
Occurrence and classification of T-shaped interactions between nucleobases in RNA structures
- 1Computational Biochemistry Laboratory, Department of Chemistry and Centre for Advanced Studies in Chemistry, Panjab University, Chandigarh 160014, India
- 2Department of Chemistry and Biochemistry, University of Windsor, Windsor, Ontario N9B 3P4, Canada
- 3Center for Computational Natural Sciences and Bioinformatics, International Institute of Information Technology, Gachibowli, Hyderabad, Telangana, 500032, India
- Corresponding authors: j.trant{at}uwindsor.ca, psharma{at}pu.ac.in
Abstract
Understanding the frequency and structural context of discrete noncovalent interactions between nucleotides is of pivotal significance in establishing the rules that govern RNA structure and dynamics. Although T-shaped contacts (i.e., perpendicular stacking contacts) between aromatic amino acids and nucleobases at the nucleic acid–protein interface have recently garnered attention, the analogous contacts within the nucleic acid structures have not been discussed. In this work, we have developed an automated method for identifying and unambiguously classifying T-shaped interactions between nucleobases. Using this method, we identified a total of 3261 instances of T-shaped (perpendicular stacking) contacts between two nucleobases in an array of RNA structures from an up-to-date data set of ≤3.5 Å resolution crystal structures deposited in the Protein Data Bank.
Keywords
INTRODUCTION
Highly complex RNA 3D architectures are supported by an array of intramolecular forces (Holley et al. 1965; Tinoco and Bustamante 1999), which include noncovalent interactions such as pairing (i.e., hydrogen bonding) and parallel π−π stacking of bases (Fig. 1; Šponer et al. 1996; Leontis and Westhof 2001; Sharma et al. 2007; Sarver et al. 2008; Stombaugh et al. 2009; Havrila et al. 2013; Waleń et al. 2014; Jhunjhunwala et al. 2021). Additional higher order hydrogen bonding (such as triplets and multiplets, Das et al. 2006; Abu Almakarem et al. 2012) and stacking (e.g., base-intercalated and base-wedge interactions, Baulin et al. 2020) motifs determine RNA architecture. However, other interactions are also significant for RNA structure and function (Zirbel et al. 2009; Ulyanov and James 2010; Šponer et al. 2012). These include the interaction of metal ions with the phosphate groups, aiding RNA folding by minimizing the electrostatic repulsion (Agris 1996; Misra and Draper 1998; Draper 2004; Kolev et al. 2018), base–phosphate interactions (Zirbel et al. 2009), and T-shaped RNA inter-nucleobase interactions (Fig. 1). This last class, far less studied than the others, involves interactions between two perpendicularly interacting aromatic rings, in which one nucleobase skeleton provides the π face (denoted the “horizontal base” in the subsequent text) and the other base an aryl group that interacts via its edge (denoted the “vertical base,” Fig. 1C).
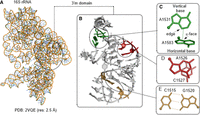
(A,B) Representation of the structure of 16S rRNA structure (PDB code: 2VQE) belonging to the 30S ribosomal subunit of Thermus thermophilus, highlighting the nucleobase-specific noncovalent interactions in its 3′m domain (H45). (C–E) Examples of T-shaped (C), base stacking (D), and base-pairing (E) interactions.
After their initial discovery in aromatic organic compounds (Sinnokrot and Sherrill 2004; DiStasio et al. 2007; Geng et al. 2010; Hobza et al. 1990, 1996; Arunan and Gutowsky 1993; Paulini et al. 2005; Jaffe and Smith 1996; Sinnokrot et al. 2002; Tsuzuki et al. 2002), T-shaped interactions were identified in biomolecules, for example, between two aromatic amino acids in proteins (McGaughey et al. 1998; Chelli et al. 2002), and between an aromatic amino acid and a nucleobase in nucleic acid–protein complexes (Rutledge and Wetmore 2008; Rutledge et al. 2009, 2010; Wilson et al. 2014, 2016). For example, Wilson et al. (2014, 2016) identified T-shaped interactions in a large crystal structure data set of DNA–protein complexes and revealed their presence in the active sites of crucial enzymes. Rutledge et al. (2009) analyzed T-shaped interactions between canonical nucleobases and aromatic amino acids using quantum mechanical calculations and revealed that their magnitude can be close to that of hydrogen bonds. The effects of methylation and the nature of monomer edge and face on T-shaped interactions were also analyzed in nucleic acid–protein complexes (Rutledge and Wetmore 2008). Another class of perpendicular π-interactions previously known in DNA (Egli and Gessner 1995; Wilson et al. 2014), and recently studied in RNA (d'Ascenzo et al. 2016), are the lone pair-π interactions that involve the interaction between nucleobase π-ring and the sugar atoms (Chawla et al. 2017) or water molecules (Kalra et al. 2020). However, no structural analysis exists on the T-shaped interactions between two nucleobases in RNA structures. As these are clearly critical in certain contexts, without this analysis, we cannot obtain a full picture of the discrete noncovalent interactions within RNA complexes.
In this present work, we build on the existing RNA base pairing and parallel π−π stacking classification schemes to propose a simple and unambiguous geometry-based theoretical framework for the classification of T-shaped interactions in RNA (Leontis and Westhof 2001; Jhunjhunwala et al. 2021). We propose a four-stage hierarchy to classify them: (1) identity of the interacting π face of the horizontal nucleobase; (2) identity of the interacting edge and the interacting atom of the vertical nucleobase; (3) mutual orientation of the glycosidic bonds of the two bases; and (4) the identity of the interacting ring(s) (five-membered, six-membered, or both) that constitute the horizontal base. Using this classification scheme in conjunction with the appropriately adapted existing approaches toward the detection of parallel π−π stacks (Jhunjhunwala et al. 2021) and T-shaped contacts (Wilson et al. 2014) in macromolecular 3D structures, we developed an algorithm for the automated identification and classification of examples of different T-shaped contacts in RNA crystal structures.
In this article, we describe the classification scheme and the algorithm. We then apply this scheme to analyze the occurrence and distribution of T-shaped interactions in a nonredundant data set of 2286 RNA structures (Supplemental Table S1) with respect to the different structural parameters. We draw attention to the role that the T-shaped associations appear to play in connecting RNA secondary structural units in the 3D space. Our data reveals that these interactions play widely different roles in determining structure in different RNA types. This serves as a starting point for further multiscale investigations and/or simulations of RNA folding processes, and greater appreciation of the functional role of these interactions in biology.
RESULTS AND DISCUSSION
Classification of T-shaped interactions involving nucleobases
We searched for T-shaped interactions in an up-to-date data set of RNA crystal structures (see Materials and Methods section). The detected T-shaped interactions were classified in several stages. In the first three stages, the interactions were assigned to a basic geometric family based on the edge of the vertical base, the face of the horizontal base and the mutual glycosidic orientation of the interacting nucleotides (Fig. 2A–C). In the last stage, the geometries were assigned to subclasses based on the participation of the (five-membered, six-membered, or both) rings that constitute a purine positioned horizontally in a T-shaped interaction (Fig. 2D), and the interacting atom belonging to the participating edge of the vertical base.
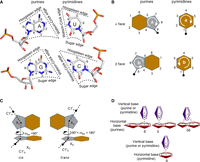
Representation of (A) edges, (B) faces, (C) relative glycosidic orientations, and (D) identities of the horizontal rings participating in a T-shaped interaction. The “b” face of purines and the “a” face of pyrimidines is presented in panel A. σhv is the torsion angle involving the C1′ atom of the ribose sugar of the horizontal base, the centroid of ring (five-membered ring in purines and six-membered ring in pyrimidines) of the horizontal base, the centroid of ring of the vertical base and the C1′ atom of the ribose sugar of the vertical base. See Materials and Methods section for details.
Basic T-shaped geometric combinations
Combination of the interacting edge and face
A T-shaped contact in RNA involves the Watson–Crick (W) edge, Hoogsteen (H) edge, or the Sugar (S) edge of the vertical base and the π-containing face (alpha [a] or beta [b]; Rose et al. 1980) of the horizontal base. For instance, if the “W” edge of the vertical base interacts with the “a” face of the horizontal base, the T-shaped interaction will be annotated as Wa or aW, depending on the order which one chooses to list the participating nucleotides. Each of the 16 sequence combinations (i.e., four purine–purine [A-A, A-G, G-A, and G-G], four purine–pyrimidine [A-C, A-U, G-C, and G-U], four pyrimidine–purine [C-A, C-G, U-A, and U-G], and four pyrimidine–pyrimidine [C-C, C-U, U-C, and U-U]) can have six distinct face-edge pairs: aW, aH, aS, bW, bH, and bS, or 96 total possible combinations. Further, in addition to canonical ribonucleotides, this annotation system is applicable to nucleotides containing base modifications (Chawla et al. 2015; Seelam et al. 2017), as well as deoxyribonucleotides of DNA.
Relative glycosidic torsion angle
In analogy with the previously proposed RNA base pairing (Leontis and Westhof 2001) and stacking (Jhunjhunwala et al. 2021) classification schemes, T-shaped interactions can be divided in terms of the relative glycosidic torsion angle (σhv). Specifically, a T-shaped interaction can be annotated as cis if 90° > σhv ≥ 0° and trans if 180° ≥ σhv ≥ 90° (Fig. 3). This leads to 12 basic T-shaped geometric families for each of the 16 nucleobase sequence combinations (Table 1, vide supra).
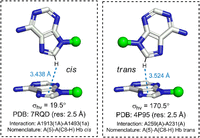
Examples of cis and trans T-shaped interactions. PDB code, crystallographic resolution, interaction code (horizontal base [chain id]—vertical base [chain id]) and complete nomenclature (i.e., vertical base [interacting atom]—horizontal base [interacting ring], edge of the vertical base, face of the horizontal base, and the relative glycosidic orientation) are provided for each interaction. Dotted line indicates the T-shaped interaction between the C-H group of the vertical base involving the Hoogsteen edge and the ring of the vertical base. The distance between the carbon atom of the C-H group of the vertical base and the center of the interacting ring of the horizontal base is provided. Although one atom belonging to the interacting edge may visually appear closer than the other atom of the same edge, the interacting atom is that which is closest to the centroid of the receiving base. This was identified using the unbiased quantitative method according to the algorithm described in the Materials and Methods section. Note that including the information on the ring identity of the horizontal base and the interacting atom/group of the vertical base is optional and can be removed in ambiguous cases.
Symbols for representations of 12 geometric families of T-shaped interactions
Identity of the interacting atom of the vertical base and the participating ring of the horizontal purine
The geometry of the T-shaped interaction can be further specified by including the identity of the interacting atom belonging to the participating edge of the vertical base (Supplemental Table S2). Further, depending on whether the five-membered, six-membered, or both purine rings participate, a T-shaped interaction involving a purine at the horizontal position can be further categorized by adding “5,” “6,” or “56” to the original face-edge nomenclature (Fig. 2D). For example, the A-C aW cis contact involving the interaction of the “5” membered ring of A and the N3 atom of C can be designated as A(5)-C(N3) aW cis. The interacting edge is definitive in the structures, but we note that in some cases the interacting atom is not so clear, and small distortions in the crystal structure could change which atom is primarily involved in the centroid interaction. The identity of the interacting atom must, consequently, be assigned with care, and we suggest a visual inspection of any interaction of interest to determine if small dynamics could shift the interaction. It can prove a very useful tool to compare two different structures of the same RNA molecule where subtly different crystallizations changed the observed interaction.
Overall, in analogy with the stacking classification scheme (Jhunjhunwala et al. 2021; Ali et al. 2023), combining six face-edge geometries, three ring identities and two glycosidic bonds, each purine–purine or purine–pyrimidine (purine as the horizontal base and pyrimidine as the vertical base) T-shaped interaction can have 36 (6 × 3 × 2) structural possibilities. However, each pyrimidine–purine or pyrimidine–pyrimidine interaction will have 12 (6 × 2) distinct possibilities. Depending on the base sequence, purine–purine and purine–pyrimidine interactions can have 288 total possibilities (36 each for A-A, A-G, G-A, G-G, A-C, A-U, G-C, and G-U), whereas pyrimidine–purine and pyrimidine–pyrimidine interactions can have 96 possibilities in total (12 each for C-A, C-G, U-A, U-G, C-C, C-U, U-C, and U-U). This gives rise to a total of 384 distinct theoretically possible T-shaped arrangements (Fig. 4), although these possibilities can further increase if the identity of the interacting atom of the vertical base is considered (Supplemental Table S2).
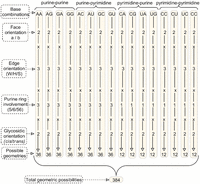
Possible arrangements of the bases in T-shaped interactions, classified with respect to the base-edge orientation, glycosidic orientation and the ring involvement of the base providing the face.
Identification of T-shaped interactions in RNA crystal structures
We used the proposed scheme to classify the T-shaped interactions present in the entire data set of RNA crystal structures
(see the Materials and Methods section). This open-source python-based identification and classification tool is accessible
through github (github.com/PSCPU/T-shaped interaction). The program generates an output file containing classification of
each T-shaped contact present in an RNA PDB file, along with the values of geometrical parameters (i.e., 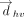 , θhv, τh, and τv) that characterize the interaction. A sample output file containing the description of T-shaped contacts identified in a
PDB is provided in the Supplemental Information (Supplemental Fig. S1). Consolidated output file containing all identified T-shaped interactions in the total data set is also provided as a separate
Supplemental Text File.
, θhv, τh, and τv) that characterize the interaction. A sample output file containing the description of T-shaped contacts identified in a
PDB is provided in the Supplemental Information (Supplemental Fig. S1). Consolidated output file containing all identified T-shaped interactions in the total data set is also provided as a separate
Supplemental Text File.
In addition to the information of base identities and face, ring, edge and glycosidic combinations of the nucleotides participating in T-shaped interactions, the output files contain information on whether the interacting bases are in consecutive positions in sequence space or separated by other nucleotides. Thus, depending on the difference between the nucleotide residue numbers, each T-shaped interaction is annotated as consecutive, nonconsecutive, or inter-RNA (Fig. 5). Though T-shaped examples between adjacent or nonadjacent nucleotides are not expected to be geometrically different, this distinction can help clarify the functional role these contacts play in different RNA regions. Further, the output file provides the identity of the atom of the interacting edge of the vertical base that is closest to the face of horizontal base in the T-shaped interaction, to help better understand the nature of atoms involved in the interaction (protonated or nonprotonated oxygen and nitrogen, and the C-H group, Supplemental Table S2). A total of 3261 instances of T-shaped interactions were found in the data set of 2286 RNA crystal structures (see Materials and Methods section), although these interactions appear in just under one-fifth (453, i.e., 19.9%) of the structures (Supplemental Tables S3, S4). These are not as ubiquitous as the stacking interactions we classified (Jhunjhunwala et al. 2021), so it is suggestive that they might play important roles, especially as they imply a tight 90° kink in an RNA strand when present between consecutive nucleotides.
Representation of T-shaped interactions based on the relative positioning of the participating nucleobases in the sequence space.
The structures containing T-shaped interactions span all biota, from viruses (e.g., PDB: 2GTT, the nucleoprotein–RNA complex of the rabies virus) to archaea (e.g., 1KQS, the 50S ribosomal subunit of Haloarcula marismortui), bacteria (e.g., 1FJG, the 30S ribosomal subunit of Thermus thermophilus), and eukarya (e.g., 4KR2, the tRNAgly-glycyl-tRNA synthetase complex in Homo sapiens) (Table 2). Perhaps unsurprisingly, extant T-shaped interactions are primarily observed in rRNA-derived structures (94.3%), although nonribosomal structures, mostly involving viral RNA structures (1.3%) and tRNA (0.9%) also show T-interactions (Table 2). This suggests that T-shaped interactions are more frequent in large RNA structures. It is important to note that there is an inherent selection bias in the data set. Crystallographers are more likely to be interested in specific RNA systems, and certain RNA sequences are also more likely to crystallize. T-shaped structures might be prevalent in other forms of RNA that have not been examined, but with the unusual conformation that they demand, it is reasonable that they would be more common in RNA macromolecules that play a catalytic or complex molecular interaction role; they would not be likely or necessary in most classical coding mRNA for instance.
Overview of the 453 PDB structures containing T-shaped interactions, categorized with respect to RNA type, source organism, the total count of T-shaped interactions, and the count of T-shaped interactions occurring in multiple independently solved structures
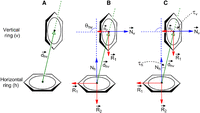
In terms of geometrical parameters, the inter-centroid distance ( ) (Fig. 6) for the majority (71.6%) of T-shaped interactions lies between 4.5 and 5.0 Å, with the plurality between 4.8–4.9 Å (Supplemental Fig. S2A). Dissecting the interactions based on the interacting atom/group (i.e., C-H, N, NH, NH2, or O) belonging to the participating edge of the vertical base reveals that although the interactions involving the C-H
(i.e., C2-H of A, C8-H of purines, and C5-H and C6-H of pyrimidines), N (N7 of purines, N1 of A and N3 of C) and N-H (N1-H
of G and N3-H of U) groups have a similar spread of
) (Fig. 6) for the majority (71.6%) of T-shaped interactions lies between 4.5 and 5.0 Å, with the plurality between 4.8–4.9 Å (Supplemental Fig. S2A). Dissecting the interactions based on the interacting atom/group (i.e., C-H, N, NH, NH2, or O) belonging to the participating edge of the vertical base reveals that although the interactions involving the C-H
(i.e., C2-H of A, C8-H of purines, and C5-H and C6-H of pyrimidines), N (N7 of purines, N1 of A and N3 of C) and N-H (N1-H
of G and N3-H of U) groups have a similar spread of  (3.8–5.0 Å), interactions involving NH2 (N2-H2 of G, N6-H2 of A and N4-H2 of C) and carbonyl O (O6 of G, O2 of pyrimidines and O4 of U) groups have
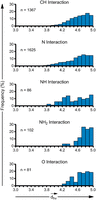
(3.8–5.0 Å), interactions involving NH2 (N2-H2 of G, N6-H2 of A and N4-H2 of C) and carbonyl O (O6 of G, O2 of pyrimidines and O4 of U) groups have 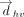 values in the range of 4.2–5.0 Å (Fig. 7). However, θhv (Fig. 6) remains relatively constant within the interval, with small peaks at 80° to 81° and 88° to 89°, and a small dip in frequency
below 78° (Supplemental Fig. S2B). As designed, all interactions exhibited τa (Fig. 6) values measuring <20° and τb (Fig. 6) values measuring over 80° (Supplemental Fig. S2C). Although structures beyond these cutoffs could be of biological significance, relaxing the θhv leads to distortion of the T-geometry of the interactions, whereas relaxing the τa and τb values lead to horizontal sliding of the vertical base with respect to the horizontal base (Supplemental Fig. S3); these would no longer be “T-shaped” interactions. Regardless, the narrow distribution of all four geometrical parameters
further validates that T-shaped contacts are a distinct phenomenon and facilitates the design of an automated search algorithm
for the removal of false positives.
values in the range of 4.2–5.0 Å (Fig. 7). However, θhv (Fig. 6) remains relatively constant within the interval, with small peaks at 80° to 81° and 88° to 89°, and a small dip in frequency
below 78° (Supplemental Fig. S2B). As designed, all interactions exhibited τa (Fig. 6) values measuring <20° and τb (Fig. 6) values measuring over 80° (Supplemental Fig. S2C). Although structures beyond these cutoffs could be of biological significance, relaxing the θhv leads to distortion of the T-geometry of the interactions, whereas relaxing the τa and τb values lead to horizontal sliding of the vertical base with respect to the horizontal base (Supplemental Fig. S3); these would no longer be “T-shaped” interactions. Regardless, the narrow distribution of all four geometrical parameters
further validates that T-shaped contacts are a distinct phenomenon and facilitates the design of an automated search algorithm
for the removal of false positives.
Representation of geometrical parameters used for locating T-shaped interactions in RNA crystal structures.
Distribution of T-shaped interactions involving various interacting atoms/groups of the vertical base, with respect to the vertical distance (Å) between ring centers (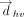 ). As designed, the distance between the interacting atom/group and the center of the horizontal base is always smaller than
). As designed, the distance between the interacting atom/group and the center of the horizontal base is always smaller than 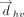 , which is defined as the centroid-to-centroid distance between the interacting ring systems.
, which is defined as the centroid-to-centroid distance between the interacting ring systems.
T-shaped interactions involve the “b” face (59%) slightly more than the “a” face (41%) of the horizontal base (Supplemental Table S5). Further, likely due to tight detection cutoffs (see Materials and Methods section), these interactions involve only one of the two constituent purine rings (six-membered [59%] or five-membered [40%]) rather than bridging both (Supplemental Table S6). However, the involvement of the H (46%) or S (39%) edge in these interactions is clearly more pronounced over the W edge of the vertical base (15%, Supplemental Table S7), because the polar W edge is generally occupied in classical base-pairing interactions (Stombaugh et al. 2009). Further, N3 (45.0%) and C8-H (55.7%) are the most common interacting atoms/groups for A and G in the vertical position, respectively, and C5-H (55.3% for C and 58.7% for U) most commonly interacts for either of the pyrimidines acting as the vertical base (Supplemental Table S8). Alternatively, in terms of the glycosidic orientation, cis-oriented interactions are dominant, and comprise around two-thirds of the total interactions (Supplemental Table S8). Finally, the presence of A as the vertical base (64% of all occurrences) is clearly dominant in T-shaped interactions, although no specific preference for the horizontal base is observed (Supplemental Table S10).
In terms of sequence combinations, around two-thirds of T-shaped contacts involve two purines (Supplemental Table S5). Further, purine–purine combinations span 86 of the 144 theoretically possible combinations (Fig. 4; Supplemental Table S10). Included in this purine–purine subset are examples from all 12 possible combinations of the edges (W/H/S), faces (a/b), and glycosidic orientations (cis/trans, Table 1; Supplemental Table S11). In contrast, only 10% of the contacts involve a purine at the horizontal position and a pyrimidine at the vertical position (Supplemental Table S5). As a result, these interactions span only 51 of the 144 theoretically possible purine–pyrimidine combinations and only nine of the 12 possible combinations of the edges, face, and glycosidic orientations (Supplemental Tables S12, S13). An additional 16% of the contacts involve a pyrimidine at the horizontal position and a purine at the vertical position (Supplemental Table S5). These interactions span 37 of the 48 theoretically possible pyrimidine–purine combinations and involve all 12 possible edge, face, and glycosidic combinations (Supplemental Tables S12, S13). Finally, the pyrimidine–pyrimidine interactions contribute only ∼9% of the total contacts (Supplemental Table S5), although they span 25 of the 48 theoretically possible pyrimidine–pyrimidine combinations and involve nine of the 12 possible edge, face, and glycosidic combinations (Supplemental Tables S12, S13). Overall, in terms of the theoretically possible geometrical combinations, crystal structure analysis led to the identification of examples of 199 out of 384 possible geometries (Fig. 4; Supplemental Table S13). Among these, A(6)-G(C8-H) bH cis is the most common combination, which along with A(5)-A(N3) aS cis and A(5)-G(N3) bS trans, comprises a quarter of all interactions (Supplemental Table S14). Further, 39 examples were identified for T-shaped interactions involving modified bases (37 involving 1-methyladenine and two involving pseudouridine) (Supplemental Tables S15, S16). It is unclear whether the conformations that are not represented are practically unlikely, or in some cases essentially impossible; the data set is simply too small to be expected to encompass all possibilities, and this exercise could be repeated after a suitable interval to determine whether these missing conformations are observed as more structures are deposited.
One-third of the T-shaped interactions occur between consecutive nucleotides, whereas the remaining two-third involve nonconsecutive nucleotides (Supplemental Table S17). These distant interactions occur between bases present in varied RNA structural elements, ranging from bases present within a loop (intraloop interactions, Supplemental Fig. S4A,B), on opposing strands of helical regions (intrahelical interactions, Supplemental Fig. S4C–F), in two different helices (interhelical interactions, Supplemental Fig. S5A–C) or two different loops (interloop interactions, Supplemental Fig. S5D) of an RNA domain. Additionally, T-shaped interactions participate in inter-domain contacts within complex RNA structures (Supplemental Fig. S6). Furthermore, the constituent bases of a T-shaped interaction are involved in highly variable parallel stacking contacts with the surrounding bases, ranging from no participation of either horizontal or vertical base in parallel stacking (Supplemental Figs. S4D, S5A,B), participation of only the horizontal base in parallel stacking (Supplemental Figs. S4C, S6B), participation of only the vertical base in parallel stacking from one (Supplemental Figs. S4A,E,F, S5C,D, S6F) or both faces (Supplemental Fig. S6B) and simultaneous participation of horizontal and vertical bases in parallel stacking (Supplemental Fig. S6A,C–E). However, despite their likely extremely important functional role (vide infra), the frequency of inter-RNA T-shaped interactions is statistically less significant (Supplemental Tables S17, S18).
Statistical analysis of nonredundant interactions
To further solidify our analysis, we considered only one representative from interactions that occur in multiple structures, but share the same nucleotide sequence, face, edge, interacting atom, ring, and glycosidic orientation. This revealed that there are 922 distinct interactions, each located in a different macromolecular context. The distribution of faces (54.2% with “b” face) and interacting ring identity of the horizontal base (six-membered in 63.5% interactions), interaction edge of the vertical base (Hoogsteen edge in 55.9% of cases), interacting atom of the vertical base (N7 [27.7%] for A, N7 [45.1%] for G, C5 for C and U [55.3% for C and 58.7% for U]), glycosidic interaction (cis in 73.2% instances), and identity of the vertical (47.2% A) and horizontal base (36.0% A) are similar to the statistics of all T-shaped interactions (vide supra) in the data set (Supplemental Tables S5–S10). However, although nonconsecutive interactions dominate the total data set, the nonredundant interactions more commonly involve T-shaped contact between consecutive residues (Supplemental Table S17). Regardless, the fact that different subsets of interactions return similar results points toward the robustness of the derived conclusions.
Annotation of T-shaped contacts in RNA structures
As previously suggested for base pairs (Leontis and Westhof 2001) and stacking interactions (Jhunjhunwala et al. 2021), the annotation of T-shaped interactions in pictorial representations of RNA structures or substructures would help better
visually recognize the noncovalent interactions in RNA motifs. In this context, we propose a symbolic formalism for the annotation
of T-shaped interactions that combines the features of Leontis and Westhof (2001) base-pairing annotations and Jhunjhunwala et al. (2021) annotations of parallel stacking interactions. Specifically, we propose that the edge of the vertical base should be specified
using the symbols used for specifying the edges of nucleobases in pairing interactions (i.e., circle for the W edge, square
for the H edge and triangle for the S edge). However, in line with the symbolic annotation of base stacks (St-Onge et al. 2007), the interacting face of the horizontal base can be designated using an arrowhead ( or
or  ), which points away from the horizontal base when it uses its “b” face, and points toward the horizontal base when it uses
its “a” face for the T-shaped interaction. The arrow can then be joined to the edge annotation of the vertical base by a dash
(–) to complete the assignment of a T-shaped interaction. Further, these symbols can be used in their filled and hollow forms
to specify cis and trans glycosidic orientations, respectively. For instance, both symbols “A
), which points away from the horizontal base when it uses its “b” face, and points toward the horizontal base when it uses
its “a” face for the T-shaped interaction. The arrow can then be joined to the edge annotation of the vertical base by a dash
(–) to complete the assignment of a T-shaped interaction. Further, these symbols can be used in their filled and hollow forms
to specify cis and trans glycosidic orientations, respectively. For instance, both symbols “A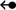 G” and “A
G” and “A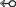 G” would represent T-shaped contacts involving the interaction of horizontal A through its “a” face and vertical G through
its W edge, albeit with cis and trans glycosidic orientations, respectively. Symbols for all 12 families of T-shaped interactions are provided in Table 1.
G” would represent T-shaped contacts involving the interaction of horizontal A through its “a” face and vertical G through
its W edge, albeit with cis and trans glycosidic orientations, respectively. Symbols for all 12 families of T-shaped interactions are provided in Table 1.
When a horizontal purine is involved in a T-shaped interaction, the above annotations can be further enriched by providing information on the identity of the interacting purine ring of the horizontal base (“5,” “6,” or “56”). Further, the interacting atom of the participating edge of the vertical base can be specified to elaborate the T-shaped assignment. To illustrate how these symbols can denote T-shaped interactions, we present examples from select geometric families obtained from high (better than or equal to 2.5 Å) resolution crystal structures (Fig. 8).
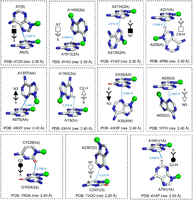
Illustration of the symbolic representation of edge-face nomenclature for T-shaped interactions, using examples from different geometrical families (Table 1). PDB code, crystallographic resolution, and interaction code (horizontal base–vertical base) are provided for each interaction. The interacting atom of the horizontal base and the participating edge of the vertical base are indicated for each annotation. The distance between the interacting atom (heavy atom for the C-H group) of the vertical base and the center of the interacting ring of the horizontal base is provided. Although in some cases, one atom belonging to the interacting edge may visually appear closer than the other atom of the same edge, the interacting atom is identified using a quantitative method according to the algorithm described in the Materials and Methods section. Note that including the information on the ring identity of the horizontal base and the interacting atom/group of the vertical base is optional and can be removed in ambiguous cases.
Structural context of the occurrence of selected T-shaped interactions in RNA structures
To aid in the visual comprehension of how the proposed classification scheme can be used to enrich our understanding of T-shaped
interactions in RNA 3D structures, we annotate these interactions in specific RNA motifs. Specifically, we used examples from
a wide range of RNA structures (Fig. 9). For example, we annotate the bS trans (i.e., 6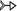 C2-H) T-shaped interaction between the six-membered ring of A975 belonging to helix 31 and the C2-H group of A1357 belonging
to helix 43, the aS cis (i.e., 5
C2-H) T-shaped interaction between the six-membered ring of A975 belonging to helix 31 and the C2-H group of A1357 belonging
to helix 43, the aS cis (i.e., 5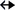 C2-H) interaction between the five-membered ring of A1503 from the single-stranded linker region connecting helices h44 and
h45 and the C2-H group of A1531 from helix 44, and aS cis (6
C2-H) interaction between the five-membered ring of A1503 from the single-stranded linker region connecting helices h44 and
h45 and the C2-H group of A1531 from helix 44, and aS cis (6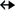 N3) intraloop T-shaped stacking interaction of the six-membered ring of G275 with N3 of A274 of 16S rRNA (PDB code: 4YBB, Fig. 9A). Further, the A231–A259 “trans” arrangement is observed in the bH (i.e., 5
N3) intraloop T-shaped stacking interaction of the six-membered ring of G275 with N3 of A274 of 16S rRNA (PDB code: 4YBB, Fig. 9A). Further, the A231–A259 “trans” arrangement is observed in the bH (i.e., 5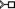 C8-H) interaction present in the three-way junction of the lariat capping ribozyme (4P95, Fig. 9B). However, the “consecutive” A6–A7 aH interaction in an ssRNA acquires a “cis” (5
C8-H) interaction present in the three-way junction of the lariat capping ribozyme (4P95, Fig. 9B). However, the “consecutive” A6–A7 aH interaction in an ssRNA acquires a “cis” (5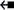 N7) arrangement (4TZO, Fig. 9C). Similarly, the A2133–A2258 aS trans, U10–A532 aH trans loop–loop interactions as well as the A1811–G1814 bH cis intraloop interactions with the crystal structure of 50S ribosomal subunit of Haloarcula Marismortui, are annotated as 6
N7) arrangement (4TZO, Fig. 9C). Similarly, the A2133–A2258 aS trans, U10–A532 aH trans loop–loop interactions as well as the A1811–G1814 bH cis intraloop interactions with the crystal structure of 50S ribosomal subunit of Haloarcula Marismortui, are annotated as 6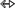 N3, 6
N3, 6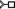 N7, and 6
N7, and 6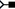 C8, respectively (1YHQ, Fig. 9D).
C8, respectively (1YHQ, Fig. 9D).
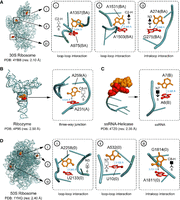
Sample T-shaped interactions from various high-resolution PDBs (PDB code and crystallographic resolution are provided for each source crystal structure), indicating the identity, residue number and chain (in parentheses) of each participating nucleotide. Each T-shaped interaction is indicated by a symbolic representation. The interacting atom of the horizontal base and the participating edge of the vertical base are indicated for each annotation. The distance between the interacting atom (heavy atom for the C-H group) of the vertical base and the center of the interacting ring of the horizontal base is provided. Although in some cases, one atom belonging to the interacting edge may visually appear closer than the other atom of the same edge, the interacting atom is identified using a quantitative method according to the algorithm described in the Materials and Methods section. Note that including the information on the ring identity of the horizontal base and the interacting atom/group of the vertical base is optional and can be removed in ambiguous cases.
In addition to T-shaped contacts between bases belonging to the same RNA sequence, our study identified a T-shaped contact
between two different RNA elements (16S rRNA and 23S rRNA) in the crystal structure of Thermus thermophilus 70S ribosomal complex (Fig. 10, 7RQD). The bH cis (i.e., 5 C8) geometry of this A1493(1a)–A1913(1A) T-shaped interaction is like the A1811–G1814 geometry in the ribosome (Fig. 9D). Together with intra-RNA T-shaped contacts, the existence of inter-RNA T-shaped contacts suggests that these interactions may play a role in noncovalent structural organization within RNA, as
well as between two RNA structures.
C8) geometry of this A1493(1a)–A1913(1A) T-shaped interaction is like the A1811–G1814 geometry in the ribosome (Fig. 9D). Together with intra-RNA T-shaped contacts, the existence of inter-RNA T-shaped contacts suggests that these interactions may play a role in noncovalent structural organization within RNA, as
well as between two RNA structures.
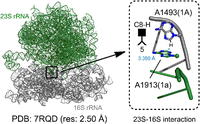
A representative inter-RNA T-shaped interaction in the crystal structure of 70S ribosome of Thermus thermophilus. The distance between the interacting atom (heavy atom for the C-H group) of the vertical base and the center of the interacting ring of the horizontal base is provided. Although one atom belonging to the interacting edge may visually appear closer than the other atom of the same edge, the interacting atom is identified using a quantitative method according to the algorithm described in the Materials and Methods section. Note that including the information on the ring identity of the horizontal base and the interacting atom/group of the vertical base is optional and can be removed in ambiguous cases.
We further aligned the sequences of all rRNA structures considered in the data set to understand if the interactions are conserved in structures both within and across species. Our results suggest that specific T-shaped interactions are conserved across many structures within the same organism, as well as between multiple organisms (Supplemental Tables S19, S20). For example, the G573–G571 (G(5)–G(N3) bS trans) interaction occurs in 23S rRNA structures belonging to three different organisms (Supplemental Table S20). Similarly, the A975–A1357 (A(5)–A(C2-H) aS cis) interaction of 16S rRNA is also present in structures from more than one organism. However, in some cases, the T-shaped interaction is retained even on base substitutions. For example, the T-shaped interaction of 23S rRNA between 1755 and 1758 is retained even on substitution of A-G with A-U (Supplemental Table S20). Overall, this points toward the likely conservation of specific T-shaped interactions in RNA structures.
Conclusion
RNA structures exploit a variety of noncovalent interactions to acquire complex 3D shapes that dictate their functions. In addition to strong edge-to-edge hydrogen bonding interactions among nucleobases (Halder and Bhattacharyya 2013), it is now recognized that RNA molecules use a plethora of additional weak stabilizing strategies that help achieve compact 3D structures and stabilize specific structural motifs. Analogous to the perpendicular π stacking between aromatic amino acids and nucleobases in DNA:protein and RNA:protein complexes (Wilson et al. 2014, 2016, 2021), T-shaped interactions between two nucleobases are also expected to play a modest stabilizing role in RNA. In the present work, we devised an automated tool for the identification of hitherto unknown T-shaped contacts between nucleobases in RNA structures. We further developed and applied a comprehensive classification scheme to unambiguously categorize these contacts in RNA structures and proposed a symbolic representation to annotate T-shaped interactions in secondary structure diagrams and 3D structural representations of RNA. Our results reveal that in analogy to the greater prevalence of parallel stacking over T-shaped interactions at the RNA:protein interface (Wilson et al. 2021), T-shaped interactions are far less common than parallel stacking interactions. Further, with the unusual conformation that they demand, it is reasonable that they would be more common in highly folded RNA and may not be necessarily important in most classical double-stranded RNA or coding regions of mRNA. Furthermore, these interactions are expected to be of similar strength to other relatively weak, but important intramolecular interactions, including lone pair-π stacking between nucleobases and sugar (Chawla et al. 2017) or water (Kalra et al. 2020), anion−π stacking (Chawla et al. 2022), and OH−π stacking between nucleobases and water (Kalra et al. 2020) that have been recently characterized in the 3D RNA structures. Consequently, it is not possible to comprehensively delineate the structural determinants of RNA structure without considering T-shaped interactions, and this requires an identification of these interactions.
Overall, our study adds to the list of previously established noncovalent effects, including but not limited to, base pairing, parallel base stacking, lone pair-π interactions between ribose and nucleobases, lone pair-π, and OH–π interactions between water and nucleobases, base-wedged and base-intercalated stacking, as well as base–phosphate interactions, that help organize RNA structures.
MATERIALS AND METHODS
Structural data set
A data set containing all RNA crystal structures deposited in the Protein Data Bank (PDB) through May 28, 2022 was used to identify the T-shaped interactions. The structures were searched from the PDB by setting the “Polymer Entity Type” to “RNA” and by choosing “X-ray” in the “Method” tab. Further, in agreement with previous studies, the refinement resolution of the crystal structures was set to ≤3.5 Å (Chawla et al. 2014, 2015; Wilson et al. 2014). A total of 3280 structures were obtained from this search, which were further filtered using the CD-HIT suite (Huang et al. 2010), to retain only a single representative from clusters of similar structures with >80% similarity. This procedure led to the final 2286 crystal structures (Supplemental Table S1). However, considering that some of the identified contacts may not be unambiguously confirmed in low resolution crystal structures, examples of T-shaped interactions are provided from better than or equal to 2.5 Å resolution, in accordance with a previous structural study on RNA-relevant contacts (Zirbel and Auffinger 2022).
Identification of T-shaped interactions
In analogy with the “Stackdetect” algorithm previously developed for detecting base–base stacking interactions (Jhunjhunwala et al. 2021), a python script was developed for automated detection and classification of T-shaped interactions in RNA crystal structures. In the first step, the Cartesian coordinates of atoms forming the five-membered ring skeleton of a purine (i.e., C4, C5, N7, C8, and N9 atoms) and the six-membered ring of a purine or a pyrimidine (N1, C2, N3, C4, C5, and C6), and C1′ of the sugar moity were extracted, and the coordinates of the centroid of each ring was determined (xc, yc, zc).
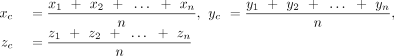 Here, (x1, y1, z1), (x2, y2, z2), (x3, y3, z3)…… (xn, yn, zn) are the Cartesian coordinates of atoms of each five-membered purine ring or six-membered purine or pyrimidine ring. The
centroid coordinates were then used to determine the position vector (
Here, (x1, y1, z1), (x2, y2, z2), (x3, y3, z3)…… (xn, yn, zn) are the Cartesian coordinates of atoms of each five-membered purine ring or six-membered purine or pyrimidine ring. The
centroid coordinates were then used to determine the position vector (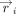 ) of each (ith) ring atom from the ring centroid. Specifically:
) of each (ith) ring atom from the ring centroid. Specifically: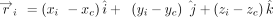 Here, (xi, yi, zi) denote the Cartesian coordinates of the atom i. Subsequently, the distance vector (
Here, (xi, yi, zi) denote the Cartesian coordinates of the atom i. Subsequently, the distance vector (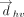 ) that connects the centroids of vertical ring (denoted as ring “v”) and the horizontal ring (denoted as ring “h”) that participate
in a T-shaped interaction was determined using the centroid coordinates of the rings (xhc, yhc, zhc) and (xvc, yvc, zvc) (Fig. 6A). Specifically:
) that connects the centroids of vertical ring (denoted as ring “v”) and the horizontal ring (denoted as ring “h”) that participate
in a T-shaped interaction was determined using the centroid coordinates of the rings (xhc, yhc, zhc) and (xvc, yvc, zvc) (Fig. 6A). Specifically: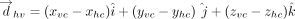 For each interacting ring, a mean plane was then defined using the two orthogonal vectors
For each interacting ring, a mean plane was then defined using the two orthogonal vectors 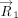 and
and 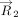 using the Cremer and Pople method (Cremer and Pople 1975).
using the Cremer and Pople method (Cremer and Pople 1975). The vectors normal to the mean plane vectors (
The vectors normal to the mean plane vectors (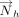 and
and 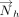 ) of each ring were then determined as the cross product of
) of each ring were then determined as the cross product of 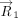 and
and 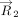 .
.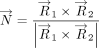 The inverse cosine of the dot product of the normals
The inverse cosine of the dot product of the normals 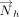 and
and 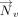 of the interacting rings h and v was used to calculate θhv, the tilt angle between the rings (Fig. 6B; Cremer and Pople 1975). Specifically:
of the interacting rings h and v was used to calculate θhv, the tilt angle between the rings (Fig. 6B; Cremer and Pople 1975). Specifically: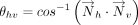 Further, the angle between
Further, the angle between 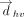 and
and 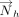 or
or 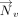 (i.e., τh or τv, Fig. 6C) was calculated for each interaction.
(i.e., τh or τv, Fig. 6C) was calculated for each interaction.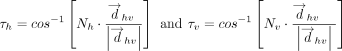 The values of τh and τv determine the relative horizontal displacement of the interacting rings.
The values of τh and τv determine the relative horizontal displacement of the interacting rings.
In line with the previous studies on T-shaped interactions between DNA nucleobases and aromatic amino acids that contain either
a single or fused carbocyclic or heterocyclic ring system (Wilson et al. 2014), the following criteria were used for locating a T-shaped interaction between the two interacting rings: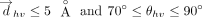
However, the ranges for τh and τv needed to be determined empirically through preliminary trials. The best cutoffs for these interactions to retain a T-shaped
conformation were found to be: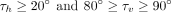
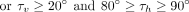
For purines, these parameters were calculated separately for both five-membered and six-membered rings, and the identity of the interacting ring was assigned accordingly (vide infra).
Classification of T-shaped interactions
For each T-shaped interaction, the vertical base can interact with the horizontal base through one of its three edges—the
Watson–Crick (W) edge, Hoogsteen (H) edge, or the Sugar (S) edge (Fig. 2A; Leontis and Westhof 2001). The interacting edge of the vertical base was computationally identified by calculating the coordinates of the center of
each edge of the base by averaging the coordinates of all donor and acceptor atoms that constitute the edge (Supplemental Fig. S7; Supplemental Table S14; Leontis and Westhof 2001).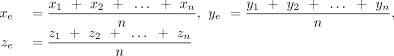 Here, (x1, y1, z1), (x2, y2, z2), (x3, y3, z3)…… (xn, yn, zn) are the Cartesian coordinates of the selected edge atoms and xe, ye, ze are the coordinates of the edge center (Supplemental Table S15). Subsequently, the distance between the centroid of the horizontal ring and the center of each edge of the vertical base
was calculated. The edge closest to the centroid of the interacting face of the horizontal ring was accordingly assigned as
the “interacting edge” for the purposes of classifying the T-shaped interaction. Within the interacting edge of the vertical
base, the atom closest to the horizontal ring was identified by calculating the distances between the centroid of the horizontal
ring and each atom that constitutes the edge (Supplemental Table S24).
Here, (x1, y1, z1), (x2, y2, z2), (x3, y3, z3)…… (xn, yn, zn) are the Cartesian coordinates of the selected edge atoms and xe, ye, ze are the coordinates of the edge center (Supplemental Table S15). Subsequently, the distance between the centroid of the horizontal ring and the center of each edge of the vertical base
was calculated. The edge closest to the centroid of the interacting face of the horizontal ring was accordingly assigned as
the “interacting edge” for the purposes of classifying the T-shaped interaction. Within the interacting edge of the vertical
base, the atom closest to the horizontal ring was identified by calculating the distances between the centroid of the horizontal
ring and each atom that constitutes the edge (Supplemental Table S24).
The identity of the interacting face of the horizontal ring was then established. Specifically, the face was designated as
alpha (a) when the standard nucleobase atomic numbering of the ring from lowest numbered atom to the subsequent higher numbered
atoms by shortest route progresses in a “clockwise” manner, when the base is viewed from the top of the interacting face (Fig. 2B; Rose et al. 1980). Alternatively, if the progression occurs in a “counterclockwise” direction, the face is named as beta (b) (Fig. 2B). Although by convention the face designation of a purine should be derived from the numbering of its six-membered ring,
irrespective of whether the six-membered or the five-membered ring interacts with the other base (Rose et al. 1980), we considered the five-membered ring for face designation in purines to preserve the similarity in face labels for purines
and pyrimidines, and to keep the face designation in conformity with previous studies on parallel stacking interactions in
RNA (Fig. 2B; St-Onge et al. 2007; Jhunjhunwala et al. 2021). Computationally, the face assignment is done using the direction of the normal to the mean plane vector of the horizontal
base ( ). As the mean plane is defined by the right-handed rotation of the six-membered ring of nucleobases (from N1 to N6, Fig. 6), the direction of the
). As the mean plane is defined by the right-handed rotation of the six-membered ring of nucleobases (from N1 to N6, Fig. 6), the direction of the 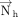 is revered in a purine compared to a pyrimidine. Therefore, analogous to studies on parallel stacking (Major and Thibault 2007; Jhunjhunwala et al. 2021), we defined a new normal vector
is revered in a purine compared to a pyrimidine. Therefore, analogous to studies on parallel stacking (Major and Thibault 2007; Jhunjhunwala et al. 2021), we defined a new normal vector 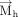 that reverses its direction compared to
that reverses its direction compared to 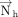 in case of purines. Specifically,
in case of purines. Specifically,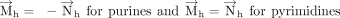 This assignment ensures that the face assignment vector
This assignment ensures that the face assignment vector 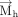 extends from the b face, in both purines and pyrimidines. The face is designated as “a” if the angle between
extends from the b face, in both purines and pyrimidines. The face is designated as “a” if the angle between 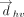 and
and 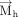 is greater than 90°, otherwise it is assigned as “b.”
is greater than 90°, otherwise it is assigned as “b.”
The mutual glycosidic orientation of each T-shaped interaction was assigned by calculating the angle σhv (∠(C1′)h–Xh–Xv–(C1′)v), where (C1′)h and (C1′)v represent the C1′ atoms of the nucleotides that include the horizontal and the vertical ring, respectively, and Xh and Xv are the centroids of the respective base rings that connect to the sugar through the glycosidic bond (five-membered ring in case of purines and six-membered ring in case of pyrimidines, Fig. 2C).
The identity of the interacting ring of the horizontal base was determined by calculating the distance between the centroid
of the vertical base and each ring (five- or six-membered ring of purines and six-membered ring of pyrimidines) of the horizontal
base. For example, if the geometrical parameters (i.e., 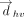 , θhv, τh, and τv) of the center of the vertical base and the center of the five-membered ring of the horizontal purine fall under the cutoff
value, the interaction is denoted as “5” (Fig. 2D). Alternatively, if the parameters of the six-membered purine ring fall within the cutoff, the ring topology is assigned
as “6” (Fig. 2D). However, if the parameters for both five-membered and six-membered purine rings fall with the cutoff, the T-shaped interaction
is noted as “56” (Fig. 2D).
, θhv, τh, and τv) of the center of the vertical base and the center of the five-membered ring of the horizontal purine fall under the cutoff
value, the interaction is denoted as “5” (Fig. 2D). Alternatively, if the parameters of the six-membered purine ring fall within the cutoff, the ring topology is assigned
as “6” (Fig. 2D). However, if the parameters for both five-membered and six-membered purine rings fall with the cutoff, the T-shaped interaction
is noted as “56” (Fig. 2D).
Identification of consecutive, nonconsecutive, and inter-RNA T-shaped interactions
Each T-shaped interaction was identified as consecutive, nonconsecutive, or inter-RNA, based on the absolute value of the difference between the nucleotide residue numbers in the PDB file, and whether the interacting nucleotides belong to the same RNA entity or to two different interacting RNA strands. A T-shaped interaction between nucleotides belonging to the same chain in the PDB file is identified as consecutive, if the difference between their residue numbers equals 1, that is, they are immediately adjacent; otherwise, the interaction is identified as nonconsecutive. However, if the interacting nucleotides belong to two different RNA present in the same PDB file, the interaction is classified as inter-RNA (Fig. 5).
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
This research project was supported by the Department of Science and Technology (DST), University Grants Commission (UGC). P.S. thanks the Department of Science and Technology (DST) and University Grants Commission (UGC), New Delhi, for financial support through the DST INSPIRE (IFA14-CH162) and the UGC FRP (F.4−5[176-FRP/2015(BSR)]) programs, respectively. J.F.T. thanks the Natural Sciences and Engineering Research Council of Canada. Z.A. thanks UGC for a Senior Research Fellowship.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079486.122.
- Received October 22, 2022.
- Accepted April 25, 2023.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is an editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Zakir Ali is the first author of the paper “Occurrence and classification of T-shaped interactions between nucleobases in RNA structures.” Zakir is currently working as a senior research fellow in the Department of Chemistry, Panjab University, Chandigarh, India. His research is focused on computational studies on noncovalent interactions in RNA structures.
What are the major results described in your paper and how do they impact this branch of the field?
Our work includes identification and classification of T-shaped interactions involving nucleobases in RNA structures. We further provide an annotation scheme for this interaction. Our results are expected to lead to further insights in the RNA architecture and functioning.
What led you to study RNA or this aspect of RNA science?
The complexity of RNA structures always fascinated me and led to my research in the field of RNA.
What are some of the landmark moments that provoked your interest in science or your development as a scientist?
From a very young age, I always wondered how exactly things unfold in nature, specifically chemically and biologically.
If you were able to give one piece of advice to your younger self, what would that be?
I would have tried to develop scientific inquisitiveness much earlier in my life. Further, I would have paid more attention to understanding things than memorizing facts.
What are your subsequent near- or long-term career plans?
Both my near-term and long-term career plans will be focused toward digging more into the details of RNA structures and functioning.








