
Ancestry of RNA/RNA interaction regions within segmented ribosomes
- Corresponding author: fox{at}uh.edu
Abstract
The ribosome is the universally conserved ribozyme that translates DNA coded instructions into proteins with the assistance of other RNA molecules, including transfer and messenger RNAs. Of particular interest is the segmentation phenomena, which is found in trypanosomatids and other protists. In these organisms, the large subunit ribosomal RNA is assembled from multiple smaller RNAs. This phenomenon posits several challenges to the folding and stabilization of such ribosomes to retain functionality and efficiency. In earlier studies, RNA/protein interactions were suggested to fully compensate for the fragmentation. Recently, several conserved RNA/RNA interaction regions were described in the cryo-EM structures of segmented ribosomes from trypanosomatids. These regions also seemed to aid in the folding and stabilization of such ribosomes, even before the ribosomal proteins start their association. In the present study, the existence of conserved RNA/RNA interaction regions shared between trypanosomatid and Euglena gracilis segmented ribosomes was confirmed, despite differences in segmentation patterns. Analysis of the crystallographic structures of unsegmented ribosomes from other Eukaryotes, Bacteria, and Archaea allowed us to estimate the relative age of highly conserved RNA/RNA interaction regions. These results strongly suggest that common interaction regions likely date far back into the ribosomes of the last common ancestor. Results also revealed that single hydrogen bonds are overwhelmingly facilitated by the 2′OH, a distinctive RNA feature. This supports the notion that RNA predates DNA and places some constraints on alternative nucleic acids proposals.
Keywords
INTRODUCTION
The modern ribosome is a ribonucleoprotein complex where coded protein synthesis takes place in all known living organisms. It has been recognized that at the core of the extant translation machinery resides a universally conserved ribozyme responsible for peptide bond formation (Cech 2000; Steitz and Moore 2003; Agmon et al. 2005; Bose et al. 2022; Rivas and Fox 2023). It is noteworthy that different reconstructions of the genetic content of the last common ancestor (LCA) strongly suggest that the translation machinery was essentially complete (Mushegian and Koonin 1996; Kyrpides et al. 1999; Harris et al. 2003; Koonin 2003; Mirkin et al. 2003; Delaye et al. 2005; Yang et al. 2005; Ouzounis et al. 2006; Ranea et al. 2006). Thus, it is likely that common features of the extant ribosome date from the LCA epoch and some even predate those days.
A typical modern ribosome is comprised of a small subunit (SSU) and a large subunit (LSU) (Steitz 2008; Fox 2010; Wilson and Doudna 2012). In prokaryotes, the SSU contains one 16S ribosomal RNA (rRNA) and ∼21 ribosomal Proteins (rProteins), while the LSU contains 5S and 23S rRNAs and ∼33 rProteins (Steitz 2008; Wilson and Doudna 2012). Eukaryotic ribosomes are typically formed by ∼80 rProteins and four larger rRNA chains, namely 5S, 5.8S, 18S, and 26S (or 28S) rRNA (Wilson and Doudna 2012). The 5.8S rRNA is homologous to the 5′-end of the prokaryotic 23S rRNA (Nazar 1980; Walker and Pace 1983).
The ribosomes of several protists contain rRNAs of unusual size and numbers. This rare phenomenon has also been described among some prokaryotic LSUs (Lessie 1965; Marrs and Kaplan 1970; Winkler 1979; Pronk and Sanderson 2001). The LSU of Crithidia oncopelti was first described to dissociate into two large rRNA components (Spencer and Cross 1976). It was subsequently realized that these two rRNAs together formed the equivalent of the 26S rRNA in Leishmania donovani (Leon et al. 1978). A similar case was reported in Leishmania tarentolae the same year (Simpson and Simpson 1978). The presence of small rRNAs was first reported in Crithidia fasciculata (Gray 1979). Shortly thereafter, the presence of additional low molecular weight rRNAs were reported in Trypanosoma brucei (Cordingley and Turner 1980). These observations were followed by reports that confirmed the presence of small rRNA components in C. fasciculata (Gray 1981; Schnare et al. 1983), and Trypanosoma cruzi (Hernández et al. 1983). Finally, it was concluded that the 26S rRNA from the LSU of several parasitic trypanosomatid protists has been replaced by two large rRNAs, and four small rRNAs (White et al. 1986; Campbell et al. 1987; Spencer et al. 1987). A far more extreme case of ribosome segmentation was identified in Euglena gracilis, a single-cell alga whose ribosome is assembled from 16 rRNAs. Three of these rRNAs are equivalent to the 18S, 5S, and 5.8S rRNAs found in other eukaryotic ribosomes. The 28S rRNA is then assembled from the 13 remaining rRNAs (Schnare and Gray 1990; Schnare et al. 1990).
The formation of an active segmented ribosome begins with the precursor rRNA being processed to release the pieces that will ultimately comprise the functional rRNA. First, the removal of internal transcribed spacers (ITSs) occurs, a process analogous to the maturation of the eukaryotic 5.8S rRNA (Spencer et al. 1987). In E. gracilis, the processing starts with the release of the 18S and the 5.8S rRNAs. Further cleavage splits the remaining rRNA into 13 pieces (Greenwood and Gray 1998). Although 14 ITSs are released during the precursor rRNA maturation process (Schnare et al. 1990), the liberation of mature rRNAs is not a sequential process. Instead, it follows a pattern of intermediate stages that most likely respond to folding requirements (Greenwood and Gray 1998). In the larger rRNA fragments, standard base-pairing seems to be involved in the assembly of the active subunit. This is not the case for several smaller fragments, which do not exhibit sites of standard base-pairing with the larger segments (Fig. 1; Matzov et al. 2020). The situation is similar in the trypanosomatid segmented ribosomes. In that case, the LSU is segmented into two large rRNAs, namely rRNA-α and rRNA-β, and four smaller segments, called srRNA-I, srRNA-II, srRNA-III, and srRNA-IV, respectively (Campbell et al. 1987; Hashem et al. 2013). It has been proposed that the srRNAs attach to the main rRNA scaffold by RNA/protein contacts (Liu et al. 2016) and long-range RNA/RNA interactions (Rivas and Fox 2022).
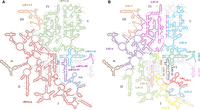
Secondary structure representation of the ribosomal segments from T. brucei and E. gracilis. Segments are drawn over the 28S rRNA secondary structure from S. cerevisiae, derived from crystallographic structure data (Petrov et al. 2014). Equivalences were defined using structural alignments as described in the Materials and Methods section. (A) Secondary structure shows the 5S (brown), the two large and four small segments that comprise the T. brucei LSU rRNA. 5.8S (blue) and rRNA-α (red) largely correspond to domains I and II. srRNA-I (orange) grossly corresponds to domain III. rRNA-β largely corresponds to domains IV and V. srRNA-II (cyan), srRNA-III (magenta), and srRNA-IV (purple) correspond to domain VI. (B) Secondary structure shows the 5S (brown) and the 14 segments that comprise the E. gracilis LSU rRNA. 5.8S/LSU-1 (red), LSU-2 (light blue), LSU-3 (yellow), and LSU-4 (rose) largely correspond to domain I. LSU-5 (light green) and LSU-6 (purple) grossly correspond to domain II. The last part of LSU-6 (purple), LSU-7 (orange), and the beginning of LSU-8 (magenta) largely correspond to domain III. Most of LSU-8 (magenta) corresponds to domain IV. The last part of LSU-8 (magenta) and LSU-9 (cyan) correspond to domain V. LSU-10 (green), LSU-11 (dark teal), LSU-12 (lavender), LSU-13 (dark blue), and LSU-14 (light brown) correspond to domain VI.
In addition, it was observed that trypanosomatid ribosomes have an uncommon abundance in expansion segments (Gao et al. 2005). This observation, as well as the involvement of trypanosome-specific proteins in ribosome biogenesis (Umaer et al. 2014), and the identification of trypanosome-specific RNA/protein interactions in T. cruzi (Umaer and Williams 2015), led to the proposal that assembly and stabilization of these segmented ribosomes was mainly a protein-dependent process involving “glue” proteins (Liu et al. 2016). On the other hand, the existence of several conserved long-range RNA/RNA interaction regions among the segmented ribosomes from T. brucei, L. donovani, and T. cruzi has been recently described (Rivas and Fox 2022). There, the authors suggested that these interactions also play an important role in the folding and stabilization of segmented ribosomes.
Here, we present a comparative analysis of the long-range RNA/RNA interaction regions from two srRNAs that are shared by the segmented ribosomes of T. brucei, L. donovani, T. cruzi, and E. gracilis. Together, these two segments represent the most conserved part of domain VI in the extant LSU. An analysis of equivalent interaction sites within unsegmented ribosomes from Saccharomyces cerevisiae, Escherichia coli, Thermus thermophilus, and Haloarcula marismortui was also possible. Implications of single hydrogen bonds (HBs) occurring among these interaction regions and the relevance of the 2′OH as an exclusive RNA feature are addressed.
RESULTS
Trypanosomatid and Euglena rRNA common segments
The trypanosomatid segmented ribosomes contain the 18S rRNA, the 5.8S rRNA, two large segments, rRNA-α and rRNA-β, and four small segments, srRNA-I, srRNA-II, srRNA-III, and srRNA-IV (Fig. 1A; Campbell et al. 1987; Hashem et al. 2013). E. gracilis segmented ribosomes contain the 18S rRNA, the 5.8S rRNA, also called LSU-1, and 13 rRNA fragments, LSU-2 to LSU-14 (Fig. 1B; Schnare and Gray 1990; Schnare et al. 1990; Matzov et al. 2020). Trypanosomatid small segments srRNA-I to srRNA-IV do not exhibit regions of canonical base-pairing that attaches them to the main rRNA scaffold (Rivas and Fox 2022). The same appears to be true for E. gracilis segments LSU-10 to LSU-14 (Fig. 1B).
Secondary and tertiary structure data have been used to show that segments srRNA-II, srRNA-III, and srRNA-IV constitute domain VI in extant trypanosomatid ribosomes (Rivas and Fox 2022). Likewise, segments LSU-10, LSU-11, LSU-12, LSU-13, and LSU-14 constitute domain VI in E. gracilis (Matzov et al. 2020). Here, structural superimpositions were used to confirm that structurally speaking srRNA-II is equivalent to LSU-10. Both comprise helixes 94, 95, 96, and 97 of the LSU secondary structure (Figs. 1, 2A,C). In the same manner, srRNA-IV is structurally equivalent to segments LSU-13 and LSU-14, which comprise helixes 99, 100, and 101 (Figs. 1, 2B,D). The remaining portion of domain IV, helix 98, is represented by srRNA-III in trypanosomatids and by LSU-11 and LSU-12 in E. gracilis (Fig. 1). Structural superimpositions of srRNA-III with LSU-11 and LSU-12 showed that folding patterns are essentially different for trypanosomatids and for E. gracilis. Furthermore, H98 has been recognized as a highly variable expansion segment commonly found in Eukaryotes and some Archaea (Penev et al. 2020). Whereas in several prokaryotic ribosomes it is almost nonexistent, this portion is completely absent from the common core of cytoplasmic rRNAs from all living creatures (Bernier et al. 2018). Therefore, it is not surprising that srRNA-III lacks a structural counterpart in other eukaryotic ribosomes. Hence, srRNA-III, LSU-11 and LSU-12 were discarded from the present comparative analysis.
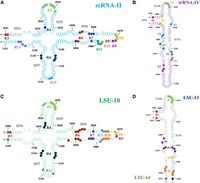
Secondary structure representations of segments srRNA-II and srRNA-IV from T. brucei and segments LSU-10, LSU-13, and LSU-14 from E. gracilis. These structures were created using the S. cerevisiae LSU secondary structure as a guide (Petrov et al. 2014); S. cerevisiae numbering system is used accordingly. Structural alignments of T. brucei and E. gracilis small segments over the S. cerevisiae LSU crystallographic structure guided their creation (PDBID 4V88) (Supplemental Tables S17, S18; Ben-Shem et al. 2011). Ribonucleotides within regions of interaction are marked with solid filled circles and a region number in the same color. Missing residues and regions were not mapped due to a lack of structural counterpart in S. cerevisiae. (A) Secondary structure of T. brucei segment srRNA-II (cyan), where interacting regions 3–13 have been mapped. These regions are a common feature among all ribosome structures (Table 1). (B) Secondary structure of T. brucei segment srRNA-IV (purple). Regions 1, 3, 5, 6, and 7 from srRNA-IV are a common feature among segmented and unsegmented ribosomal structures (Table 2). (C) Secondary structure of E. gracilis segment LSU-10 (green), where interacting regions 3–12 have been mapped. Regions 3–13 are a common feature among all ribosomes (Table 1). (D) Secondary structure of E. gracilis segments LSU-13 (dark blue) and LSU-14 (light brown). Interacting regions 1, 3, 5, 6, and 7 are a common feature among segmented and unsegmented ribosomes (Table 2).
Conserved regions of long-range RNA/RNA interaction in segmented ribosomes
There are 13 regions of long-range RNA/RNA interaction in the segment srRNA-II that have been proposed to aid in the stabilization and association with the main rRNA scaffold (Rivas and Fox 2022). Identification of equivalent regions in LSU-10 from E. gracilis was guided by structural alignment of corresponding segments. Regions 3–13 are persistent elements in the trypanosomatid and E. gracilis ribosomes regardless of their segmentation pattern (Table 1; Fig. 2). Likewise, seven regions of long-range RNA/RNA interaction were described for the segment srRNA-IV from trypanosomatids (Rivas and Fox 2022). Detection of equivalent regions was equally performed for segments LSU-13 and LSU-14 within E. gracilis. Regions 1, 3, and 5–7 are also persistent elements shared by both types of segmented ribosomes (Table 2; Fig. 2).
Distribution of the 13 long-range RNA/RNA interaction regions of srRNA-II (LSU-10 in E. gracilis) among segmented and unsegmented ribosomes
Distribution of the 13 long-range RNA/RNA interaction regions of srRNA-IV (LSU-13 and LSU-14 in E. gracilis) among segmented and unsegmented ribosomes
Variation within conservation
All previously described conserved interaction regions are the result of several long-range RNA/RNA interactions and single HBs converging into specific areas where rRNA elements, like helixes or loops, meet with other distant rRNA elements (Rivas and Fox 2022). Despite ribonucleotides being distant in primary sequence, they end near each other in the 3D space due to the folding process. Although their prevalence seems high among phylogenetically distant species, the number and nature of these interactions varies from organism to organism regardless of their segmentation nature.
For instance, in the segment srRNA-II from T. brucei, region 4 is made from six ribonucleotides that establish nine single HBs and two nonstandard base pairs with rRNA-α and rRNA-β. In L. donovani the equivalent region involves six ribonucleotides that establish eight single HBs, one A-minor motif, and one nonstandard base pair with rRNA-α and rRNA-β. This same region involves four ribonucleotides in T. cruzi that establish six single HBs with rRNA-β. In the case of E. gracilis, region 4 is represented by nine ribonucleotides from LSU-10 establishing nine single HBs, two A-minor motifs, one nonstandard base pair, and one base stacking element with LSU-9 (Table 3; Fig. 2).
Diversity of interaction types in region 4 from T. brucei, L. donovani, T. cruzi (srRNA-II), and E. gracilis (LSU-10)
Another example of such interaction heterogeneity can be seen in segment srRNA-IV from T. brucei, where region 3 is made from four ribonucleotides that establish three single HBs and two A-minor motifs with rRNA-β. In L. donovani the same region involves five ribonucleotides that establish five single HBs, one A-minor motif, and two base stacking elements with rRNA-β. This same region involves five ribonucleotides in T. cruzi that create six single HBs, one A-minor motif, and one large base stacking element with rRNA-β. In E. gracilis, region 3 is created by five ribonucleotides from LSU-13 that establish three single HBs, one A-minor motif, and one large base stacking element with LSU-8 and LSU-9 (Table 4; Fig. 2).
Diversity of interaction types in region 3 from T. brucei, L. donovani, T. cruzi (srRNA-IV), and E. gracilis (LSU-13 and LSU-14)
Dynamic conservation of RNA/RNA interaction regions within unsegmented ribosomes
It has been established that segments srRNA-II and srRNA-IV represent the most conserved part of domain VI in trypanosomatid ribosomes. The same is true for LSU-10, LSU-13, and LSU-14 in E. gracilis. They represent a portion of the common core of cytoplasmic rRNAs from all living creatures (Bernier et al. 2018). This observation encouraged the search for the long-range RNA/RNA interaction regions beyond segmented ribosomes and into ribosome crystallographic structures from Archaea, Bacteria, and other Eukaryotes. T. brucei segments and structures were previously identified as having most of the elements resolved in the cryo-EM structure, contrary to other trypanosomatid structures with poorly resolved areas. Therefore, it became the guiding structure for search purposes (Rivas and Fox 2022).
Structural alignments of T. brucei segments srRNA-II and srRNA-IV over X-ray crystallographic structures of unsegmented ribosomes from S. cerevisiae, E. coli, T. thermophilus, and H. marismortui guided the search for interaction regions and their characterization. Results reveal that most of the RNA/RNA interaction regions described previously for these two segments in trypanosomatids were also present in E. gracilis, and the unsegmented ribosomes from S. cerevisiae, E. coli, T. thermophilus, and H. marismortui (Tables 1, 2).
Interaction regions that were localized in the corresponding portions of domain VI from unsegmented ribosomes showed a significant amount of variation as described above for the segmented ribosomes. Take for instance region 4 in S. cerevisiae, which is made from six ribonucleotides that create five single HBs, two A-minor motifs, one nonstandard base pair, one ribose zipper, and one base stacking element. In E. coli this same region is made by 10 ribonucleotides that establish eight single HBs, two A-minor motifs, two nonstandard base pairs, and one ribose zipper. In T. thermophilus this region is made from 11 ribonucleotides that create 10 single HBs, three A-minor motifs, one nonstandard base pair, one ribose zipper, and one base stacking element. In H. marismortui, nine ribonucleotides create region 4, and they establish eight single HBs, two A-minor motifs, one nonstandard base pair, and one ribose zipper (Supplemental Tables S9, S11, S13, S15).
Another example of the diversity of interactions is seen in region 3 from srRNA-IV, which also shows significant variation. In S. cerevisiae, it is made from six ribonucleotides that create seven single HBs, one A-minor motif, and one large base stacking element. In E. coli, region 3 is made from six ribonucleotides that establish nine single HBs, one A-minor motif, and one large base stacking element. In T. thermophilus, this same region is made from six ribonucleotides that create eight single HBs, one A-minor motif, and one large base stacking element. In H. marismortui, four ribonucleotides create region 3. Together they establish three single HBs and two A-minor motifs (Supplemental Tables S10, S12, S14, S16).
The strong force of hydrogen bonding
Long-range RNA/RNA interactions such as A-minor motifs, ribose zippers, nonstandard, and standard base pairs are among the well-known interactions that nowadays can be predicted by software tools. They are essentially made from several HBs that occur between two or more ribonucleotides at the same time. Single HBs, on the other hand, are not always presented as an isolated element by existing tools. They have been previously recognized as an important force that provides stability to the trypanosomatid small segments as they anchor to the main ribosomal scaffold (Rivas and Fox 2022). Detailed analysis of interaction regions revealed that single HBs are a common element that also occur in the interaction regions of E. gracilis and unsegmented ribosomes from Archaea, Bacteria, and other Eukaryotes.
Results presented here pay special attention to the single HBs that were discovered to occur between ribonucleotides from the interaction regions of segmented and unsegmented ribosomes. It was found that among the diversity of such single HBs, those that are established by the 2′OH from the ribose part are by far the most abundant. Their numbers range from nearly 70% to over 90% depending on the organism (Table 5). On average, 2′OH single HBs that occur in domain VI from segmented and unsegmented ribosomes add up to 85% of all single HBs (Table 5). The capacity to form 2′OH single HBs is an intrinsic characteristic of RNA due to its unique chemical structure. More generally the 2′OH clearly contributes to the capacity of RNA to form stable, compact, and intricate structures.
Dominance of the 2′OH while forming single HBs within long-range RNA/RNA interaction regions in segments that form the universal core of domain VI
DISCUSSION
Due to their high conservation, segments srRNA-II and srRNA-IV from the trypanosomatids were chosen to elucidate the possible ancestry of the long-range RNA/RNA interaction regions within domain VI. The segmentation process in E. gracilis (Schnare and Gray 1990; Matzov et al. 2020) releases structural counterparts, LSU-10, LSU-13, and LSU-14 (Fig. 1). Previous reports revealed that most of the interaction regions are also a common feature among segmented ribosomes from trypanosomatids (Rivas and Fox 2022). The prevalence of those interaction regions in other segmented ribosomes, such as E. gracilis, was performed thanks to the availability of these types of ribosome structures in public databases.
Together, srRNA-II and srRNA-IV represent the most conserved part of domain VI, which is recognized as a piece of the universal structural core of the ribosome (Bernier et al. 2018). The latter observation immediately suggests that the search of the already established interaction regions from these two small rRNAs could in principle be extended into the crystallographic structures of unsegmented ribosomes from other Eukaryotes and even Procaryotes. Search and characterization of equivalent interaction regions within unsegmented LSU rRNA was performed. This effort was guided by structural alignments of the two srRNAs from trypanosomatids over their corresponding LSU counterparts. Through this methodology we were able to verify that many of the long-range nonstandard RNA/RNA interaction regions are a common feature that most likely dates back in time into the rRNA of the LCA (Tables 1, 2). It has been previously suggested that protein structural features are more conserved than their sequence. Therefore, they retain a deeper and stronger evolutionary signal (Rivas et al. 2018). We speculate that the same is true for RNA features that remain over time despite sequence variation. RNA's chemical nature most likely enhances these capacities.
Although segmentation phenomenon does not seem to be of any evolutionary advantage, it can be regarded as the consequence of RNA maturation enzymes acting fortuitously over surface-exposed areas as previously suggested (Matzov et al. 2020). The level of conservation of the interaction regions presented here strongly suggests that they predate the emergence of the segmentation phenomenon. We suggest that the conserved regions can be seen as the consequence of a soft exaptive/adaptive process (Frenkel-Pinter et al. 2022) in which these interaction regions initially represented a folding advantage that transitioned into an assembly advantage once the segmentation was established. In other words, the advantage granted by such regions has likely been used in order to retain structure and functionality in the face of such a disruptive phenomenon.
If the latter is true, there must be at least two types of these interaction regions in terms of timing events. One that is most likely old, and which was in place even before the segmentation process took place. The conservation of regions 3–13 from the srRNA-II and regions 1, 3, 5, 6, and 7 from the srRNA-IV strongly suggests that these regions are ancient persistent features of extant ribosomes that arose at least at the time of the LCA (Tables 1, 2). Such interaction regions have been preserved likely because they represent an advantage to the stability and folding of the compact structure of the rRNA. The second type of interaction regions are undoubtably the most recent ones, they likely arose as a consequence of the segmentation process. In these cases, the endoribonuclease that hydrolases the phosphate backbone releases either the 5′-end or the 3′-end as a single-chain free from canonical base pairs. Hence, these ends tend to have long extensions of unpaired ribonucleotides that extend and stick to other distant parts of the ribosome. It seems that these extensions reach stabilization by interacting with other rRNA segments, like srRNA-II R1 and R2 (Table 1; Supplemental Tables S1, S3) and srRNA-IV R2 and R4 (Table 2; Supplemental Tables S2, S4).
RNA unlike DNA is a very dynamic informational polymer with catalytic potential long established (Cech et al. 1981; Guerrier-Takada et al. 1983). The heterogenicity of the elements that integrate the long-range RNA/RNA interaction regions clearly showed its intrinsic flexibility. By using a combination of different nonstandard interactions, small rRNAs can assemble into a fully functional segmented ribosome (Rivas and Fox 2022). Most of these interaction regions seem to be an inheritance from before the segmentation process was established. Among the repertoire of long-range nonstandard interactions are the A-minor motif, the noncanonical base pairs, the ribose zippers, and even the base staking elements that begin with one or more splayed-apart ribonucleotides, which reach parts of the structure that are far apart in their sequence. Even allowing the interaction of independent RNA chains with no complementary base pair involved. In this regard we have suggested that long-range RNA/RNA interactions have been of crucial importance at the very origin of the ribosome, during the RNA world (Rivas and Fox 2020, 2023; Tirumalai et al. 2021).
Notwithstanding the importance of canonical long-range RNA/RNA interactions, when two rRNA elements get near each other, interactions like simple hydrogen bonding seem to play an especially important role. Simple hydrogen bonding can be established by different elements of the ribonucleotides. RNA structures can act as either a proton donor or acceptor. The phosphate backbone can act as a proton acceptor, the ribose can act as both proton acceptor and proton donor, and the nucleobases can also act as both acceptor and donor. The results presented here strongly indicate that the 2′OH from the ribose plays a major role in the formation of single HBs among the interaction regions. It accounts for over 85% of all the single HBs that were identified in the common core of domain VI from segmented and unsegmented ribosomes (Table 5). This is an exclusive characteristic of RNA that undoubtedly contributes to the folding and stabilization of extant segmented ribosomes (Rivas and Fox 2022). Together, the long-range RNA/RNA interactions and the single HBs stablished by the 2′OH most likely have contributed during the early evolution of RNA molecules, possibly since the RNA world.
Conclusions
There are two types of RNA/RNA interaction regions in the segmented ribosomes. The first type is the one that predates segmentation processes. They are conserved and well distributed among unsegmented ribosomes from other Eukaryotes, as well as Bacteria and Archaea. The second type are those interaction regions that arose as a consequence of the segmentation process. They are mainly made from extra ribonucleotides either at the 5′-end or the 3′-end of each chain. These new interactions are of course limited to segmented ribosomes and even among them some are newer forms, exclusive to certain organisms (Tables 1, 2).
Compact, stable, yet flexible RNA structures are the result of the intrinsic nature of the RNA molecule. Long-range nonstandard interactions likely assist in the very beginning of the folding process, even before the RNA associates with rProteins. The intrinsic nature of these interactions being weak when seen individually guarantees a certain level of flexibility, but when seen as a whole, they represent a strong enough force that creates compact and stable RNA structures.
Among all the single HBs that are established in the segmented ribosomes, the ones created by the ribose being so dominant was unexpected. Data presented here contributes to previous observations of this phenomenon in the trypanosomatid segmented ribosomes (Rivas and Fox 2022). The 2′OH being an exclusive characteristic of RNA clearly speaks in favor of this molecule's intrinsic abilities to form complex, compact, and diverse structures. This feature must have been of crucial importance during the early evolution of the ribosome and other equally important RNA systems at the dawn of life. Even further, results of intrinsic RNA features, like the canonical long-range RNA/RNA interactions, support the proposal of RNA being an ancient molecule and DNA being a later development during evolution. It also places chemical constraints on alternative nucleic acids proposals.
MATERIALS AND METHODS
Structure recruitment and selection criteria
Cryo-EM structures from trypanosomatid segmented ribosomes were collected from the Protein Data Bank (Berman et al. 2000) for T. brucei (PDBID 4V8M) (Hashem et al. 2013), L. donovani (PDBID 5T2A) (Zhang et al. 2016), and T. cruzi (PDBID 5T5H) (Liu et al. 2016) as well as for the cryo-EM structure of the highly segmented ribosome from E. gracilis (PDBID 6ZJ3) (Matzov et al. 2020). X-ray crystallographic structures of unsegmented ribosomes from S. cerevisiae (PDBID 4V88) (Ben-Shem et al. 2011), E. coli (PDBID 4V9D) (Dunkle et al. 2011), T. thermophilus (PDBID 4WPO) (Lin et al. 2015), and H. marismortui (PDBID 1FFK) (Ban et al. 2000) were also retrieved from the PDB. These structures were selected because they were obtained free from interaction with foreign molecules such as antibiotics, thereby minimizing unintended alteration of possible RNA/RNA interactions. At the time of collection, the cryo-EM structures of segmented ribosomes were the ones with the highest resolution. Crystallographic information files (CIF) were edited to extract srRNA and rRNA chains without losing their relative position with one another using PyMOL (The PyMOL Molecular Graphics System, version 2.4, Schrödinger, LLC). PDB files containing all srRNA chains from the LSU were obtained for further analysis.
Artificial segmentation of the LSU rRNA
Trypanosomatid segments srRNA-II, srRNA-III, and srRNA-IV constitute almost all the rRNA that forms domain VI of the LSU in the extant ribosome (Fig. 1). Highly conserved srRNA-II and srRNA-IV segments were isolated from T. brucei and structurally aligned with the LSU rRNA from unsegmented ribosomes from S. cerevisiae, E. coli, T. thermophilus, and H. marismortui. The latter allowed the identification of highly conserved structures within domain VI that structurally correspond to these segments. srRNA-II and srRNA-IV equivalents in unsegmented ribosomes were artificially cut from the LSU rRNA without losing their relative position or interactions with respect to the rest of the rRNA using PyMOL. Structural alignments were done with the cealign algorithm implemented in PyMOL. PDB files containing the remaining rRNA from the LSU and the recently cut srRNA-II and srRNA-IV were obtained for further analysis.
Analysis of RNA chains
The rRNA containing files were subject to an automated search for predefined long-range RNA/RNA interactions using the DSSR software v1.8.9 (Lu et al. 2015). This included the A-minor motifs (Nissen et al. 2001) and ribose zippers (Batey et al. 1999; Tamura and Holbrook 2002) formed between different srRNA chains, or single splayed-apart nucleotides that base stack (Batey et al. 1999) or base pair with distant chains. Visual inspection and measurements of single HBs were performed with PyMOL software. Visual inspection was carefully done as a strategy to include long-range nonstandard interactions that are not commonly defined in the available software. Hydrogen bonding distances were included according to the Jeffrey HB classification where they range from strong (<2.5 Å) to moderate (2.5–3.2 Å) to weak (3.2–4.0 Å) (Jeffrey 1997).
Regions of interaction
Several regions of RNA/RNA interaction were previously described for the trypanosomatid srRNA segments (Rivas and Fox 2022). Among those, srRNA-II presented 13, while srRNA-IV presented seven. Long-range interactions as well as simple hydrogen bonding were identified and grouped into equivalent regions for all segmented and unsegmented ribosome structures. Distribution of these interactions is presented in Tables 1 and 2. A comprehensive description of each interaction region is presented in Supplemental Tables S1–S16.
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
Mario Rivas Medrano's research was initially supported by an appointment to the NASA Postdoctoral Program at the NASA Astrobiology Program, administered by the Universities Space Research Association under contract with NASA. This work was also supported in part by a subcontract to the University of Houston from NASA, contract 80NSSC18K1139 under the Center for the Origin of Life, at the Georgia Institute of Technology. The authors would like to thank Dr. Quyen Tran for help in implementing and maintaining computational tools.
Author contributions: M.R. and G.E.F. conceived the work. M.R. conducted all the computational work. Results were discussed with G.E.F. Both authors contributed equally to the writing process and preparation of the manuscript.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079654.123.
- Received March 7, 2023.
- Accepted May 14, 2023.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








