifangjia{at}pku.edu.cn
-
↵4 These authors contributed equally to this work.
Abstract
Over the past decade, N6-methyladenosine (m6A) has emerged as a prevalent and dynamically regulated modification across the transcriptome; it has been reversibly installed, removed, and interpreted by specific binding proteins, and has played crucial roles in molecular and biological processes. Within this scope, we consolidate recent advancements of m6A research in plants regarding gene expression regulation, diverse physiologic and pathogenic processes, as well as crop trial implications, to guide discussions on challenges associated with and leveraging epitranscriptome editing for crop improvement.
Keywords
- N6-methyladenosine (m6A)
- m6A effectors
- gene regulation
- physiologic and pathogenic processes
- crop improvement
INTRODUCTION
Gene expression, which intricately intertwines with physiological and pathogenic processes, is precisely modulated through mRNA metabolism and protein synthesis throughout the course of development. Recently, RNA chemical modifications across diverse RNA subtypes have emerged as a regulatory mechanism orchestrating both cellular transcriptomes and proteomes in various physiological processes. Over 170 different types of postsynthesized modifications have been identified in cellular RNA, notably within ribosomal RNA (rRNA), transfer RNA (tRNA), messenger RNA (mRNA), and noncoding RNA (ncRNA), bearing heavy modifications (Boccaletto et al. 2018). Among these, N6-methyladenosine (m6A) stands out as the most extensively characterized RNA modification (Dominissini et al. 2012). Much like DNA epigenetics and histone modifications, m6A methylation undergoes dynamic deposition, removal, and recognition by specific cellular components, respectively. These components play pivotal roles in modulating gene expression throughout development and in response to environmental stimuli (Frye et al. 2018).
In the realm of plant epitranscriptomics, m6A is recognized as the most widespread and extensively studied internal RNA modification found across diverse plant species. Herein, we highlight its functional mechanisms governing gene expression regulation and summarize recent studies regarding its critical roles in plant development and environmental stress response, as well as crop trial implications. Additionally, we underscore the persisting unanswered questions about the plant epitranscriptome as well as forthcoming challenges in crop improvement in relation to m6A epitranscriptomics.
Regulatory network of m6A effectors in plants
m6A cotranscriptionally incorporates to RNA polymerase II-transcribed transcripts, facilitated by an evolutionarily conserved multicomponent m6A methyltransferase complex (referred to as the “writers”) (Huang et al. 2019). In mammalian cells, m6A installation is charged by the highly conserved methyltransferase complex comprising methyltransferase like 3 (METTL3), methyltransferase like 14 (METTL14), Wilms’ tumor 1-associating protein (WTAP), VIRMA, CBLL1, RNA-binding motif protein 15/15B (RBM15/15B), and zinc finger CCCH-type containing 13 (ZC3H13) (Song et al. 2021a). Likewise, this complex in Arabidopsis comprises two core methyltransferases, mRNA adenosine methylase (MTA) and MTB (Zhong et al. 2008), along with accessory proteins like FKBP12 interacting protein 37KD (FIP37) (Shen et al. 2016), virilizer (VIR), and HAKAI (Fig. 1; Růžička et al. 2017). FIP37 and VIR play crucial roles in stabilizing MTA and MTB, serving as key subunits maintaining the molecular function of the m6A methyltransferase complex (Shen 2023). Recent findings propose HAKAI-interacting zinc finger protein, HIZ2, as a necessary component for m6A abundance, further contributing to the Arabidopsis m6A writer complex (Zhang et al. 2022a). Interestingly, FPA, akin to metazoan m6A writer components RBM15/RBM15B, interacts with m6A writers (Parker et al. 2021). However, its abnormal expression does not globally affect m6A levels, suggesting limited or locus-specific methylation fine-tuning by FPA. Additionally, another known m6A methyltransferase FIONA1, a human METTL16 ortholog, acts independently to modify a subset of transcripts and U6 small nuclear RNA (snRNA), contributing modestly to overall m6A levels (Sun et al. 2022; Wang et al. 2022; Xu et al. 2022). In rice, enhanced downy mildew 2-like (OsEDM2L) has been proposed as the third identified m6A methyltransferase, characterized by a highly conserved N6-adenine methyltransferase-like (MTL) domain. Decreased transcriptome-wide m6A levels in osedm2l mutants emphasize the critical role of OsEDM2L in shaping the m6A methylation landscape (Ma et al. 2021).
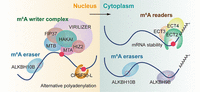
Regulation of m6A methylation for gene expression in Arabidopsis. Almost all mRNA m6A modification is catalyzed by a multiprotein complex containing MTA, MTB, FIP37, VIR, HAKAI, and HIZ2, and removed by the m6A demethylases ALKBH10B and ALKBH9B. YTH domain-containing proteins recognize m6A to modulate gene expression.
Although many types of methyltransferases have been identified, understanding how m6A writers achieve transcript- and site-specific methylation in response to developmental and environmental cues remains a challenge. To date, two predominant models have been proposed to explain the specificity of m6A methylation. One mechanism depends on the recruitment of the writers with RNA-binding proteins (RBPs) and other associated proteins. For instance, OsFIP37-associated protein 1 (OsFAP1) recruits the m6A writer OsFIP37 to add m6A on OsYUCCA3 transcripts during male meiosis in rice (Cheng et al. 2022). Under blue light, cryptochrome 2 (CRY2) undergoes liquid–liquid phase separation (LLPS), forming CRY2-nuclear bodies. It interacts with MTA, MTB, and FIP37, facilitating m6A addition on transcripts of core circadian clock genes (Wang et al. 2021). Conversely, photoexcited CRY2 co-condenses FIONA1 solely in the presence of suppressor of phytochrome A (SPA1). CRY2 and SPA1 work together to activate FIONA1's RNA methyltransferase activity, while CRY2 and FIONA1 (but not MTA) are crucial for light-induced methylation and translation of mRNAs regulating chlorophyll homeostasis (Jiang et al. 2023). The other model is the cotranscriptional m6A deposition. As methylation occurs cotranscriptionally, events during transcription probably guide the site-specific methylation coupled with histone modifications (Huang et al. 2019). In Arabidopsis, m6A sites are associated with H3K36me2 marks (Shim et al. 2020), suggesting potential epigenetic-mediated m6A deposition in plants.
Unlike m6A methyltransferases that require multiple complex proteins to function, m6A demethylases (referred to as the “erasers”) appear to act independently in removing m6A modifications. Currently, only two Fe(II)/α-KG-dependent m6A demethylases, fat mass and obesity-associated protein (FTO) and ALKBH5, have been characterized in mammals (Jia et al. 2011; Zheng et al. 2013). In Arabidopsis, while FTO lacks plant orthologs, there are 13 ALKBH-homologous proteins, of which ALKBH9A/9B/9C/10A/10B share homology with ALKBH5 (Duan et al. 2017). Herein, ALKBH9B and ALKBH10B have been well-characterized as m6A demethylases in Arabidopsis, conferring m6A demethylation in their specific targets (Fig. 1; Duan et al. 2017; Martínez-Pérez et al. 2017). ALKBH9B possesses m6A demethylation activity and specifically removes m6A from the genomic RNA of Alfalfa mosaic virus (AMV) (Martínez-Pérez et al. 2017). Recent study has revealed that ALKBH9B selectively demethylates a heat-activated retrotransposon, ONSEN, and thereby releases it from stress granule (SG) confinement, allowing for its mobilization (Fan et al. 2023). Moreover, ALKBH10B is abundant in most organs and loss-function of ALKBH10B leads to a significant m6A level increase (Duan et al. 2017). In addition, a study in tomatoes found that SlALKBH2 is an RNA m6A demethylase that localizes to the endoplasmic reticulum (ER) (Zhou et al. 2019). Interestingly, although both ALKBH9B and ALKBH10B are bona fide Arabidopsis m6A methyltransferase, the depletion of either ALKBH9B or ALKBH10B resulted in hypomethylated m6A peaks predominantly enriched in the CDS region and 3′ UTR, respectively (Duan et al. 2017; Tang et al. 2022). Furthermore, ALKBH9B localizes in the cytoplasm and SG during heat stress, while ALKBH10B is a nucleo-cytoplasmic protein. This suggests that these proteins function independently in demethylation, targeting distinct m6A sites within different cellular compartments.
The precise regulation of m6A on RNA metabolism relies on direct and indirect recognition by its binding proteins (referred to as the “readers”). The YT521-B homology (YTH) domain was discovered to directly engage with the m6A molecule through an “aromatic pocket,” where two or three tryptophan residues spatially enclose the m6A site. This ∼150-amino acid YTH domain is highly conserved across various eukaryotes, spanning humans, fruit flies, yeast, and plants (Song et al. 2021a). In mammals, five YTH proteins (YTHDF1–3, YTHDC1–2) are known to tightly bind m6A-modified transcripts in both the cytoplasm and nucleus (Wang et al. 2014, 2015; Xiao et al. 2016; Shi et al. 2017; Wojtas et al. 2017). Notably, plant genomes encode more YTH domain proteins compared to other eukaryotes: 13 in Arabidopsis, 12 in rice, 9 in tomato, and 39 in wheat. Furthermore, these genes exhibit diverse expression patterns across different developmental stages and in response to various stimuli. This suggests that these YTH domain-containing proteins redundantly and precisely target specific transcripts, playing distinct regulatory roles.
In Arabidopsis, 12 out of 13 proteins contain the YTH domain, primarily from the EVOLUTIONARILY CONSERVED CARBOXY-TERMINAL REGION (ECT) family, labeled as ECT1–12. The remaining one is a predominant isoform of the polyadenylation factor CLEAVAGE AND POLYADENYLATION SPECIFICITY FACTOR30 (CPSF30), known as CPSF30-L. Among these, ECT2 has been first characterized as m6A reader and shown to interact with ECT3 and ECT4 in the cytoplasm, enhancing m6A-binding capabilities and exhibiting genetically redundant functions (Arribas-Hernández et al. 2018; Wei et al. 2018). In line with their spatial interaction, ECT3 shares most of its target genes with ECT2, and ECT2/ECT3/ECT4 collectively stabilizes their targeted m6A-modified mRNAs, yet does not influence translation, thereby post-transcriptionally regulating mRNA abundance (Arribas-Hernández et al. 2021; Song et al. 2023). In mechanism, ECT2 directly interacts with the poly(A) binding proteins, PAB2 and PAB4, contributing to the stabilization of m6A-modified mRNAs (Song et al. 2023). A recent study showed that ECT1 undertakes LLPS to form cytosolic biomolecular condensates, such as processing bodies (PBs) and SG, in response to salicylic acid (SA) or bacterial pathogens, therefore sequestering SA-induced m6A modification-prone mRNAs and facilitating their decay (Lee et al. 2023). Additionally, CPSF30-L acts as a nuclear m6A reader protein and undergoes LLPS to form liquid-like nuclear bodies, where CPSF30-L recognizes the m6A-modified far-upstream element (FUE) alone or in combination with the near-upstream element (NUE) signal AAUAAA to control poly(A) site choice and transcriptional readthrough (Fig. 1; Song et al. 2021b). The diverse subcellular locations and regulatory mechanisms of m6A-binding proteins are fundamental in fine-tuning various aspects of RNA processing and metabolism, highlighting the precision and complexity of m6A regulation. Apart from the model plant Arabidopsis, counterparts such as MhYTP2 in apple (Guo et al. 2022), SlYTH2 in tomato (Ao et al. 2023), and YTH03/05/10 in rice (Cai et al. 2023) have been identified, all of which play pivotal roles in RNA metabolism.
Beyond the YTH family, additional putative m6A readers (IGF2BP1–3) have been discovered (Huang et al. 2018), directly interacting with methylated transcripts through their RNA-binding domain. FLOWERING LOCUS K (FLK), an RBP containing K-homology (KH) motifs, has emerged as a novel mRNA m6A reader (Amara et al. 2023). It directly binds to the m6A site in the 3′ UTR of FLC transcripts, thereby repressing FLC levels by reducing stability and affecting splicing. However, the existence of indirect m6A reader proteins in plants and the mechanisms through which these distinct and redundant proteins specifically and accurately recognize m6A, modulating RNA expression, require further exploration for a comprehensive understanding of the m6A epitranscriptome in plants.
Biological functions of m6A in diverse plant physiologic and pathogenic processes
Having established the complex processes of m6A methylation and their significant role within plant cells, it is crucial to extend our perspective to its specific impacts on plant growth, development, and response to environmental stresses, as demonstrated by the characterization of a collection of mutants in RNA modification effectors. Thus, it is beneficial to consider how these molecular mechanisms translate into concrete biological outcomes in plants.
Given that loss-function of core m6A components, such as MTA, MTB, and FIP37, lead to embryo lethality, strategies using the ABI3 and LEC1 promoters, which strongly drive the expression of downstream genes during the embryonic state, have been devised to express MTA or FIP37 (Bodi et al. 2012; Shen et al. 2016). This approach enables bypassing of the embryonic development stage that is disrupted by null mutations, thereby enhancing our understanding and study of m6A-mediated functions. In accordance with this strategy, an ABI3:MTA/mta mutant was generated with significant m6A reduction, displaying severe developmental defects in apical dominance and organ definition (Bodi et al. 2012). Likewise, the fip37-4/LEC1:FIP37 mutant shows a considerable over-proliferation of shoot apical meristem (SAM), with reduced m6A methylation resulting in the destabilization of WUSCHEL (WUS) and SHOOT MERISTEMLESS (STM) transcripts (Shen et al. 2016). Additionally, impaired expression of other m6A writers also results in defective root growth, suggesting that m6A methylation is critical for developmental decisions (Růžička et al. 2017; Zhang et al. 2022a). m6A reader protein, ECT2, has been reported to control leaf and trichome morphogenesis, in redundancy with ECT3 and ECT4 (Arribas-Hernández et al. 2018; Wei et al. 2018). Moreover, more defective phenotypes were observed in ect2/ect3/ect4, encompassing root growth, stem length, delayed flower formation, aberrant flower morphology, and a decreased cell division ratio (Arribas-Hernández et al. 2020). Intriguingly, both ECT2 overexpressed and ect2/ect3/ect4 plants exhibit abnormal siliques and aborted seeds (Arribas-Hernández et al. 2020; Wu et al. 2020), suggesting that a precise quantity of m6A readers is fundamental for fertilization in Arabidopsis (Fig. 2).

Biological functions of m6A in plant development. m6A plays critical functions in plant development, including embryogenesis, stem cell fate determination, root development, leaf morphology, floral transition, sporogenesis, fruit ripening, photosynthesis, and circadian rhythm. Figure was created with Biorender.com.
Floral transition is important for plants, with several regulatory proteins of m6A providing key insights for this process. For instance, the absence of ALKBH10B and FIONA1 leads to late and early flowering, respectively (Duan et al. 2017; Wang et al. 2022; Xu et al. 2022). Removing CPSF30-L causes a delayed flowering time by regulating the poly(A) site choice of the SUPPRESSOR OF OVEREXPRESSION OF CO 1 (SOC1) (Song et al. 2021b). The newly discovered m6A reader FLK binds and regulates FLC to regulate floral transition in an m6A-dependent manner (Amara et al. 2023). As for the reproduction process in plants, in Oryza sativa, the loss of function in OsFIP37 results in a lack of viable pollen, a reduced panicle, and smaller anthers (Zhang et al. 2019). A separate study identified OsEDM2L as crucial for anther and pollen development, regulating tapetal programmed cell death (PCD) in an m6A-dependent manner (Ma et al. 2021). Fruit ripening is a meticulously coordinated process governed by thousands of genes, responsible for the gradual softening or hardening of fruit layers. In tomatoes, the DNA demethylase SlDML2, crucial for ripening, governs the transcription of m6A demethylase SlALKBH2. This, in turn, associates with SlDML2 transcripts, facilitating the degradation of SlDML2 mRNA relying on m6A (Zhou et al. 2019). In strawberries, the m6A level and distribution are gradually varying during fruit maturation. Plants subjected to MTA and MTB RNA interference showed a lag in fruit maturation (Fig. 2; Zhou et al. 2021).
The sensing of light in the environment is important to plants; it can be crucial for competition and survival. It has been noticed that loss of FIP37 does not significantly impact photosynthesis under normal conditions, but it is vital for photosynthesis efficiency and chloroplast function during cold acclimation (Govindan et al. 2022), while genetic deactivation of VIR significantly diminishes photosynthetic activity when exposed to high light (Zhang et al. 2022b). Recent study has shown that the removal of FIONA1 leads to an extension of the hypocotyl in both long-day and short-day conditions (Wang et al. 2022). Additionally, the CRY2/SPA1 complex undergoes blue light-induced LLPS to condense FIONA1 in the mediation of photomorphogenesis and photosynthesis in plants (Jiang et al. 2023). It has also been discovered that CRY photoreceptors govern the blue-light regulation of m6A modification in over a tenth of the mRNAs, particularly those influenced by the circadian rhythm. In ABI3:MTA/mta and cry1cry2 mutants, the circadian cycle was significantly extended in continuous white light, as well as the m6A level and abundance of CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) mRNA decreased under blue light (Fig. 2; Wang et al. 2021).
The m6A modification plays a regulatory role in various cellular responses and alterations in m6A writers trigger significant changes in the expression of genes that respond to stresses. In Arabidopsis, under salt stress, MTA, MTB, and VIR can regulate the m6A levels on the transcripts of genes that regulate salt stress response, affecting the stability of these transcripts in order to maintain homeostasis under stress (Anderson et al. 2018; Hu et al. 2021). Moreover, FIONA1-mediated m6A methylation regulates the production of reactive oxygen species (ROS) and influences the transcription levels of various salt stress-responsive genes, which in turn facilitate the homeostasis of plants in salt stress (Cai et al. 2024). Likewise, m6A demethylases, ALKBH9B/10B, also regulate the stability of the transcripts of several key genes to modulate the response through m6A modification under not only salt stress but also abscisic acid (ABA) (Shoaib et al. 2021; Tang et al. 2021, 2022). Recent research on cold stress has revealed that MTA can regulate the freezing tolerance of Arabidopsis as ABI3:MTA/mta mutant displays cold hypersensitivity determined by primary root growth, biomass, and ROS accumulation (Govindan et al. 2022). In line with m6A writers and erasers, m6A reader proteins are required for diverse abiotic stress response. The m6A-binding protein CPSF30-L affects the length of the 3′ UTR of certain transcripts through m6A recognition, participating in the ABA and nitrogen signaling pathway (Hou et al. 2021; Song et al. 2021b). ECT2/ECT3/ECT4 could redundantly affect seedling germination under high-concentration ABA (Fig. 3; Song et al. 2023). In addition to Arabidopsis, the m6A-mediated abiotic stress responses have been thoroughly investigated in a variety of crops and trees, including cadmium stress in barley (Su et al. 2022); salt stress in sweet sorghum and sugar beet (Zheng et al. 2021; Cui et al. 2022); lead stress in soybean (Zhang et al. 2023); low-temperature stress in tomato (Yang et al. 2021); drought and oxidative stress in apple (Hou et al. 2022); and drought tolerance in poplar (Lu et al. 2020; Gao et al. 2022). However, the regulatory mechanism of m6A in the stress response of crop species remains largely unexplored, making it crucial to gain insights into how m6A methylation intricately contributes to the complex regulatory network not only in development but also in stress response.
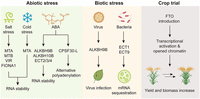
Biological functions of m6A in stress responses and crop trial. m6A plays critical functions in abiotic and biotic stress, including salt, cold, ABA, virus, bacteria stresses, and crop yield improvement. Figure was created with Biorender.com.
Apart from abiotic stresses, plant diseases are the foremost threat to sustainable agriculture and present the substantial risk for production. The RNAs of plant viruses, AMV and Cucumber mosaic virus (CMV), are known to contain m6A methylation (Martínez-Pérez et al. 2017). An analysis revealed that the demethylation activity of Arabidopsis ALKBH9B influences the infectivity of AMV, but not CMV (Martínez-Pérez et al. 2017). This mechanism is not exclusive to Arabidopsis but is also observed in other plant species. For instance, the down-regulation of the Nicotiana benthamiana AlkB homologs from the plant-specific ALKBH9 clade resulted in a significant decrease in the accumulation of Plum pox virus (PPV) and Potato virus Y (PVY) (Yue et al. 2022a). It is interesting that the RNA-dependent RNA polymerase (RdRp) that Pepino mosaic virus (PepMV) encodes has the capacity to interact with SlHAKAl, leading to the degradation process (He et al. 2023a). This discovery suggests a new tactic in the continuous battle between plants and viruses. Excluding virus infection, the plant diseases caused by bacteria and fungi could also be severe. In Arabidopsis, global depletion of m6A results in increased resistance to bacterial and fungal infections (Prall et al. 2023). m6A readers ECT1 and ECT9 form condensates triggered by SA, and the overexpression of ECT1 promotes bacterial multiplication in Arabidopsis (Fig. 3; Lee et al. 2023; Wang et al. 2023). As for crops, MTA1 plays a crucial role during the infection process of Magnaporthe oryzae in rice (Ren et al. 2022). MhYTP2, from Chinese crab apple, has been found to significantly enhance resistance to powdery mildew caused by Podosphaera leucotricha (Guo et al. 2022). Additionally, the role of m6A methylation extends beyond infection processes. Research has also shown that m6A also plays a significant part during the conidial germination in Fusarium graminearum. This discovery lays the groundwork for future investigations into m6A modification sites in filamentous fungi and provides a deeper understanding of these pathogens and how they interact with their hosts (Kim et al. 2024).
Advances in using m6A of crop improvement
With the rapid advancements in understanding the physiological effects of m6A on plant development and stress resistance, a crucial consideration arises: How can m6A be effectively applied to enhance crop yield? A groundbreaking study demonstrated that the yield of two crops (rice and potatoes) could be increased by nearly 50% through the heterologous expression of FTO (Yu et al. 2021), an m6A demethylase expressed in mammals. The primary discovery of this research is the capability to elevate the grain yield of rice over threefold under greenhouse conditions. Field trials further revealed that the introduction of FTO into rice and potatoes boosted both yield and biomass approximately by half. Additional findings from the study indicate that FTO stimulates root meristem cell proliferation and tiller bud formation, enhances photosynthetic efficiency, and drought resistance, all without inducing any negative side effects (Fig. 3). Moreover, the underlying molecular mechanisms between m6A and heterosis, a phenomenon extensively used to increase productivity in crops, remain largely elusive. Recently, researchers conducted a comprehensive analysis of RNA m6A methylation and gene expression changes in two Arabidopsis ecotypes and their reciprocal hybrids (Xu et al. 2021b). This study provides an overall pattern of inheritance of mRNA m6A modifications from parents to hybrids. Another research yielded comprehensive transcriptome profiles of mRNA abundance, m6A methylation, and translational efficiency from the maize F1 hybrid B73×Mo17, alongside its two parental lines (Luo et al. 2021). Although the findings showed that the overall abundance and distribution of m6A remained consistent, it revealed a substantial number of genes exhibiting alterations at the m6A level. This research substantially enriches our comprehension of the molecular mechanisms driving heterosis in crops.
OUTLOOK
Recent studies have increasingly highlighted the significance of the epitranscriptomic mark, m6A, as a critical regulator of gene expression, demonstrating its fundamental importance in various aspects of plant biology and the development of crop yield. Despite the progress made in this field, our comprehension of the plant epitranscriptome remains in its early stages. There remain numerous unanswered questions concerning the precise regulatory mechanism of m6A, its role in environmental signal transduction, and its potential for crop resilience and yield improvement, all of which require further exploration.
Current research on the m6A regulatory mechanism in plants remains inadequate, and understanding its interplay with other epitransgenic modifications, such as histone modification and DNA methylation, remains elusive. In Arabidopsis, m6A has mainly been detected in polymerase II-transcribed transcripts, and modulates gene expression via alternative polyadenylation, mRNA stability, miRNA processing, and alternative splicing (Shen et al. 2023). However, in mammalian cells, m6A has been found to be abundant in various types of ncRNAs including rRNA, snRNA, snoRNA, lncRNA, circRNA, as well as repeat RNAs, exerting significant influence on gene expression and biological processes (Song et al. 2021a). This suggests that m6A methylation harbors diverse and unexplored functional roles within plants. Moreover, transcriptional regulation involves intricate dynamics, where histone modifications and DNA methylation are pivotal components. Recent studies have unveiled roles: METTL3-mediated m6A methylation of retrotransposons coordinating with H3K9me3 histone modifications to guide heterochromatin formation, modulating retrotransposon repression, and safeguarding embryonic stem (ES) cell identity (Xu et al. 2021a). FTO mediates m6A demethylation of LINE1 RNA, regulating LINE1 RNA abundance and local chromatin state during mouse oocyte and embryonic development (Wei et al. 2022). Additionally, METTL3-mediated m6A methylation recruits TET1 cotranscriptionally to demethylate DNA 5mC through an m6A reader, FXR1 (Deng et al. 2022). Notably, FTO expression in rice mediates m6A demethylation, enhancing chromatin accessibility and transcriptional activation (Yu et al. 2021), indicating that m6A may play a role in orchestrating transcriptional regulation through its interaction with histone and DNA methylations. These findings collectively contribute to advancing and broadening our understanding of the plant epitranscriptome.
In response to environmental changes, signaling pathways undergo rewiring of gene expression programs, in which the m6A level is dynamically altered and recognized by its specific reader proteins to fine-tune downstream gene expression. Under various abiotic stresses, such as salt and ABA stimuli, there is a notable alteration in the expression level of m6A regulators (Hu et al. 2021; Tang et al. 2021). Considering that transcription factors (TFs) are key regulators of intrinsic and extrinsic processes, the transcriptional axis involving TF, m6A, and gene expression enabling m6A regulators “monitor” external stress to maintain plant homeostasis during stress responses. Besides, the intricate mechanisms by which m6A writers, erasers, and readers confer external or internal stress signaling transduction are not yet fully understood. Given the dependency of cellular signaling processes on post-translational protein modifications (PTMs) for the rapid reprogramming of individual protein functions, it is imperative to explore whether these components harbor PTMs and how these PTMs tune m6A-mediated gene expression regulation. Hence, further research in this area is essential to gain a comprehensive understanding of how m6A perceives environmental changes and stimuli in plants.
The multifaceted roles of m6A in the developmental and stress adaptation processes of varied plant species suggest that modifying the epitranscriptome holds potential in enhancing crop disease resilience. Commonly, short-term disease resistance is achieved via chemical treatments like fungicides or pesticides, despite their propensity to cause environmental pollution. It is been observed that pathogen infections can alter the global m6A levels in plants (Yue et al. 2022b), indicating that chemicals activating or deactivating m6A writers and erasers could bolster short-term defense against diseases caused by viruses or other pathogens. Biocontrol agents, beneficial organisms adept at controlling plant diseases, offer another approach. Exploring m6A modifications in pathogens could uncover new aspects of epigenetic regulation in both plants and pathogens, which could help us better understand the molecular mechanisms behind infections, plant immune responses, and the development of pathogen resistance. Notably, current research has shown that alterations in m6A modification in viral or host RNA can control viral infection (He et al. 2023b), while the precise regulatory mechanism remains largely unexplored. Future investigations should consider various factors like cell type, viral and fungi or bacteria strains, and infection duration, and use optimal sequencing technologies to systematically examine specific mechanisms of m6A in the life cycle not only in plants but also pathogens. In addition to short-term plant disease resistance improvements, long-term disease control often relies on strategies like breeding disease-resistant plant varieties or managing environmental conditions to reduce disease risk. Importantly, CRISPR/Cas13 systems have been developed and function as valuable tools for RNA targeting and editing in plant studies, predominantly used to enhance viral resistance in plants (Aman et al. 2018). Given the close association of m6A modification with biological processes in plants, integrating m6A effectors with (d)Cas13 could be a novel strategy for the precise regulation of target functional genes. For example, dCas13 in conjunction with m6A writers or erasers could regulate the m6A level of specific transcripts. Further, m6A readers could trigger downstream regulatory functions of certain genes. Considering its current uses and future potential, the combination of m6A and the CRISPR/Cas13 system could develop a potent platform for RNA targeting in plants, paving the way for innovative agricultural applications.
During the plant growth process, plants are subject to the trade-off between plant growth and disease resistance, an essential component in determining their survival strategies, growth trends, and evolutionary paths. Generally, plants have a fixed amount of resources that they can allocate to various functions, such as growth, reproduction, and defense against pests and diseases. When conditions are favorable, plants may prioritize growth, investing more resources into the production of new tissues and expanding their size. However, this often comes at the cost of reduced investment in defense mechanisms, leaving the plant more vulnerable to be attacked by pathogens and diseases. Conversely, when plants are under threat from pests or diseases, they may shift their resource distribution toward defense (Figueroa-Macías et al. 2021). Thus, how to balance and utilize resources at the maximal level is essential for crop improvement. Recent research has revealed that IDEAL PLANT ARCHITECTURE 1 (IPA1) enhances both crop yield and disease resistance by maintaining a balance between growth and immunity (Wang et al. 2018). Another study has shown that the wheat Tamlo-R32 mutant maintains normal crop growth and yields, while providing strong resistance to powdery mildew (Li et al. 2022). Additionally, using epitranscriptome editing provides a promising strategy to fine-tune the trade-off process through precise gene expression regulation. Indeed, introducing the human m6A demethylase in rice and potato not only increases biomass but also enhances drought tolerance (Yu et al. 2021), highlighting the potential of combining epigenetic modifications to develop crop varieties that not only have high yields, but also possess robust and long-lasting disease resistance. Overall, mechanistic understanding of epitranscriptome marks, precise identification of m6A-associated signaling transduction responding to environmental stress, exploration of efficient molecular inhibitor or activator, development of effective plant RNA editors, as well as application of epitranscriptomic marks in crops are crucial components of epitranscriptome advancing, promising to unlock its potential in crop improvement.
ACKNOWLEDGMENTS
This work was supported by the National Natural Science Foundation of China (nos. 22225704, 21820102008, and 92053109) and the National Basic Research Program of China (2019YFA0802201).
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079951.124.
-
Freely available online through the RNA Open Access option.
This article, published in RNA, is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








