
Fungi of the order Mucorales express a “sealing-only” tRNA ligase
- Department of Microbiology and Molecular Genetics, UT MD Anderson Cancer Center UTHealth Houston Graduate School of Biomedical Sciences, University of Texas Health Science Center, Houston, Texas 77030, USA
- Corresponding author: ambro.van.hoof{at}uth.tmc.edu
-
Handling editor: Eric Phizicky
Abstract
Some eukaryotic pre-tRNAs contain an intron that is removed by a dedicated set of enzymes. Intron-containing pre-tRNAs are cleaved by tRNA splicing endonuclease, followed by ligation of the two exons and release of the intron. Fungi use a “heal and seal” pathway that requires three distinct catalytic domains of the tRNA ligase enzyme, Trl1. In contrast, humans use a “direct ligation” pathway carried out by RTCB, an enzyme completely unrelated to Trl1. Because of these mechanistic differences, Trl1 has been proposed as a promising drug target for fungal infections. To validate Trl1 as a broad-spectrum drug target, we show that fungi from three different phyla contain Trl1 orthologs with all three domains. This includes the major invasive human fungal pathogens, and these proteins can each functionally replace yeast Trl1. In contrast, species from the order Mucorales, including the pathogens Rhizopus arrhizus and Mucor circinelloides, have an atypical Trl1 that contains the sealing domain but lacks both healing domains. Although these species contain fewer tRNA introns than other pathogenic fungi, they still require splicing to decode three of the 61 sense codons. These sealing-only Trl1 orthologs can functionally complement defects in the corresponding domain of yeast Trl1 and use a conserved catalytic lysine residue. We conclude that Mucorales use a sealing-only enzyme together with unidentified nonorthologous healing enzymes for their heal and seal pathway. This implies that drugs that target the sealing activity are more likely to be broader-spectrum antifungals than drugs that target the healing domains.
Keywords
INTRODUCTION
In all eukaryotes, a subset of tRNA precursors contain introns that are removed by a specialized set of tRNA splicing enzymes (Popow et al. 2012; Phizicky and Hopper 2023). In contrast to spliceosomal splicing of mRNAs and other noncoding RNAs, tRNA splicing is a protein-mediated process initiated by cleavage of the pre-tRNAs by the tRNA splicing endonuclease (TSEN) complex (Peebles et al. 1979; Hayne et al. 2023). TSEN generates a 5′ exon that ends with a 2′3′-cyclic phosphate and a 3′ exon with a 5′ hydroxyl in a catalytic mechanism that is similar to that used by RNase A (Peebles et al. 1983). This initial step of tRNA splicing appears conserved in all eukaryotes, but two distinct mechanisms are used for the subsequent ligation step of two tRNA halves (5′ and 3′ exons) (Greer et al. 1983; Phizicky et al. 1986).
Most Metazoa and some protozoa use a “direct ligation” mechanism to join the tRNA exons, which is carried out by RTCB enzymes (Fig. 1A). RTCB uses a catalytic histidine residue to transfer GMP onto the 3′ end of the 5′ exon, generating an activated exon that can then be ligated to the 3′ exon (Chakravarty et al. 2012; Englert et al. 2012; Desai et al. 2013; Jacewicz et al. 2022). RTCB has been implicated in tRNA splicing in humans, Caenorhabditis elegans, and Drosophila melanogaster, and homologs of RTCB are encoded in most metazoan genomes (Popow et al. 2011; Kosmaczewski et al. 2014; Lu et al. 2014; Nandy et al. 2017).
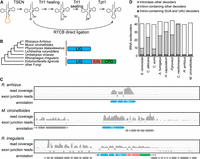
The Mucorales contain an atypical Trl1 ortholog with only the “sealing” domain. (A) Schematic of the tRNA splicing pathway in yeast and human. (B) Domain architecture of Saccharomyces cerevisiae Trl1 and orthologs from Mucorales (Rhizopus arrhizus, Mucor circinelloides, Phycomyces blakesleeanus, Lichtheimia corymbifera), the closely related Umbelopsidales Umbelopsis vinacea, and the more distantly related Rhizophagus irregularis and Entomortierella lignicola. (LIG) ligase domain, (KIN) kinase domain, and (CPD) cyclic phosphodiesterase domain. The species tree is from Zhao et al. (2022). (C) The tRNA LIG of M. circinelloides and R. arrhizus express as sealing-only single domain Trl1s, whereas R. irregularis Trl1 contains three domains. Shown is RNA-seq read coverage of the TRL1 orthologs of M. circinelloides, R. arrhizus (both Mucorales), and the related R. irregularis tRNA LIG. Colors reflect domains as in A. (D) The Mucorales contain a minimal set of intron-containing tRNAs. The number of intronless isodecoders (white), intron-containing isodecoders (gray), and intron-containing Tyr GUA (decoding UAC and UAU codons) and Ile UAU (decoding AUA codons) (dark gray) isodecoders in the genomes of major pathogenic fungi is shown.
In contrast, most fungi and plants and some other protozoa use a “heal and seal” mechanism (Fig. 1A) carried out by the multifunctional enzyme Trl1 to join tRNA exons (Konarska et al. 1981; Greer et al. 1983; Phizicky et al. 1986; Abelson et al. 1998; Sawaya et al. 2003; Lopes et al. 2015; Phizicky and Hopper 2023). The heal and seal mechanism involves three distinct steps that are carried out by three different catalytic domains of Trl1. In one “healing” reaction, the CPD domain converts the 2′3′-cyclic phosphate of the 5′ exon to a 3′-OH, 2′ PO4 end. In a second “healing” reaction, the KIN domain converts the 5′ hydroxyl of the 3′ exon to a 5′ phosphate. Finally, in the “sealing” reaction, the LIG domain uses a catalytic lysine residue to transfer AMP onto the 3′ exon, which activates it for ligation to the 5′ exon (Wang et al. 2006; Banerjee et al. 2019). The initial ligation product contains a 2′ phosphate at the exon junction, which is removed by tRNA 2′-phosphotransferase (Tpt1) (Spinelli et al. 1997; Steiger et al. 2001). Thus, both the catalytic residue (His vs. Lys) that carries out activation, as well as the exon (5′ vs. 3′) that is activated, differ between the direct ligation and heal and seal pathways.
Although bacteria do not have a similar tRNA splicing pathway, some bacteria contain RTCB homologs that instead function in tRNA repair (Supplemental Fig. 1), including the Escherichia coli homolog RtcB (Tanaka and Shuman 2011; Tanaka et al. 2011; Chen and Wolin 2023). The bacteriophage T4 encodes an alternative tRNA repair pathway that uses a heal and seal strategy that is slightly different and negates the need for Tpt1 (Supplemental Fig. 1; Amitsur et al. 1987; Shuman 2023). Although the main function of yeast Trl1 under laboratory conditions is in tRNA ligation (Cherry et al. 2018), it also functions in tRNA repair under natural conditions where yeast grows in the presence of other species that secrete tRNA-cleaving toxins (Nandakumar et al. 2008). Strikingly, these distinct pathways are interchangeable and the yeast TRL1 gene can be replaced by either E. coli RtcB or T4 PNK and T4 Rnl1 (Schwer et al. 2004; Tanaka et al. 2011).
Both RTCB and Trl1 have an additional function in the splicing of a single mRNA during the unfolded protein response (UPR) (Sidrauski et al. 1996; Gonzalez et al. 1999; Mori 2009). During endoplasmic reticulum (ER) stress, the endonuclease Ire1 cleaves the S. cerevisiae (hereafter “yeast”) HAC1 and human XBP1 mRNAs. Similar to TSEN, Ire1 also generates a 5′ exon that ends with a 2′3′-cyclic phosphate and a 3′ exon with a 5′ hydroxyl, and these exons are ligated by RTCB and Trl1 in Metazoa and Fungi, respectively, using the same mechanism used for tRNA ligation.
The fungal tRNA exon ligation mechanism has been proposed as an attractive target for antifungal drug development for several reasons (Wang and Shuman 2005; Tanaka et al. 2011; Chakravarty et al. 2012; Remus et al. 2016): (i) tRNA splicing is essential in all eukaryotes. Without it, eukaryotes cannot generate a complete set of tRNAs and cannot translate a subset of codons. (ii) Fungi are relatively closely related to humans, and there are very few fundamental differences in their biochemical pathways. As a result, there are very few genes that are essential in fungi but that do not have a close homolog in the human genome (Braun et al. 2005; Liu et al. 2006). (iii) The biochemical mechanism of tRNA exon ligation by fungal Trl1 is distinct from the metazoan enzyme RTCB. (iv) All three domains of Trl1 are required for its function in tRNA ligation and cell survival in the model fungus S. cerevisiae (Sawaya et al. 2003; Wang and Shuman 2005; Wang et al. 2006). Therefore, a Trl1 inhibitor that targets any of the three essential domains is a potential antifungal drug candidate.
Novel antifungal drugs are urgently needed. There are only three classes of approved antifungal drugs for invasive disease, and resistance to these drugs limits their use. The CDC considers Candida auris as one of six urgent threats of antimicrobial-resistant pathogens, other drug-resistant Candida species as a serious threat, and azole-resistant Aspergillus as a watch list threat (https://www.cdc.gov/drugresistance/biggest-threats.html). In the USA, fungal diseases are responsible for seven billion dollars in annual healthcare costs (Benedict et al. 2019) and 3% of all hospitalized patients receive an antifungal drug (Vallabhaneni et al. 2018). The WHO issued a report recently calling for increased research into 19 fungi that represent the greatest threat to human health (https://www.who.int/news/item/25-10-2022-who-releases-first-ever-list-of-health-threatening-fungi). Fungal infections alarmingly have become one of the major global health problems among immunocompromised individuals. Among several types of fungal infections, invasive and pulmonary fungal diseases such as candidiasis, aspergillosis, cryptococcal meningitis, pneumocystis pneumonia, histoplasmosis, and mucormycosis are leading causes of morbidity and mortality worldwide (Garber 2001; von Lilienfeld-Toal et al. 2019; Kainz et al. 2020; Reddy et al. 2022). In addition, patients with existing health conditions such as HIV, cancer, and SARS-CoV-2 appear to have a high risk of fungal coinfections (Bodey et al. 1992; Limper et al. 2017; Soltani et al. 2022). Strikingly, a sharp increase in deaths caused by fungal infection was observed during the coronavirus disease pandemic (Gold et al. 2023). Widespread antifungal resistance against commonly used drugs, particularly for C. auris, worsens the current threat of fungal infection (Pristov and Ghannoum 2019; Miramón et al. 2023). For infections caused by Mucorales, drugs are generally insufficient, and radical surgical debridement is an important but disfiguring treatment.
The fungi that cause invasive lethal infections in humans are spread throughout the fungal kingdom, including members of the phyla Ascomycota (including Candida albicans, C. auris, Aspergillus fumigatus, Histoplasma capsulatum, and Pneumocystis jirovecii), Basidiomycota (Cryptococcus neoformans), and Mucoromycota (R. arrhizus a.k.a. R. oryzae). In previous studies, Trl1 homologs from the human ascomycete pathogens C. albicans, A. fumigatus, and Coccidioides immitis have been shown to complement an S. cerevisiae trl1Δ strain (Remus et al. 2016), supporting that Trl1 function is conserved and a good candidate for being targeted by antifungal drugs.
In this study, we identified functional homologs of ScTrl1 in a broader range of human pathogenic fungi species that represent diverse evolutionary relatedness. The Trl1 homologs that have been shown to complement trl1Δ are all from other Ascomycete fungi. Here, we have identified “typical” Trl1 orthologs in Ascomycota, Basidiomycota, and Mucoromycota pathogens that have all three heal and seal domains and are capable of complementing trl1Δ, supporting that Trl1-targeting drugs might be broad-spectrum antifungals. Interestingly, we identified an “atypical” Trl1 ortholog in the order Mucorales (R. arrhizus and M. circinelloides). These Trl1 orthologs contain a Trl1 ligase domain, but lack CPD and KIN domains. These Mucorales Trl1 “sealing-only” orthologs can functionally complement defects in the ligase domain of ScTrl1 but not trl1Δ. Using orthologous mutation in the conserved nucleotide-binding motif, we demonstrate that the sealing-only Trl1 of Mucorales is mechanistically similar to ScTrl1. Finally, the sealing-only Trl1 of Mucorales can perform the sealing step of HAC1 mRNA splicing in yeast, indicating that both the known functions of Trl1 are retained. This suggests that drugs that target the ligase domain are likely to be broader-spectrum antifungals than those targeting the CPD or KIN domains. These findings imply that the Mucorales use KIN and CPD enzymes in a heal and seal pathway that are not orthologs to Trl1.
RESULTS
Most pathogenic fungi contain a typical Trl1 ortholog, except the Mucorales
We sought to identify the orthologs of S. cerevisiae tRNA ligase (ScTrl1) in a diverse range of pathogenic fungi species, with a focus on pathogens that cause invasive diseases with high morbidity and mortality. Two primary criteria were considered for identifying ScTrl1 orthologs: (a) the sequence-level similarities among the fungal TRL1 genes with ScTRL1, and (b) the presence of three domains in the fungal TRL1 genes. First, we compared the human pathogens that belong to the phylum Ascomycota. The Ascomycota are subdivided into three subphyla that each include important human pathogens. Among the subphylum Saccharomycotina, the genomes of two important human pathogens, C. albicans and C. auris (causative agents of candidiasis), encode “typical” Trl1 orthologs with all three catalytic domains and an overall sequence similarity to ScTrl1 of 59% and 55%, respectively. Two representative fungi of the subphylum Pezizomycotina, A. fumigatus (causative agent of aspergillosis) and H. capsulatum (causative agent of histoplasmosis), also contain ScTrl1 orthologs with overall sequence similarity of 54% and 55%, respectively. Finally, the subphylum Taphrinomycotina includes one significant human pathogen, P. jirovecii, and its genome encodes a typical Trl1 ortholog with 52% similarity to ScTrl1. Thus, the genomes of all human pathogens within the ascomycetes encode a typical Trl1 ortholog. Within the phylum Basidiomycota, C. neoformans is the most important human pathogen and its genome encodes a ScTrl1 ortholog with 46% similarity to ScTrl1.
Strikingly, we could not identify a typical Trl1 ortholog in the genome of Rhizopus species, the most important pathogen within the Mucoromycota (e.g., R. arrhizus), or in closely related species (e.g., M. circinelloides). Instead, the Trl1 orthologs in these species were much smaller and appeared to consist of only a ligase domain (Fig. 1B). These ligase domains were highly similar to that of ScTrl1 (55% and 49%). Similar sealing-only Trl1 homologs were encoded in other genomes of the order Mucorales and the related Umbelopsidales, and all of them show sequence similarity with the ligase domain of ScTrl1. In contrast, in other Mucoromycota, we readily identified typical Trl1 orthologs that contain all three domains (e.g., R. irregularis, overall 52% similar to ScTrl1). This suggests that a typical Trl1 ortholog was present in the common ancestor of the Mucoromycota, Basidiomycota, and Ascomycota, but was shortened after the divergence of the Mucorales and Umbelopsidales from the other fungi. Despite extensive searches, we were unable to identify any genes in the Mucorales that were orthologous to the CPD and KIN domains of Trl1.
To confirm that the Trl1 orthologs from Rhizopus and Mucor were truncated, we analyzed their gene structure by RNA-seq. A reanalysis of publicly available RNA-seq data confirms that transcripts of R. arrhizus and M. circinelloides TRL1 orthologs encode only the ligase domain. In M. circinelloides, the ligase domain is encoded in a single exon that is not spliced with any other exon. In R. arrhizus, the ligase domain is encoded by three exons that are spliced together, but not to any other exons. In contrast, the four exons that encode the ligase domain of R. irregularis Trl1 are clearly spliced together with additional exons that encode the KIN and CPD domains (Fig. 1C). Overall, our data suggest that most human fungal pathogens express a typical Trl1 ortholog, but the Mucorales instead express a “sealing-only Trl1.”
The Mucorales contain a small number of tRNA introns and other components of a heal and seal tRNA splicing pathway
We investigated several possible explanations for the absence of a typical Trl1 from the Mucorales. First, we considered the possibility that a lack of tRNA introns in the Mucorales means they do not need a typical Trl1. Although all eukaryotes are thought to have tRNA introns, the proportion of tRNA genes that contain introns varies widely. We observed the same pattern when focusing on human fungal pathogens (Fig. 1D; Supplemental Table 1). Eukaryotic genomes generally encode 42–50 distinct tRNAs (isodecoders) that each decode one to four different codons. Each of these isodecoders can be encoded by a single tRNA gene or by multiple tRNA genes. If an isodecoder is encoded by multiple genes, typically either all of the genes for that isodecoder contain an intron, or none of them do. Among the fungal pathogens analyzed, we saw similar patterns, with C. neoformans containing the largest number of tRNA introns (92% of its tRNA genes). In contrast, R. arrhizus and M. circinelloides contain a relatively small number of intron-containing tRNAs (3% and 4% of its tRNA genes, respectively). Further analysis of isodecoder tRNAs in fungal pathogens revealed that all of the genes for two specific tRNAs had an intron: one tyrosine tRNA (which decodes both UAU and UAC) and the isoleucine tRNA that decodes AUA codons (but not AUU and AUC Ile codons). Thus, R. arrhizus and M. circinelloides require tRNA splicing to decode three of the 61 sense codons. Whereas the genes for these same tRNAs contained introns in all fungal human pathogens, there was a wide variation in the number of other intron-containing tRNA isodecoders and genes (Fig. 1D). These observations rule out the possibility that the Mucorales do not need a typical Trl1 because they might lack tRNA introns.
The TSEN complex mediates the endonuclease cleavage of the intron-containing tRNA (Fig. 1A) and comprises two catalytic subunits (Sen2 and Sen34) and two structural subunits (Sen15 and Sen54). The two catalytic subunits are more highly conserved at the sequence level, and we readily identified orthologs of both in the genomes of all the major invasive fungal pathogens, including R. arrhizus and M. circinelloides (Supplemental Table 2). The presence of both tRNA introns and a TSEN in the Mucorales strongly suggests that they must contain a fully functional tRNA splicing pathway.
One possible explanation for the absence of a three-domain Trl1 is that Mucorales acquired an RTCB-like LIG. We therefore searched their genomes but failed to detect any RTCB homologs. A third ligase (C12orf29) has recently been identified in humans and suggested to possibly function in tRNA splicing (Yuan et al. 2023), but we could not find any orthologs in the Mucorales genomes.
If the Mucorales indeed use the truncated Trl1 in a heal and seal tRNA splicing pathway, they should have homologs of Tpt1, the enzyme that removes the 2′ phosphate left after the sealing step (Fig. 1A). Indeed, the genomes of the Mucorales each include an obvious Tpt1 ortholog (Supplemental Table 2). Whereas a Tpt1 is required for the fungal heal and seal pathway, Tpt1 enzymes are also found in bacteria, human, and archaea where they perform other (partially unknown) functions (Spinelli et al. 1998; Ohira et al. 2022).
The most likely explanation of these bioinformatic analyses is that the Mucorales have functionally replaced the “healing” domains of Trl1 with a nonorthologous CPD and KIN. Similar replacements have previously been artificially achieved in S. cerevisiae (see Discussion), but to the best of our knowledge have not been reported naturally.
The sequence and predicted structure of sealing-only Trl1s of Mucorales suggest they are functional RNA ligases
The shorter tRNA ligase candidates of R. arrhizus and M. circinelloides contain 376 and 372 amino acids, respectively. Previous studies have identified that the amino-terminal ligase domain of Trl1 orthologs contains six conserved peptide motifs (I, Ia, III, IIIa, IV, and V) that are found in the nucleotide-binding pocket (Wang and Shuman 2005). An amino acid sequence alignment reveals that these conserved motifs are present in R. arrhizus and M. circinelloides and therefore suggests that these proteins encode functional RNA ligases (Supplemental Fig. 2).
To gain insights into the structural features of Mucorales Trl1, we generated predicted structures of R. arrhizus and M. circinelloides Trl1 using both AlphaFold2 (Varadi et al. 2022) and Phyre tools (Fig. 2A–E; Kelley et al. 2015). Notably, although these tools use different approaches to predict structure, they give very similar results with high confidence. The predicted structures obtained from the analyses indicate that both Mucorales Trl1s share high structural similarities with the ligase domain of the available X-ray structure of the Ascomycete fungus Chaetomium thermophilum Trl1 (protein data bank [PDB]: 6N0V, 6N0T, and 6N67) (Fig. 2A–C; Banerjee et al. 2019; Peschek and Walter 2019). The secondary structures predicted by AlphaFold2 of the amino-terminal subdomain of Mucorales Trl1 were superimposed with C. thermophilum Trl1 using the Matchmaker tool of Chimera (Fig. 2D,E). The overall architecture and the catalytic pocket, including the conserved motifs, are conserved. The only notable difference in the amino-terminal subdomains is a small amino-terminal extension (∼30 amino acid residues) on the C. thermophilum protein that is distant from the catalytic pocket and unlikely to alter the function. The carboxy-terminal subdomain of Mucorales Trl1 also shows high structural similarity to the C. thermophilum X-ray structure. In C. thermophilum, this domain consists of four α-helices, and all four are conserved in the AlphaFold predicted Mucorales structures. The only notable difference between the AlphaFold2 predictions and the C. thermophilum structures is that AlphaFold2 predicts two short antiparallel β strands inserted between the second and third α-helix of the carboxy-terminal subdomain. These strands are distant from the catalytic pocket and unlikely to alter the function of these proteins. Furthermore, AlphaFold2 predicts that the amino- and carboxy-terminal subdomains are oriented very similar to those in C. thermophilum.
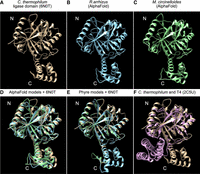
The predicted structures of Mucorales Trl1 resemble the ligase domain of fungal tRNA ligase. (A) X-ray crystal structure of the ligase domain of the ascomycete fungus C. thermophilum (6N0T). (B) The AlphaFold predicted the structure of R. arrhizus sealing-only Trl1. (C) The AlphaFold predicted the structure of M. circinelloides sealing-only Trl1. (D) Superimposition of X-ray crystal and AlphaFold predicted structures from A to C. Colors are as in A–C. For R. arrhizus RMSD is 0.913 Å over 247 Cα atoms; for M. circinelloides it is 0.901 Å over 244 Cα atoms. (E) Superimposition of X-ray crystal and Phyre predicted structures. Colors are as in A–C. (F) Superimposition of X-ray crystal structures of Trl1 ligase from C. thermophilum (6N0T) and T4 RNA ligase (2C5U; plum colored). The amino- and carboxy-terminal subdomains are indicated in each panel.
To more objectively compare our AlphaFold2 predictions of M. circinelloides Trl1 and R. arrhizus Trl1 to known structures, we performed a DALI search of the PDB. For M. circinelloides and R. arrhizus Trl1, the top DALI recovered structure was the X-ray structure of the ligase domain of the C. thermophilum (PDB: 6N0T and 6N0V) with a z-score of 41.8 and 41.3, respectively (Banerjee et al. 2019). The second most related structure from the DALI search was the X-ray structure of T4 RNA ligase (Rnl1) (PDB: 2C5U) (El Omari et al. 2006). A key distinction between Trl1 and T4 Rnl1 is that Trl1 uses a substrate with a 2′ PO4 3′ OH, whereas T4 Rnl1 uses a substrate with 2′ OH 3′ OH (Supplemental Fig. 1; Schwer et al. 2004; Banerjee et al. 2019). This biochemical difference is mediated by a carboxy-terminal extension on the Trl1 ligase domain (Banerjee et al. 2019). The 6N0T structure identifies key residues that recognize the 2′ PO4 (N150, H227, R334, and R337), and each of these residues is conserved in the Rhizopus and Mucor proteins and positioned in the same place in the AlphaFold2 models. The R334 and R337 residues are part of a YRxxR motif in the carboxy-terminal subdomain, and the T4 Rnl1 carboxy-terminal subdomain does not contain a similar motif. The T4 Rnl1 subdomain is also positioned differently relative to the amino-terminal subdomain (Fig. 2F). Taken together, the predicted structures of Mucorales Trl1 suggest these proteins are active RNA ligases that require a 2′ PO4.
The sealing-only Trl1s of Mucorales functionally complement the ScTrl1 ligase domain
In order to determine whether the Trl1 orthologs from invasive pathogenic fungi are functional, we expressed Trl1 orthologs in an S. cerevisiae trl1Δ strain. A set of plasmid shuffle assays allowed for the selection of the yeast cells that express fungal Trl1 orthologs and have lost the wild-type ScTrl1 plasmids (Fig. 3A). Each of the tested three domains containing “typical fungal Trl1s” from pathogenic fungi complemented trl1Δ. Specifically, replacing Trl1 with the Trl1 orthologs from the closely related C. albicans and C. auris resulted in growth that resembled wild type. Complementation with the ortholog of the more distantly related Ascomycota A. fumigatus, H. capsulatum, or P. jirovecii also resulted in growth. This complementation by orthologs from other Ascomycota confirms and extends previous reports (Remus et al. 2016). Remarkably, the full-length Trl1 orthologs from the Basidiomycota C. neoformans and the Mucoromycota R. irregularis can also complement trl1Δ. Some of the trl1Δ complemented strains grew at a reduced rate, whereas others grew at near-normal rate. Unexpectedly, the expression of some of the Trl1 orthologs caused some dominant-negative growth defects in the presence of yeast Trl1 that were not well correlated with divergence time (control plates in Fig. 3A). These likely are consequences of the heterologous expression strategy and may not reflect meaningful differences between the Trl1 orthologs. Overall, these results indicate that all of these typical three-domain fungal Trl1s can perform tRNA ligation in budding yeast. In contrast, as expected the truncated Trl1 orthologs from R. arrhizus and M. circinelloides failed to complement trl1Δ and behaved similar to an empty vector control.
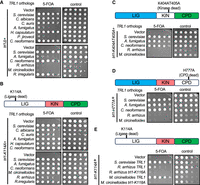
Complementation of Trl1 defects by orthologs from fungal pathogens. (A) The Trl1 orthologs of representative pathogenic fungi complement trl1Δ, except the sealing-only Trl1s of Mucorales. The indicated Trl1 orthologs were expressed in S. cerevisiae trl1Δ [TRL1, URA3] strain background and subjected to serial dilution growth assay on media containing 5-FOA (selecting against the TRL1 plasmid) or control media. (B–D) The Mucorales Trl1 can complement a defect in the ligase domain of ScTrl1 (B) but is unable to complement the defects in the kinase (C) or CPD (D) domain of ScTrl1. The indicated Trl1 orthologs were expressed in a strain that contains a URA3 plasmid with a wild-type TRL1 gene and another plasmid with either the sealing-defective trl1-K114A (B), the kinase-defective trl1-K404AT405A (C), or the CPD-defective trl1-H777A (D) subjected to serial dilution growth assay on media containing 5-FOA (selecting against the TRL1 plasmid) or control media. (E) A conserved lysine in the active site is critical in the Mucorales Trl1s. Amino acid changes orthologous to ScTrl1-K114A were introduced in Mucorales Trl1 and subjected to plasmid shuffle assay in the sealing-defective ScTrl1 strain as in B.
We postulated that because the identified Mucorales Trl1s only contain the ligase domain, they could only complement the exon ligation step (sealing step) but not the healing steps of the fungal type tRNA splicing pathway. A conserved lysine residue in position 114 of ScTrl1 is required for the function of the ligase domain. Amino acid change of the K114 residue to alanine (K114A) exhibits defects in the sealing function of Trl1 and is lethal (Sawaya et al. 2003). We therefore performed a plasmid shuffle assay to test whether Trl1 orthologs of pathogenic fungi can functionally complement a ligase domain defect (trl1-K114A). As expected, all of the fungal Trl1 orthologs that complement trl1Δ also complemented trl1-K114A. Importantly, the sealing-only Trl1s from Mucorales can complement this ligase-dead ScTrl1 (Fig. 3B). As expected, we confirmed that the Mucorales Trl1s cannot perform the “healing” reactions of the tRNA splicing pathway in budding yeast: they do not complement mutations in either the ScTrl1 KIN domain (trl1-K404AT405A) or the CPD domain (trl1-H777A) (Fig. 3C,D).
We examined whether the Mucorales Trl1s use the same catalytic lysine as ScTrl1. Sequence alignment identified the corresponding conserved lysine residues as K116 for R. arrhizus and K118 for M. circinelloides. A plasmid shuffle experiment indicates that changing these lysine residues to alanine in either of the Mucorales proteins disrupted the ability to complement the defect in the ScTrl1 ligase domain (Fig. 3E). Overall, these data indicate that Mucorales Trl1s are functional tRNA ligases and are able to perform the tRNA sealing step but not the tRNA healing steps.
Mucorales Trl1 can perform HAC1 mRNA splicing during the unfolded protein response
As discussed in the Introduction, in addition to tRNA splicing, Trl1 takes part in the nonconventional HAC1 mRNA splicing mechanism during the UPR that is initiated by Ire1. The UPR plays a critical role in the stress tolerance and virulence of pathogenic fungi (Richie et al. 2009; Cheon et al. 2011; Askew 2014; Krishnan and Askew 2014). During infection, fungal pathogens encounter several biotic and abiotic stresses in the host. Thus, an increased protein folding capacity in the ER via UPR signaling is required to overcome the host stresses associated with the increased pathogenicity of the fungal pathogens.
The Mucorales genomes contain an obvious ortholog for Ire1, but Hac1 orthologs are poorly conserved in sequence and difficult to detect across large evolutionary distances. Nevertheless, we identified a likely Hac1 ortholog in R. arrhizus (hypothetical protein G6F37_009984) and RNA-seq analysis identified an unannotated noncanonical intron (Fig. 4A).
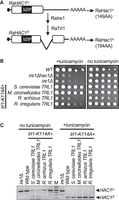
Mucorales sealing-only Trl1s can perform HAC1 splicing during the UPR. (A) R. arrhizus appears to have an Ire-dependent intron in HAC1 mRNA (encoding the 149 amino acid hypothetical protein G6F37_009984). Sequence similarity to Hac1 is highest in the indicated bZIP domain. Depicted is the unspliced (RaHAC1u) and spliced (RaHAC1s) mRNA structure based on aligning RNA-seq data (SRR1013749) to the genome with RNA STAR, and the predicted function of Ire1 and sealing-only Trl1 in generating spliced HAC1 mRNA. (B) The M. circinelloides, R. arrhizus, and R. irregularis Trl1 were expressed in S. cerevisiae trl1Δ [trl1-K114A] [TRL1, URA3] strain background and subjected to plasmid shuffle assay in the presence or absence of 0.2 µg/mL tunicamycin. The wild-type, ire1Δhac1Δ, and ire1Δ strains were used as a control. (C) RT-PCR of HAC1 indicates that upon UPR induction, the Mucorales Trl1 gives rise to spliced HAC1 mRNA (HAC1S). The HAC1 splicing defective ire1Δ strain was used as a control, which only produced the unspliced HAC1 mRNA (HAC1U).
To test whether R. arrhizus and M. circinelloides Trl1 are able to perform HAC1 mRNA splicing in budding yeast, we repeated the plasmid shuffle assay in the sealing-defective ScTrl1 strain (trl1-K114A) without or with 0.2 µg/mL tunicamycin. Tunicamycin causes unfolded proteins to accumulate in the ER; thus, the UPR is required for growth in the presence of 0.2 µg/mL tunicamycin. Indeed, ire1Δ and ire1Δhac1Δ control strains failed to grow in the presence of tunicamycin. Importantly, the strains that express Trl1 from the Mucorales and a sealing-dead ScTrl1 grew in the presence of tunicamycin and thus complemented the UPR defect (Fig. 4B), although the M. circinelloides ortholog performed better than the R. arrhizus ortholog. As a control, the three-domain Trl1 from the Mucoromycota R. irregularis also complemented the UPR defect.
To confirm that growth on tunicamycin reflected the ability to splice HAC1, we assayed HAC1 mRNA splicing directly by RT-PCR with primers that span the intron (Fig. 4C). When the cells were grown without UPR stress, the major RT-PCR product was the size expected for unspliced HAC1 mRNA. RT-PCR of the strains that express Mucoromycota Trl1s in addition to the ScTrl1 sealing-dead mutant resulted in a major product of 247 bp after UPR induction, reflecting the removal of the HAC1 intron. The UPR defective ire1Δ strain used as a control produced only unspliced HAC1 mRNA. Whereas the yeast strain expressing M. circinelloides Trl1 grew much better than the strain expressing R. arrizhus Trl1, the RT-PCR assay detected only spliced HAC1 mRNA in both strains. These heterologous expression assays were performed with different exposure times and doses of tunicamycin and are unlikely to reflect biological differences in the UPR of the two species. Taken together, these results indicate that R. arrhizus and M. circinelloides Trl1s are able to carry out the sealing step of HAC1 splicing during UPR stress when expressed in S. cerevisiae.
DISCUSSION
In this study, we show that the tRNA ligase (Trl1) orthologs from a diverse range of clinically relevant fungal pathogens are functionally conserved and can replace the Trl1 enzyme in S. cerevisiae. These fungal species contain a subset of tRNA genes that require splicing to produce functional tRNA molecules. This indicates that tRNA splicing is essential in all invasive fungal human pathogens. There is an urgent need for novel antifungal drugs. There are very few enzymes that are essential across the fungal kingdom, but are absent in humans. The heal and seal pathway is a prominent exception and thus an attractive target for antifungal drugs.
Pathogenic fungi including A. fumigatus and C. neoformans utilize the UPR to fine-tune protein folding capacity in the ER, which is linked to virulence, membrane homeostasis, cell wall homeostasis, and antifungal drug resistance (Richie et al. 2009; Cheon et al. 2011; Askew 2014; Krishnan and Askew 2014; Weichert et al. 2020). Thus, targeting the fungal UPR has been suggested as a potential alternative strategy for antifungal development. The advantage of Trl1 as a drug target is that a Trl1 inhibitor would inhibit both tRNA splicing and the UPR in pathogenic fungi.
We proposed that, unlike other representative fungi, Mucorales (e.g., R. arrhizus and M. circinelloides) have separate sealing and healing enzymes to carry out tRNA exon ligation, with the healing enzymes being unidentified. The phylogeny of species with sealing-only Trl1 enzymes (Fig. 1B) suggests the loss of healing domains occurred before the Mucorales diverged from the Umbelopsidales (an estimated 389 million years ago according to timetree.org) but after they diverged from the Rhizophagus ancestor (∼769 million years ago). Our identified Trl1 orthologs in Mucorales are functional ligases that can carry out the sealing reactions in both known Trl1 functions: tRNA and HAC1 mRNA splicing. Furthermore, this activity is conserved almost 400 million years after the loss of the healing domains, strongly suggesting that the sealing activity performs a critical function for the Mucorales. However, the lack of the Trl1 KIN and CPD domains in these orthologs suggests that the sealing-only enzymes likely act with some other polynucleotide KIN and phosphodiesterase that carry out the healing steps of tRNA and HAC1 splicing. Although the presence of a Tpt1 ortholog is consistent with this proposal, Tpt1 homologs are also found in species that do not carry out heal and seal tRNA splicing (Spinelli et al. 1998) and thus the presence of a Tpt1 is not strong evidence for a heal and seal pathway. Alternative models for tRNA splicing in Mucorales seem less likely, but cannot be completely excluded. One alternative possibility is that the Mucorales use an unknown RNA ligase (i.e., not detectably similar to Trl1, RTCB, or C12orf29). This possibility would imply that the conserved sealing-only enzyme has some other unknown function. Another alternative explanation is that the endonuclease that cleaves out tRNA introns in Mucorales generates ends that can be directly ligated by the sealing activity (i.e., 2′ and 5′ monophosphates). We consider it more likely that some other phosphodiesterase or KIN activity has taken over the Trl1 healing function, and there are several candidate enzymes encoded in the Mucorales genomes that are not orthologous to Trl1.
There is experimental precedence of replacing Trl1 healing activities with nonorthologous enzymes: The healing domains of Trl1 in yeast can be replaced by the distantly related mammalian RNA 5′-KIN CLP1 and 2′,3′ cyclic nucleotide phosphodiesterase CNP (Ramirez et al. 2008; Schwer et al. 2008). Furthermore, the Mucorales have a Clp1 ortholog, which is a candidate for one of the healing reactions. Because the Mucorales proteins can seal the 2′ PO4 and 5′ PO4 ends generated by yeast healing domains, the unknown KIN and CPD likely also generate 2′ PO4 and 5′ PO4 ends.
We note that the Rhizopus and Mucor that express a sealing-only Trl1 also have only two tRNA isoacceptors that contain an intron: tRNA-Tyr-GUA and tRNA-Ile-UAU tRNA genes. tRNAscan-SE (Lowe and Eddy 1997) confidently detects these same two tRNA introns in the other three species from Figure 1B with sealing-only Trl1s: P. blakesleeanus, L. corymbifera, and U. vinacea. However, it also detects with lower confidence a possible tRNA-Tyr-AUA with an intron for U. vinacea. Although it is tempting to speculate that the variant tRNA splicing pathway in Mucorales may be mechanistically linked to its low tRNA intron count, this may not be the case. For E. lignicola, which appears to have a full-length Trl1 (Fig. 1B), tRNAscan-SE also detects only tRNA genes tRNA-Tyr-GUA and tRNA-Ile-UAU with introns with high confidence but additional intron-containing tRNAs Lys-CUU and Asn-AUU with low confidence. Thus, the tRNA repertoire of E. lignicola resembles that of the Mucorales and U. vinacea, but its Trl1 is typical.
Our study supports the possibility of targeting the tRNA exon ligation mechanism for antifungal development. The ligase domain of fungal Trl1 is conserved in all of the important invasive human pathogens, including the Mucorales. Thus, a drug that inhibits the function of the Trl1 ligase domain should be a broad-spectrum antifungal against these fungal pathogens. In contrast, drugs that target the KIN and CPD domains are less likely to be effective against the Mucorales. However, an antifungal that targets Ascomycete and Basidiomycete Trl1s would still be useful, as these taxa include the most urgent fungal threats (https://www.cdc.gov/drugresistance/biggest-threats.html) and account for most of the cost of fungal infections (Benedict et al. 2019).
MATERIALS AND METHODS
Identification of fungal Trl1 genes
To identify Trl1 candidates in representative pathogenic fungi, extensive BLASTP and PSI-BLAST searches were performed starting with either ScTrl1 or the R. irregularis ortholog. The TRL1 genes were identified based on the similarity of the domain composition and the sequence. Orthologs of Sen2, Sen34, and Tpt1 were similarly identified with BLAST and PSI-BLAST. The search for alternative ligases used human RTCB, human C12ORF29, and E. coli RtcB protein sequences.
One explanation for failing to detect full-length Trl1 orthologs in the Mucorales by BLASTP and PSI-BLAST is that these algorithms only search annotated proteins. To rule out that we missed orthologs because they were unannotated, we also used TBLASTN to search genomes and assembled transcriptomes from the Mucorales in the WGS, HTGS, and TSA sections of GenBank.
To identify the correct exon junctions, publicly available RNA sequencing reads from R. arrhizus strain 99–892 (SRR1013749 and SRR1013747), M. circinelloides strain R7B (SRR1611143, SRR1611144, and SRR1611152), and R. irregularis (SRR14294959 and SRR14294960) (Dallaire et al. 2021) were aligned to the corresponding reference genome assemblies (GCA_024220505.1, GCA_001638945.1, and GCF_000439145.1, respectively) (Tisserant et al. 2013) using RNA STAR (Dobin et al. 2013).
tRNA annotations and their introns are from the genomic tRNA database (http://gtrnadb.ucsc.edu/index.html); for species not included in that database, tRNAs were identified using tRNAscan-SE 2.0, the same algorithm used for the genomic tRNA database (Chan and Lowe 2016; Chan et al. 2021).
Plasmids
Plasmids used are listed in Supplemental Table 3. Each of the TRL1 orthologs was cloned into p425GPD (Mumberg et al. 1995). For C. albicans and C. auris, the single exon genes were PCR amplified from genomic DNA (a kind gift from Michael Lorenz) and ligated with the vector. For the other species, we did not have ready access to genomic DNA or cDNA, and therefore the TRL1 ORF was codon optimized for expression in S. cerevisiae and synthesized by GenScript. The TRL1 gene was then cloned into the same vector by Gibson assembly. Expression of the P. jirovecii gene appeared to be toxic in E. coli and thus the plasmid for its expression was assembled directly in yeast by cotransforming a PCR product and a gapped plasmid.
The catalytic mutants for McirTrl1 (Mcir trl1-K118A) and RarrTrl1 (Rarr trl1-K116A) plasmids were created using the QuikChange Lightning Site-Directed Mutagenesis Kit following the manufacturer's instructions.
The catalytic mutants for ScTrl1 (trl1-K114A, trl1-H777A, and trl1-K404A-T405A) were generated by Gibson assembly into pRS413 (Sikorski and Hieter 1989; Sawaya et al. 2003).
All plasmids were sequence verified either by sequencing the whole plasmid using nanopore sequencing (Plasmidsaurus) or by sequencing the insert using Sanger sequencing (Genewiz).
Complementation of fungal Trl1s in trl1Δ yeast
The yeast strains used are listed in Supplemental Table 4. To identify whether the Trl1 of representative fungi can complement the loss of ScTrl1, the corresponding TRL1 expression plasmids of fungi were transformed into S. cerevisiae trl1Δ [TRL1, URA3] strain. This parent strain was created from the S. cerevisiae heterozygous knockout library by transforming a TRL1, URA3 plasmid (pAv1511) and sporulating the transformants. The resulting strain was matA, ura3-Δ0, leu2-Δ0, his3-Δ1, lys2-Δ0, trl1Δ::NEO, [TRL1,URA3] and unable to grow on 5-FOA-containing media.
The transformants for yeast cells containing TRL1 expression plasmids were selected in the SC media lacking leucine and uracil (SC-Leu-Ura, Sunrise Science). A plasmid shuffle experiment was performed, where the serially diluted yeast strains were spotted on the 5-FOA-containing medium that selects for yeast cells that have lost the WT ScTRL1, URA3 plasmid and contains only the TRL1 of corresponding fungi (Boeke et al. 1987). Growth was recorded after 4 d at 30°C.
To identify the ability of the fungal Trl1 to complement the defects in individual domains (ligase, kinase, CPD) of ScTrl1, the corresponding TRL1 plasmids were transformed into trl1Δ strains that contain sealing-dead, CPD-dead, or KIN-dead Trl1s in pRS413 (CEN, HIS3) plasmids in addition to the TRL1 gene on a CEN, URA3 plasmid. The transformants were selected on SC-Leu-His media (Sunrise Science), and a plasmid shuffle assay was performed as described above.
To identify the ability of the fungal Trl1 to complement the Trl1 function in the UPR, trl1Δ yeast cells containing a LEU2 plasmid expressing the TRL1 of other fungi, a HIS3 plasmid expressing Sc trl1-K114A and a URA3 plasmid expressing WT ScTRL1 were serially diluted and spotted on 5-FOA-containing media supplemented with or without 0.2 µg/mL tunicamycin (Sigma-Aldrich). Growth was recorded after 4 d at 30°C. The control ire1Δ strain was obtained from the knockout collection (Giaever et al. 2002). The control ire1Δhac1Δ strain (yAv4415) was generated by standard genetic crosses of single mutants obtained from the knockout collection. The hac1Δ::KANMX allele was converted to hac1Δ::NATMX using plasmid p4339 (obtained from Charles Boone).
RNA extraction and RT-PCR
Overnight cultures of yeast strains were diluted to ∼0.1 OD600 in YPD and grown to ∼0.6 OD600 at 30°C. UPR induction was carried out by adding 2.5 µg/mL tunicamycin for 2 h. The cells were harvested, and total RNA was extracted using the hot phenol method (He et al. 2008). The RNA samples were treated with DNase I (Invitrogen), and cDNA was synthesized with SuperScript II Reverse Transcriptase (Invitrogen). The HAC1 spliced and unspliced fragments were amplified with intron-spanning primers (ACCTGCCGTAGACAACAACAAT and AAAACCCACCAACAGCGATAAT) and analyzed on a 2% agarose gel (Cherry et al. 2018).
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
This work was funded by the National Institutes of Health (NIH) grant R35GM141710 to A.v.H. and a Dr. John J. Kopchick Research grant from the UT MD Anderson Cancer Center UTHealth Houston Graduate School of Biomedical Sciences to K.S.A. We thank Sean Johnson for help with AlphaFold2 predictions and Michelle Steiger, Mike Lorenz, Sean Johnson, and members of the van Hoof lab for insightful comments.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079957.124.
- Received November 16, 2023.
- Accepted January 20, 2024.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Khondakar Sayef Ahammed is the first author of this paper, “Fungi of the order Mucorales express a ‘sealing-only’ tRNA ligase.” Sayef is a PhD candidate in the laboratory of Professor Ambro van Hoof at the MD Anderson Cancer Center UTHealth Houston Graduate School of Biomedical Sciences. His research focuses on understanding the functional defects caused by disease-linked RNA exosome variants and validating tRNA ligase (Trl1) as a potential antifungal drug target.
What are the major results described in your paper and how do they impact this branch of the field?
The tRNA ligase Trl1 is required for the splicing of tRNAs and the HAC1 mRNA in plants and fungi and contains three different catalytic domains: two healing domains that generate the correct RNA ends that can then be ligated by the sealing domain. In this work, we have identified functional homologs of the “typical” three domains containing the tRNA ligase (Trl1) enzyme in a range of human pathogenic fungi. Interestingly, we found that fungi from the order Mucorales contain mini-tRNA ligases with only the sealing domain. We demonstrate that these Mucorales Trl1s can perform the sealing step of the tRNA and HAC1 splicing. This study opens up interesting questions including: Why do Mucorales choose an offbeat way for tRNA splicing, what are the alternative healing enzymes in Mucorales, and is this connected to the low tRNA intron content of Mucorales genomes?
What led you to study RNA or this aspect of RNA science?
During my childhood, I was totally convinced that there is a hidden world living with us that we can't see with our naked eyes. That curiosity led me to learn about microbial sciences during my undergraduate studies. Soon after I started my PhD journey, I realized that one of the ancient molecules on the earth, RNA, has a lot of untold stories. Every RNA molecule experiences a series of molecular events during its lifetime in the cell. How do the RNA molecules perform their assigned function inside the cell? What happens with the RNA when it finishes its job and is no longer required in the cell? These fundamental questions provoked me to learn about fascinating RNA-centric molecular machines and discover new stories about RNA.
During the course of these experiments, were there any surprising results or particular difficulties that altered your thinking and subsequent focus?
Mucormycosis causes very difficult-to-treat infections and yet there are very limited tools to study the Mucorales. I therefore had to rely on expressing Mucorales Trl1 in the very distantly related budding yeast. This worked surprisingly well on the first try.
What are some of the landmark moments that provoked your interest in science or your development as a scientist?
My first RNA meeting was in Boulder, Colorado. I came across several of the brightest minds in the RNA biology field, which significantly motivated my scientific mind. However, every little success and failure in my career has its role in developing my scientific curiosity and reorienting myself in the right direction.
If you were able to give one piece of advice to your younger self, what would that be?
I would advise my younger self, don't be afraid of making mistakes and explore new ideas! Over time, I learned that doing science is not at all a straightforward path. Stringent thinking mostly results in a predictable outcome, whereas the ability to think in an unconventional way may result in innovation. In fact, the greatest scientific discoveries that saved millions of lives are often accidental, including penicillin and cosmic radiation from the Big Bang.
Are there specific individuals or groups who have influenced your philosophy or approach to science?
My greatest source of motivation is Ambro! He has nurtured me uniquely, and I have slowly improved myself in several aspects of my scientific career. My advisory committee and the “van Hooligan” gang (my laboratory members) played an important role in developing my scientific philosophy.
What are your subsequent near- or long-term career plans?
“The woods are lovely, dark and deep,
But I have promises to keep, …
And miles to go before I sleep.”
—Robert Frost
Please read, “miles to go before I graduate!” There are several projects I hope to finish in the van Hoof lab. Ultimately, I would like to continue learning about RNA as a postdoc and an independent scientist in a research-intensive institute.








