
Which came first: the chicken, the egg, or guanine?
- Department of Chemistry and Biochemistry and Center for RNA Biology, The Ohio State University, Columbus, Ohio 43210, USA
- Institute of Biophysics, Ilia State University, Tbilisi 0162, Republic of Georgia
- Corresponding author: kankia.1{at}osu.edu
Abstract
The main goal of the origin of life (OoL) hypothesis is to reconstruct the missing link between the primordial soup and the extant biology. However, the OoL itself is just the initial part of the link representing the bootstrapping operation of Darwinian evolution. The rest of the link is the emergence of the evolution to the present day primary biological system—the ribosome-based translation apparatus. A valid hypothesis must (i) not invoke Darwinian evolution in the bootstrapping and (ii) transform the ab initio life form into the translation apparatus without violating the principle of continuity (i.e., only incremental steps without foresight). Currently, no such hypothesis exists. Here, I discuss the Quadruplex World hypothesis, which fully complies with these requirements and suggests a spontaneous emergence of the ab initio life form. The spontaneity of OoL arises from the physicochemical properties of guanine monomers in a manner of causal determinism: each step of the process (i.e., scaffolding, polymerization, and folding) is caused by the most recent past step such that in the end only the specific 3D architecture forms. The architecture (i) has a length-independent folding pattern; (ii) can play the role of the predecessor of tRNA and single-handedly conduct a primitive form of translation; and (iii) can evolve into the extant translation apparatus without any paradoxes.
Keywords
INTRODUCTION
We cannot solve our problems with the same thinking we used when we created them (attributed to Albert Einstein).
Before the discovery of the double-helix complementarity principle (Watson and Crick 1953), the solution to the origin of life (OoL) was inconceivable. The complementarity principle turned OoL into a scientific problem through the RNA World hypothesis (Gilbert 1986). This hypothesis posits that RNA can represent a primitive life form by reproducing itself and establishing the genotype–phenotype relationship without any external help, including from proteins. Both functions are achieved through the complementarity principle, which allows RNA to (i) store information (G•C and A•U base pairs used as coding and recognition elements) and (ii) program functional 3D architectures, or ribozymes (base-pairing used as catalytic and structural/building elements) (Fig. 1). This is possible because nucleic acid (NA) secondary structure is formed through hydrogen bonding between complementary nucleobases characteristic of each nucleotide. In contrast, protein secondary structures are formed through hydrogen bonding between the polypeptide backbone—common to all amino acids. As a result, proteins lack programmability and are unable to conceive an OoL hypothesis. Even though the RNA World represents a conceivable hypothesis, it is full of paradoxes and is unable to explain major OoL issues such as polymerization of the very first polynucleotide, breaking chirality, compartmentalization, and metabolism (Fig. 2; Joyce 2002b; Orgel 2004; Koonin 2012; Runnels et al. 2018). This led to suggestions that RNA might be preceded by some workhorse of abiogenesis capable of solving these issues and paving the way for ribozymes. Several attempts have been made to design such molecules; however, these studies could not solve the problems (Joyce 2002a; Orgel 2004; Anastasi et al. 2007). Importantly, all these studies propose artificial double-helical polymers using the complementarity principle.

Establishing the genotype–phenotype relationship by RNA or programmability in the RNA World.
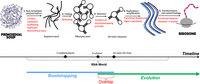
Visualization of the RNA World scenario. Names of the steps are numbered and shown in black; problems and the paradoxes are listed in red. The lower panel shows an approximate timeline of the main events (black) and reveals the main problem of the RNA World—overlap between the bootstrapping (blue) and the evolution (green) due to using the process of natural selection before the arrival of the ab initio life form.
For more than half a century, scientists have attempted to solve the OoL problem exclusively by invoking the same thinking that was used to formulate it—the complementarity principle. Can an alternate paradigm be invoked to resolve the problem? For instance, can a message be written in the NA using recognition elements other than complementarity?
A clue about the programmability of NAs in the absence of the complementarity principle comes from one of the paradoxes of the RNA World—astronomically large sequence space. The complementarity principle requires that the ab initio polymer be produced from the variegated or multicomponent mixtures of nucleotides, which is the sole reason of the paradox. For instance, a standard RNA library size, assuming a four-letter alphabet and a 100-mer ribozyme, is 4100 ≈ 1060. The total weight of such a library, containing only a single copy of each sequence, is 1040 g. For comparison, the age and weight of the Earth are only 1017 sec and 1028 g. Thus, the natural selection of a randomly polymerized RNA library is unimaginable. Shortening of the construct to a 50-mer and reduction to a two-letter alphabet does not reduce the library to a reasonable size (450 = 2100 ≈ 1030). However, further reduction to a single-letter alphabet drops the library to single sequence (1100 = 1n = 1). Of course, this eliminates the possibility of complementarity and appears to be a logical fallacy—existence of a mythical “informational homopolymer” with a strictly defined and length-independent folding pattern. However, such a homopolymer exists—polyG is programmable despite the conventional thinking that only aperiodic polynucleotides can encode information (Kankia 2021a; Kankia 2022). I hypothesize that polyG is the long-sought-after workhorse of the primordial world, which can solve the OoL problem by introducing a new paradigm based on G-tetrads as recognition and structural elements (Fig. 3).

The programmability of polyG using G-tetrads as the recognition (A) and the structural (B) elements. According to the quadruplex domain (QD) folding pattern, a homopolymer (in this case G30) represents specifically arranged G3 and G6 segments (blue) linked by single Gs (black) and it is preprogrammed to fold into two QD domains stacked to each other. The gray discs represent the G-tetrads, and the curved black lines correspond to the Z-shaped (or propeller) loops formed by single Gs.
PROGRAMMABILITY OF POLYG
Programmability is the inherent ability of a polynucleotide to fold into a desired 3D architecture dictated by the information (message) encoded in its nucleotide sequence, as shown in Figure 1. In this paper, the terms “programmability” and “establishing genotype–phenotype relationship” are used interchangeably. For instance, in the RNA World, a genotype (primary sequence of RNA) dictates folding of a phenotype (functional 3D architecture or ribozyme) via the complementarity rules (base-pairing, antiparallel strand alignment, and U-shaped loops) (Fig. 1). In the G4 World, this is achieved via tmNA (tetrahelical monomolecular NA) folding rules (G-tetrads, parallel strand alignment, and Z-shaped G-loops) (see Fig. 3). Both DNA and RNA polyG homopolymers fold into tmNA architectures using G15-segments as the tandem repeat units (Kankia 2014, 2019; Kankia et al. 2016; Lomidze et al. 2022). Each G15-segment contains a stably folded QD with three G-tetrads and three Z-shaped (or propeller) G-loops (Fig. 3; Sengar et al. 2014). Each QD is stacked on its neighbors forming an uninterrupted tetrahelix. As a result, polyG represents not just a regular homopolymer but a specifically programmed sequence—G3 and G6 segments (Fig. 3B, blue) connected by G-loops (black). This transforms the polyG or Gn homopolymer into the tmNA architecture comprised of n/15 QDs, or a poly(QD).
Both structural motifs of the QD, the G-tetrads and loops, consist of guanines. Hypothetically, the length of the repeat unit could be different from G15. For instance, a domain with two G-tetrads and single-nucleotide loops would contain G11 repeats, while a domain with four G-tetrads and 2-nt loops would use G22 as the repeat unit. However, the folding pattern shown in Figure 3B (i.e., three G-tetrads with single G-loops) is invariant (polyG is not able to use any other folding pattern) and is not affected by any changes in the polyG length or experimental conditions (Kankia 2018). Thus, the folding pattern of polyG is unambiguously defined.
Since tmNA has domain assembly, the function of the phenotype is independent of the polyG length and elongation is accompanied by extending the tmNA architecture without structural reorganization. Therefore, due to the feature of domain assembly, the phenotype in the G4 World is length-independent (1n = 1). Thus, the simple homopolymer polyG is capable of unambiguously establishing the genotype–phenotype relationship.
SPONTANEOUS FORMATION OF THE AB INITIO LIFE FORM
Since the OoL cannot invoke evolution, it must be a spontaneous process (Darwin 1860; Cairns-Smith 2008). But despite this simplicity, the nonliving components of the ab initio polynucleotide had to solve the following fundamental problems: (i) scaffolding, (ii) polymerization, (iii) breaking chirality, (iv) separating genotype from phenotype, or avoiding functional duality, and (v) compartmentalization. Moreover, these issues had to be addressed without any external help using a source of energy readily available in the primordial world (e.g., temperature fluctuations) (Deamer and Weber 2010; Salditt et al. 2020).
Free G-monomers (guanine bases and their derivatives—nucleosides or nucleotides with both ribose or deoxyribose sugars) meet all of the requirements of the OoL hypothesis due to their unique physical and chemical properties: (i) the 90° angle between the Watson–Crick and Hoogsteen edges and ideal distribution of the hydrogen bond donor–acceptor groups resulting in self-assembly into the G-tetrad (Fig. 3A); (ii) the partially negatively charged O6 carbonyl groups pointing toward the center of the G-tetrads resulting in cation chelation between neighboring G-tetrads; and (iii) strong stacking interactions between G-tetrads with 30° right helical turns resulting in the maximum interconnection between Gs (each base has direct contact with all six neighbors). As a result, in the presence of specific cations with a coordination number of eight (e.g., K+), free G-monomers assemble into a crystalline structure with the identical helical properties as quadruplexes folded from polynucleotides (Zimmerman 1976; Wu and Kwan 2009; Chen et al. 2020). G-tetrads do not simply stack with each other but juxtapose the G-monomers into the right position to catalyze self-polymerization (Fig. 4, steps 1, 2, and 4) (Pino et al. 2008; Costanzo et al. 2009; Morasch et al. 2014). The crystalline structures are both thermodynamically and kinetically stable (Wu and Kwan 2009; Smith et al. 2018), which facilitates their polymerization at higher temperatures by eliminating the activation barrier (Kankia 2021a,b). Thus, the crystalline structure represents a catalytic system capable of driving self-polymerization using the most abandoned primordial energy source—temperature fluctuation.

Suggested steps between the primordial world and the ribosome. The lower panel shows an approximate timeline of the main events (black) and colocation of the bootstrapping (blue) and the evolution (green).
The temperature cycles could not only drive the polymerization but could also be used to purify the crystalline structures and break the chiral symmetry (Fig. 4, step 3). While the impurities (i.e., other bases) could be removed relatively easily due to their nonspecific interaction with G-tetrads, establishing homochirality seems more challenging since both d- and l-enantiomers, d-G and l-G, are expected to assemble into G-tetrads with equal affinity. However, in the right-handed crystalline structures, only two adjacent d-Gs can ligate to each other through the standard 5′–3′ phosphodiester backbone. For l-G monomers incorporated into the right-handed helix, it is sterically impossible to achieve the 5′–3′ orientation (Joyce et al. 1984). Therefore, l-G-monomers would terminate polymerization by not ligating to oligo(d-G)s or ligating in the wrong orientation and capping them (Joyce et al. 1984; Kankia 2021a). Thus, after the first cycle of ligation, the crystalline structures would contain some amount of oligo(d-G)s with proper backbone linkages; upon the subsequent temperature cycle, they can dissociate from the mixed stacks and reassemble into the thermodynamically more favorable homochiral assemblies (Fig. 4, step 3).
Interestingly, only G-monomers and oligomers shorter than the critical length, G15, are involved in the polymerization process (Fig. 4, steps 1–4). Once the critical length is reached, oligomers fold into the QDs (steps 5 and 6) and are removed from the production process. Thus, in the G4 World, the production of the genotype and the phenotype are two separate processes resolving the paradox of functional duality (Ivica et al. 2013); production of the genotype is a template-free process primed by the self-assembly of the G-monomers.
Since the production of the genotype in the G4 World is the simultaneous coproduction of four identical polymers without any parent–offspring relationship, it is a massively parallel process—practically all G-monomers can be polymerized within a few temperature cycles. Thus, the entire pool/reservoir (i.e., the primordial soup) can be transformed into a gigantic communal cell (Woese 2002)—QDs surrounded by amino acids and other components of the primordial soup.
In summary, the G4 World addresses all the fundamental problems of the OoL before even introducing the complementarity principle.
FUNCTION OF THE AB INITIO LIFE FORM
Any OoL hypothesis must reconstruct the missing link between the primordial soup and the translation apparatus (Smith et al. 2008; Bowman et al. 2015; Bowman et al. 2020). The link can be extended in both directions—bottom-up or top-down. The previous section describes the bottom-up approach—the emergence of polyG from G-monomers and establishing the very first genotype–phenotype relationship (Fig. 4, step 6). The question arises: What was the function of the ab initio QD?
The QD presumably had to play the role of tRNA, the adapter molecule in protein synthesis. There are some morphological similarities between QDs and tRNAs (Kankia 2022), but the most compelling reason that QDs may be predecessors of tRNAs comes from the top-down approach (i.e., the step-by-step simplification of ribosomes to the ab initio polymer). The reduction process reaches a dead end quickly—deletion of even noncatalytic components (i.e., ribosomal proteins) results in a complete loss of peptidyl transferase activity (Khaitovich et al. 1999; Bowman et al. 2020). This is because the ribosome is not a typical enzyme that functions simply to accelerate a reaction rate above the background (no enzyme) rate. It instead enforces the sterically unfeasible process—simultaneous attachment of two tRNAs (with an anticodon stem–loop helical diameter of 2.3 nm) to two adjacent codons (which span ∼1 nm length). In the absence of external help, this could be achieved by using either (i) longer codons to allow the assembly of tRNAs along mRNA without skipping any nucleotides between the codons, or (ii) an earlier adapter (proto-tRNA) with strictly defined geometry matching the triplet codon length. The former can be excluded since the codon length variation would have a lethal effect and require a false start (Crick 1968). Thus, we are left with the latter scenario. Based on the following properties, QD is the ideal candidate for the role of the proto-tRNA: (i) invariance of the folding pattern (only three G-tetrads with single-nucleotide G-loops, see Fig. 3) perfectly matching the length of a triplet codon; (ii) fully accessible hydrophobic ends for the higher-order stacking assembly; (iii) unusual Z-shaped loops with exposed bases for hybridization to proto-mRNA; and (iv) potential amino acid binding pockets between the grooves and the Z-loops (Kankia 2022). Based on these properties, poly(QD) alone or a multimolecular QD-assembly in the presence of a proto-mRNA (Fig. 4, step 7) could provide a scaffold for peptide synthesis and conduct primitive translation.
Thus, the de novo emerged QD architecture lays the foundation of the protein scaffold and establishes the structural (grammar) rules of the extant translation apparatus—triplet coding with nonoverlapping and comma-less reading mechanism.
FROM PRIMITIVE TO EXTANT TRANSLATION (G4 → RNA TRANSFORMATION)
The QD is arguably the most stable NA structural motif (Jing and Hogan 1998; Kankia 2014). The unprecedented stability is attributed to the highly organized 3D arrangement of Gs, similar to the stacking of bricks in a wall—each G has direct contact with six neighboring bases (two from the same G-tetrad via hydrogen bonds and four from the adjacent G-tetrads via stacking interactions). In addition, diagonally positioned Gs have indirect contact via cation coordination.
Here are two obvious questions: How does such well-packed and stable architecture transform into the less stable RNA? Why do we not see QDs in the extant rRNA? The answer to both questions is in the vulnerability of the QD architecture to random nucleotide modifications (insertion, deletion, or substitution), resulting in dramatic destabilization effects, which lead to all-or-none structural outcomes. For instance, any base substitution in the G-tetrad positions (12 out of the 15 bases) is accompanied by a complete unfolding of QD (Kelley et al. 2011; Kankia et al. 2016). Such substitutions could be facilitated by thermal cycles: G bases dissociate upon heating, while pyrimidines, characterized by more stable glycosidic bonds, are substituted for the Gs upon cooling. This inhibits quadruplex folding and introduces complementarity via G•C and G•U base-pairing, allowing almost irreversible G4 → RNA transformation.
In addition, very stable and rigid quadruplexes could not conduct efficient catalysis in extant biochemistry, which usually requires flexible 3D architectures capable of rapid structural rearrangements. Therefore, even if initially the QDs were part of rRNA, the selection pressure for efficient enzymes would transform them into RNA via nucleotide substitutions. Similarly, QDs needed to be transformed into more flexible tRNA with higher specificity toward amino acids and mRNA to increase the translation specificity. Therefore, production of polyG (Fig. 4, steps 1–5) and polypeptides (step 7) would be accompanied by the emergence of complementarity-based RNA structures, presumably proto-rRNA. Thus, the G4 World would be followed by the almost simultaneous arrival of RNA and polypeptides. Based on this scenario, another obvious question arises: Why is extant translation catalyzed by RNA and not by superior protein catalysts?
While the production of QD or poly(QD) is a predetermined/programmed event, the transformation of quadruplexes into complementarity-based RNA structures and the formation of the first peptides must be largely random processes. Hypothetically, the RNA resulting from random base substitutions in polyG and the primitive translation apparatus (Fig. 4, step 7) represents a rather nonspecific scaffold for amino acid polymerization. While the combinatorial chemistry of RNA can lead to functional and soluble molecules (i.e., aptamers) (Ellington and Szostak 1990; Tuerk and Gold 1990), random peptide sequences, especially those made up of simple amino acids, rarely form an active molecule and tend to aggregate (Wei et al. 2003). Therefore, during the early days of molecular evolution, RNAs were more efficient enzymes than proteins. In addition, base-to-base contact is the simplest way to establish the specific interactions between RNA and the components of the primitive translation apparatus, which are all NAs.
G4 WORLD VERSUS RNA WORLD
The question of the OoL is also the question of the origin of Darwinian evolution; the evolution can take over only after ab initio life forms appear. Therefore, OoL itself, representing only the first segment of the missing link between the primordial soup and the ribosome (blue part on the timeline shown in lower panel of Fig. 4), should be a spontaneous process that proceeded without external help. The second segment of the timeline (green) represents the emergence of the evolution, which leads to the ribosome. Thus, there are two main requirements for a valid OoL hypothesis: to build the first segment of the link (blue) without any external help and to build the remainder (green) without violating the continuity principle (only Darwinian steps without foresight and miracles) (Orgel 1968; Koonin 2012).
The G4 World fully complies with the requirements: (i) Free G-monomers self-scaffold, polymerize and establish the genotype–phenotype relationship in a manner of causal determinism—nothing other than the QD architecture is formed. The process does not have any alternative, so no selection/evolution is involved; (ii) the QDs play the role of the protein scaffold, establish the grammar rules of the genetic code (triplet codon length, nonoverlapping, and comma-less reading), and launch the primitive translation; and (iii) base substitutions in oligoG sequences introduce the complementarity principle, which generates the QD and mRNA diversity. It is at this point that evolution emerges and starts honing and fine-tuning the primitive translation system (Fig. 4, step 8).
In contrast to the G4 World hypothesis, which addresses all OoL-related questions, the RNA World requires help from evolution before the appearance of the ab initio life form and is unable to address any major OoL questions without violating the continuity principle (for the problems and violations see Fig. 2). A major flaw of the RNA World hypothesis is the primary assumption that the ab initio ribozyme uses the complementarity principle, even though the principle is too complex to appear de novo (Kankia 2021a, 2022). Interestingly, all of the RNA World paradoxes and problems are related to the properties of the complementarity principle: (i) multicomponent nature leading to astronomical sequence pool sizes (see Introduction); (ii) template-based reproduction, creating Eigen's (Eigen 1971) and functional duality (Ivica et al. 2013) paradoxes; and (iii) structural and chemical instability of RNA, requiring external help for scaffolding (Ferris et al. 1996; Cairns-Smith 2008) and creating cyclization (Horowitz et al. 2010) and hydrolysis (Deamer and Weber 2010; Runnels et al. 2018) problems.
The premature launch (or anachronism) of the complementarity principle ties the RNA World hypothesis into an unsolvable “Gordian knot”; any attempt to solve the problems within the complementarity principle further complicates it (Orgel 2004). Beginning the OoL process with single-component and stable quadruplexes, capable of template-free amplification, sets the chronology right and creates a space for complementarity rules to evolve. As a result, the G4 World hypothesis fully untangles the knot and creates a paradox-free OoL scenario.
It must be emphasized that while quadruplexes preceded the complementarity rules, the G4 World does not represent a prelude to the RNA World—the world of ribozyme-governed life forms functioning in the absence of proteins does not exist in this scenario. This becomes obvious upon segmentation of the G4 World timeline into three main periods (Fig. 4):
-
G4 period from stacking of free G-monomers to the first peptide scaffolds (steps 1–6), which fully relies on G-tetrads and quadruplexes.
-
G4-RNA–peptide period with added complementarity and peptide formation. The base-pairing initiates recognition between mRNA and QDs and creates rRNA (major catalytic component of the system and only surviving ribozyme of the extant biology). In parallel, the first noncatalytic peptides (proto-ribosomal proteins) emerge via primitive translation, chaperoning the translation apparatus.
-
RNA–protein period where (i) quadruplexes become incorporated into double-stranded helices (see the following section) and disappear; (ii) translation becomes a complementarity-governed process; and (iii) protein enzymes evolve and become the main catalysts. This ideally addresses another unanswered question of the RNA World: Why is only the ribosome preserved as a ribozyme in the extant biology? (Bowman et al. 2015). Finally, DNA emerges, probably through the process of reverse transcription, and the gap between the primordial soup and the extant biology fully closes.
EXPERIMENTAL FEASIBILITY OF THE G4 WORLD HYPOTHESIS
The G4 World scenario is not only sterically and thermodynamically realistic, but many of the steps are already experimentally tested. Specifically, the G-tetrad crystalline structures of free G-monomers (step 1, Fig. 4) with helical parameters of quadruplexes were observed for GMPs (Zimmerman 1976; Wu and Kwan 2009; Chen et al. 2020). Nonenzymatic ligation of the crystalline structures with a canonical 5′–3′ phosphodiester backbone (step 3) was demonstrated for 3′,5′-cyclic GMP (Pino et al. 2008; Costanzo et al. 2009; Morasch et al. 2014). The ligation of G-wires (step 4) was demonstrated on (dG)10 in the presence of EDC (carbodiimide) (Kankia 2021b). Folding of QD and tmNA architecture (step 6) from both poly(dG) and poly(rG) was demonstrated earlier (Kotlyar et al. 2005; Kankia 2014, 2018).
Future experiments might include the demonstration of: (i) self-scaffolding (the crystalline structure formation) of free bases, nucleosides or nucleotides followed by ligation of the monomers; (ii) the origin of homochirality using equimolar mixtures of both sugar enantiomers; (iii) oligomer ligation in aqueous solution (in the absence of condensing agents); (vi) the stacking of QDs in the presence of mRNA (or QD-assembly); and (v) peptide bond formation in the presence of QD-assembly or tmNA.
VESTIGES OF G4 WORLD
After introducing base-pairing rules and replicases, the quadruplexes became easily invadable targets by the complementary strands to incorporate them into the double helixes. Therefore, despite numerous G-rich segments, presented in all the genomes and capable to fold in quadruplexes in vitro, it is hard to detect folded quadruplexes in vivo (Rhodes and Lipps 2015). This echoes with Darwin's assumption that “a proteine compound [polyG] was chemically formed, ready to undergo still more complex changes [adding other nucleotides and the complementarity principle], at the present day such matter would be instantly devoured [quadruplexes being invaded by the complements]” (letter to Joseph Hooker,1871). So, it is problematic to detect direct vestiges of the G4 World. However, a trace of the G4 World had to be preserved in the genetic code, representing a set of rules determining the translation process. According to the continuity principle, newly arrived rules/features had to be added to already existing ones without rewriting the latter to avoid false starts. Indeed, GGG triplet codes glycine, the simplest and only achiral amino acid, and plausibly initiates evolution of the genetic code (Hartman 1975; Kankia 2021a). However, the most convincing vestige of G4 World must be the grammar rules of the extant genetic code predetermined by the unique properties of the QD architecture (see section “Function of the ab initio life form”).
CONCLUSIONS
G4 World represents a paradox-free alternative to the RNA World hypothesis, capable of explaining the spontaneity of OoL and addressing all the OoL-related questions without violating the continuity principle. The hypothesis initiates the process with the standard NA chemistry but uses G-tetrads as the recognition and structural elements instead of the base pairs. The complementarity principle and the heredity emerge/develop later. Through the G-tetrads, polyG establishes the genotype–phenotype relationship. The emergence of the ab initio life form, or QD architecture, is predetermined by the physical and chemical properties of the G-monomers. The architecture could play the role of proto-tRNA, single-handedly conduct a primitive translation, and evolve into the extant translation apparatus.
ACKNOWLEDGMENTS
I thank Drs. Karin Musier-Forsyth (The Ohio State University), Dipankar Sen (Simon Fraser University, Vancouver), Jean-Luis Mergny (Ecole Polytechnique, Palaiseau, France), James Kadonaga (University of California, San Diego), and Ulrich Göringer (Technical University Darmstadt, Germany) for comments and discussions. I would like to thank Tamari Kankia for constructive criticism and improvements in the writing of the manuscript.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079613.123.
- Received January 30, 2023.
- Accepted May 23, 2023.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








