
Starting and stopping RNA polymerase III transcription on single-stranded DNA oligonucleotides
- Lodoe Lama1,2 and
- Kevin Ryan1,2,3
- 1Department of Chemistry and Biochemistry, The City College of New York, New York, New York 10031, USA
- 2Biochemistry and Chemistry Ph.D. Programs, The City University of New York Graduate Center, New York, New York 10016, USA
- 3Chemistry Ph.D. Programs, The City University of New York Graduate Center, New York, New York 10016, USA
- Corresponding author: kryan{at}ccny.cuny.edu
Abstract
Circularized single-stranded DNA oligonucleotides, or coligos, show promise as promoter-independent RNA polymerase III (Pol III) transcription templates for generating small RNA in human cells. Using a modified small RNA-seq method, we studied the sequence and secondary structure characteristics that determine Pol III initiation and termination on six coligo templates. The coligos each consisted of an imperfectly base-paired stem flanked by one larger and one smaller loop and were unrelated in sequence. Small RNA-seq data from Pol III coligo transcripts revealed a strong preference for initiating transcription within a 5-nucleotide (nt) window spanning the stem-larger loop junction (loop size 11–24 nt). Transcription in all cases proceeded into the stem rather than into the larger loop, indicating the junction is a site-specific, secondary structure-based Pol III transcription initiator. On average, 81% of sequencing reads showed initiation within this 5 nt junction region, with a template start site nucleotide preference of C > T >> A > G, and a requirement for a template purine at Tss-1. Termination was less precise than initiation and occurred in the larger loop at the same end of the stem where transcription initiated. Termination efficiency was on average 82% and was distributed among the first 11 single-stranded larger loop nt following the stem. The size heterogeneity of Pol III coligo transcripts is thus mainly due to 3′ end heterogeneity, whereas the RNA 5′ ends were more predictable and homogeneous. Transcription termination did not require an oligo dA template sequence, indicating that termination in this context may be mechanistically different than Pol III's normal gene-context termination. A stepwise model for coligo transcription by Pol III is proposed.
Keywords
INTRODUCTION
We are investigating circularized synthetic DNA oligonucleotides, or coligos, as potential vectors for engineering the expression of small RNA in human cells. As expression vectors, coligos are unique in that they contain only the template strand and lack a transcription promoter sequence, instead appearing to rely on sequence- and secondary structure-triggered transcription initiation and termination (Seidl and Ryan 2011; Seidl et al. 2013; Lama et al. 2014; Patel et al. 2016). Coligos lack 5′ and 3′ ends, a feature that reduces susceptibility to nucleases and further distinguishes them structurally from other promoter-independent eukaryotic transcription templates like the 3′ overhang and oligo dC-tailed templates (Dedrick and Chamberlin 1985; Campbell and Setzer 1992; Bardeleben et al. 1994; Schroder et al. 2003). To date, we have designed coligos to encode transcripts resembling pre-miRNA with the aim that, when generated in cells, the transcripts may enter the natural miRNA maturation pathway and lead to mature miRNA or siRNA. To encode this hairpin RNA motif, the coligos contain an imperfectly base-paired stem flanked by one larger and one smaller loop, and thus resemble asymmetric DNA dumbbells (Erie et al. 1987). Pre-miRNA-encoding coligos generally produce hairpin transcripts with modest to very good end heterogeneity, as judged by electrophoresis and preliminary cDNA sequencing analyses (Seidl et al. 2013; Lama et al. 2014; Patel et al. 2016). Most coligo-dependent transcripts result from a single circumtranscription around the template, with in most cases negligible rolling circle transcription. This behavior distinguishes them from the DNA nanocircles that have been observed to undergo promoter-independent rolling circle transcription by bacterial and bacteriophage RNA polymerases (Daubendiek et al. 1995; Ohmichi et al. 2002).
We have found evidence that RNA polymerase III (Pol III) is responsible for transcribing coligo DNA templates in human tissue culture cells and their whole cell and nuclear extracts (Seidl et al. 2013). Experiments with Flag-tagged Pol III immunoprecipitated from HEK293 cells have further implicated Pol III as the main coligo-transcribing polymerase (Lama et al. 2014). The sizes of RNA templated from three different coligos transcribed in cells, as visualized by Northern blotting, agreed well with those from the same coligos transcribed by Pol III in cell extracts (Seidl et al. 2013). This observation has facilitated progress in understanding coligo template behavior by showing that Pol III transcription in vitro largely predicts the coligo-dependent transcripts made by Pol III in cells, though cellular exonucleases may trim the primary transcripts’ ends. During Flag-Pol III immunoprecipitation, nucleases are removed, along with the abundant small RNA in the size range typical for coligo transcripts, for example, 50–150 nt. Removing nucleases ensures that the transcripts observed are primary transcripts, and removing endogenous RNA facilitates cDNA library preparation (Lama et al. 2019).
To exploit the usefulness of coligos as small RNA delivery templates, whether as research tool or as potential small therapeutic RNA delivery vector, it will be necessary to learn how DNA coligo sequence and secondary structure determine Pol III transcription initiation and termination sites. Toward that end, we recently developed a modified small RNA-seq protocol, termed coligo-seq (Lama et al. 2019). Coligo-seq uses Illumina sequencing of a cDNA library made from coligo in vitro transcripts, and works well even when the RNA contains a strong secondary structure that would otherwise interfere with the 5′ sequencing adaptor ligation used in standard small RNA-seq library preparation protocols. Coligo-seq yields whole transcript sequences and now enables us to analyze transcript heterogeneity by assessing sequencing reads from all transcripts, be they rare or abundant. The transcripts can then be mapped to the coligo sequence and predicted secondary structure to understand how these features correlate with Pol III initiation and termination.
Here we have carried out in vitro Pol III transcription using five new coligo templates designed to produce imperfectly base-paired RNA hairpins in the 45 to 90 nt range, and used coligo-seq to sequence the transcript cDNA to learn precisely the Pol III initiation and termination sites on the coligos. We map the transcript reads onto the predicted coligo secondary structures and, in combination with data from a sixth, previously published coligo (Lama et al. 2019), describe the determinants for initiating and terminating RNA Pol III transcription on coligos that encode RNA hairpins. Beyond pre-miRNA, imperfectly base-paired hairpins constitute a fundamental unit of RNA secondary structure from viruses to eukaryotes, and our findings provide useful guidance for harnessing cellular Pol III to convert chemically synthesized DNA oligonucleotides into hairpin RNA of interest. Our findings also raise interesting questions on what DNA template features trigger Pol III initiation and termination outside of the context of natural promoter-based Pol III gene transcription.
RESULTS AND DISCUSSION
Hairpin-encoding coligos
To investigate Pol III coligo transcription behavior, we studied five coligos encoding RNA hairpins with imperfectly base-paired stems but otherwise unrelated in sequence. While these derive mainly from pre-miRNA genes, they represent the ubiquitous hairpin structural motif found in RNA from all forms of life and viruses. Imperfectly base-paired hairpins of similar size can be found, for example, in small RNA such as spliceosomal RNA, tRNA and ribozymes, and within larger RNAs such as viral genomic RNA (Kuo and Masters 2013) and ribosomal RNA. RNA stem–loop motifs are also the targets of some RNA-binding proteins (Dominguez et al. 2018). Coligos show promise for generating sequence-diverse RNA fragments containing this motif by the action of Pol III in mammalian cells (Seidl et al. 2013).
The coligos studied here range in size from 53 to 89 nt. The sequence of coligo 122 (89 nt) was adapted from the cDNA of human pre-miRNA 122 (hsa-mir-122) (Griffiths-Jones et al. 2006). This coligo has previously been found to be transcribed by Pol III into an ∼83 nt RNA in cells and in vitro (Seidl et al. 2013). Coligo 15a-3 (53 nt) was adapted from the cDNA of hsa-mir-15a gene. The coligo mut2B (61 nt) sequence was adapted from an anti-luciferase shRNA (Paddison et al. 2002). Coligo 221 (67 nt) was adapted from hsa-mir-221 cDNA, and coligo 143-2 (56 nt) from hsa-mir-143 cDNA. The coligos were transcribed in vitro by immunoprecipitated Flag-tagged HEK293 Pol III. Terminated transcripts have previously been observed to be released from the Pol III beads (Lama et al. 2014). Coligo-seq (Lama et al. 2019) was carried out on the main cluster of transcripts resulting from single circumtranscription events (Fig. 1A, n = 1, open circular arrows) as determined by the gels in Figures 1 and 2. The RNA that was excised for sequencing is indicated on the gels by dotted line rectangles. In all coligos observed thus far, low levels of termination read through leads to successively diminishing percentages of tandem multimer transcripts (n > 1, spiral arrows). The striking periodicity of the transcript gel patterns is consistent with common 5′ and 3′ end sequence distributions for all major and minor coligo transcripts. Thus, sequencing the n = 1 single-round transcripts yields information on all coligo-dependent RNA resulting from Pol III transcription. During analysis we added coligo-seq data from a sixth coligo, Dcr3 (Lama et al. 2019), based on the cDNA of hsa-mir-19a. Dcr3 was used as a test case during the development of coligo-seq, but has not otherwise been analyzed in detail.

RNA Pol III initiation and termination sites on coligo 15a-3. (A) Denaturing polyacrylamide gel showing in vitro Pol III transcripts (IVT) made from coligo 15a-3 and, for reference, coligo 122. The RNA within the dashed line rectangle was cut from the gel for coligo-seq cDNA library preparation. Spiral arrow indicates tandem multimer transcripts (n > 1) resulting from termination read through. The periodic transcript pattern indicates similar initiation and termination sites in n = 2,3,4 multimer transcripts. A line between lanes indicates lanes originally separated on the same gel. RNA was visualized by uniform [α-32P]-UTP incorporation; multimers are overrepresented by multiples of incorporated label. Phosphorimager quantitation is corrected for uniform labeling and rounded off. M, RNA marker. (B) Read length distribution of cDNA library after Illumina sequencing. (C) The 15a-3 Pol III transcription initiation (i.e., start) sites (Tss, green bars) and termination (Term.) sites (red bars) were compiled and mapped to the predicted 15a-3 secondary structure. Read Coverage indicates the percentage of the total reads summarized by the red and green bars. If no number is shown next to a nucleotide, then reads initiating or terminating there were <1% of the total reads. LL, larger loop; SL, smaller loop; Stem, double-stranded region. Black triangle, circligation site. Curved line, possible Pol III termination sequence. Transcription proceeds counterclockwise around the coligo as drawn. (D) Most abundant n = 1 transcript. ppp, 5′ triphosphate.
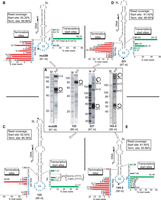
Coligo-seq data summary for Pol III transcription of coligos mut2B, 122, 221, and 143-2. (A) Denaturing polyacrylamide gels showing in vitro transcript profiles. The RNA within the dashed line rectangles was cut from the gel for coligo-seq. Spiral arrows: tandem multimer transcripts. IVT, in vitro transcription. M, RNA marker. A line between lanes indicates lanes originally separated on the same gel. (B) Pol III initiation sites (green bars) and termination sites (red bars) on coligo mut2B from compiled coligo-seq reads mapped onto the sequence illustrated with its predicted secondary structure. (C) Coligo-seq results for coligo 122, as in panel B. A total of 35.99% of transcripts began at the -TTT- sequence with the following 5′ RNA end distributions: 5′rAA (7.27%); 5′rAAA (4.83%); 5′rAAAA (18.91%); and 5′rAAAAA (4.98%). Untemplated rA nt in the latter two transcripts may indicate Pol III slippage during initiation. (D) Coligo-seq results for coligo 221, as in panel B. (E) Coligo-seq results for coligo 143-2, as in panel B.
Pol III coligo transcripts deduced from coligo-seq
The Pol III transcripts made from coligo 15a-3 are representative of this group of coligos. Figure 1A shows the denaturing electrophoresis gel profile of 15a-3 transcripts made in vitro by Flag-Pol III. Transcripts from coligo 122 are also shown for comparison (see also below). As noted above, the periodicity of tandem multimer transcripts, indicated by the spiral arrow, is characteristic of coligo transcription in vitro, and indicative of site-specific initiation and termination. Note that tandem multimer transcripts, which result when termination after one circumtranscription fails to occur, are overrepresented by the uniform labeling method used here. We have found that the n = 2 multimers are, when corrected for label amount, usually about 10% of the n = 1 transcripts, and higher multimers taper off rapidly, though can remain visible on the gel due to the higher multiples of incorporated label (Seidl et al. 2013). The cluster of monomer (n = 1) transcripts spanning 45–55 nt are the most abundant transcripts produced. These were excised from the gel and subjected to coligo-seq library preparation. Illumina sequencing read lengths correlated strongly with the size of the excised transcripts (Fig. 1B).
The predicted secondary structure of coligo 15a-3 is shown in Figure 1C. We mapped the transcription initiation and termination sites from whole transcript reads onto the coligo predicted secondary structure. Green bars mark transcription initiation sites, and red bars mark transcription termination sites. The height of the bar indicates the percentage of coligo-derived reads initiating or terminating at this template nucleotide. Coligo nucleotides that have no color bar were not involved in initiation or termination (i.e., <1% of total reads). While it is difficult to judge transcript heterogeneity from gel data, the coligo-seq data revealed a striking preference for transcription initiation at a single C in the larger loop at the junction with the coligo stem, so that over 83% of the transcripts began with the rG encoded by this position.
The larger loop was also the region where Pol III terminated. Here, though, termination was less precise and was distributed over most of the first ten larger loop nucleotides (Fig. 1C, red bars). This result shows that the n = 1 transcript size range visible in the gel is due to heterogeneity at the 3′ end. The coligo 15a-3 larger loop has a canonical Pol III oligo dA termination signal (Hamada et al. 2000), though due to the coligo's single-stranded nature it is outside the normal gene context of double-stranded (ds) DNA. Our data show that most termination overlaps with this signal, and thus the signal may have caused some termination, but data from other coligos show that oligo dA is not required for Pol III termination (see below). While it is clear that termination occurs in the 15a-3 larger loop, the observation of tandem multimers indicates that a fraction of Pol III (10% for coligo 15a-3, see Table 1) reads through the normal termination sites, producing the periodic pattern of tandem multimer transcripts. Quantification of the tandem multimers (Fig. 1A) shows that they decline rapidly with size, consistent with common 5′ ends and strong but incomplete termination each time the polymerase enters the larger loop. The sequence of the most abundant coligo 15a-3 transcript is shown in Figure 1D. All other coligo-derived monomer transcripts in the sequenced cluster are variations on this main transcript, differing only in end heterogeneity.
Termination efficiency for the six coligos studied
The Pol III transcript gel profiles for the other four coligos are shown in Figure 2A, and the coligo-seq data from the excised transcripts, indicated on the gels by the dotted line rectangles, are shown mapped to the predicted coligo secondary structures in Figure 2B–E. Coligo mut2B is very similar in size and secondary structure to coligo 15a-3. As with 15a-3, transcription initiation on mut2B occurred primarily at a single dC nucleotide near the stem-larger loop junction, though in this case the dC was located on the stem side of the junction. Note that transcription initiation at the junction is site-specific, always proceeding into the stem, and never from the stem into the loop, consistent with our structural prediction of a unidirectional alignment of the coligo within the Pol III active site at initiation (Patel et al. 2016). Also similar to 15a-3, transcript size heterogeneity was due to 3′ end heterogeneity, and termination was distributed among the first ten or eleven larger loop nucleotides. Coligo mut2B has no canonical oligo dA Pol III termination site, yet as observed in 15a-3, termination occurred mainly after one circumtranscription. This observation shows unequivocally that an oligo dA sequence is not required for transcription termination on coligo templates.
Coligo 122 (89 nt) was one of the first coligos studied and gives rise to a nearly homogeneous 83 nt Pol III transcript. Despite the lack of any Pol III termination site, coligo-seq showed that about 76% of termination occurred at the dAGA sequence starting 3 nt into the larger loop (Fig. 2C). All 15 clones from a preliminary 3′ RACE experiment showed 3′ ends at this site as well (Seidl et al. 2013). Thus, coligo 122, which lacks a canonical Pol III termination site, yields transcripts terminating within a very narrow range of larger loop nucleotides. This example demonstrates that moderately precise termination on coligos is possible, but 122 is the exception in this regard among coligos studied thus far. With regard to initiation, coligo 122 has no dC within 5 nt on either side of the stem-larger loop junction where transcription toward the stem would be expected to take place based on the 15a-3 data. Instead, the polymerase used dTs on both sides of the stem larger loop junction, one dT two nt into the larger loop (41.52%), and two in the stem (sum 12.1%). It is known that most multisubunit RNA polymerases (Busby 2021), including Pol III (Zecherle et al. 1996), prefer to start at a template strand pyrimidine. The 122 start site preference appears to be a combination of that principle with Pol III recognition of the coligo stem-larger loop junction as a structure-based initiator. One peculiarity in the 122 coligo-seq data was noted. Between the initiating dT sites at the stem–loop junction there are also two purines, G and A (5′-TTGAT-3′). Approximately 24% of reads were of a length consistent with initiation at G and A, but the reads began with dT (transcript began with rA) rather than dG or dA (Fig. 2C, Tss). We speculate that some additional 122 initiation occurred in the stem T3 sequence and the template repositioned before elongating, that is, underwent slippage (Dieci et al. 1995) within the Pol III active site, adding one or two untemplated adenylates to the 5′ end of the RNA. Another possibility is that the reverse transcriptase, Superscript III, added one or two untemplated nucleotides to the 3′ end of some cDNA during cDNA library construction. Unlike Superscript II, according to the manufacturer, Superscript III has almost no terminal transferase activity under the conditions we used (Lama et al. 2019). Thus, for 122, this explanation is not likely but may explain the much smaller percentage of transcripts in coligo Dcr3 (1.1%–3.5%) (Lama et al. 2019) and mut2B (1.7%), where the 5′ nucleotide did not match the coligo sequence nucleotide. Coligo 122 is the only coligo thus far observed to be transcribed by Pol III with a significant percentage of untemplated nucleotides.
Coligo 221 lacks a dC within 2 nt of the stem-larger loop junction, and coligo-seq showed that Pol III chose two dTs close to the junction to initiate over 78% of transcription events captured by coligo-seq reads (Fig. 2D). A dC 3 nt into the larger loop was used for initiation in only 1.21% of the reads, suggesting that an initiator pyrimidine in the larger loop must be within 2 nt of the stem-larger loop junction to serve as an efficient initiation site. We also note that this dC has a pyrimidine on its 3′ side, which is much less favorable than having a purine at this position (Basu et al. 2014), discussed further below. Termination on 221 was typical in that it was spread over many (nine) nucleotides in the larger loop but, atypically, termination started in the stem just prior to reaching the larger loop. We speculate that the large stem bulge 4 bp from the larger loop, and unique to 221 among the coligos studied here, may have triggered early termination compared to the other coligos, which only have single nt internal loops close to the larger loop. (Some unpaired stem nucleotides are needed for complete circumtranscription [Lama et al. 2014].) The coligo 221 gel profile also shows that, despite lacking an oligo dA terminator in the larger loop, termination was strong compared to the other coligos, as it gave rise to fewer tandem multimers than usual.
In contrast to 221, coligo 143-2, gave rise to greater than normal multimer transcripts, though it has an oligo dA run in the larger loop. Otherwise, it behaved typically, with most transcription (53.4%) initiating at dCs near the stem-larger loop junction, and termination distributed over several larger loop positions.
Initiator determinants: junction sequence and secondary structure
Our coligo-seq results show that the coligo stem-larger loop junction is a secondary structure-based initiator capable of promoting site-specific Pol III transcription. When we examined initiation in the region centered on the junction, spanning 3 nt into the stem and 2 nt into the larger loop, here designated as +3 to −2, with the junction situated between +1 and −1 (Fig. 3A), we found that 53% to 93% of initiation sites (average 81%) fell within these 5 nt positions in the six coligos analyzed by coligo-seq (Fig. 3B). In the region flanking this 5 nt window, initiation was very low, and elsewhere in the coligo it approached zero (Fig. 3C, and Fig. 2). Consistent with what is known for all multisubunit RNA polymerases, the nucleotide identity at the initiation site was also important (Fig. 3D): A dC was preferred, but dT was used if dC was absent. Importantly, a template purine nucleotide should follow on the 3′ side of this dC or dT to allow for the stabilizing stacking interaction between the initiating rNTP and this template purine (Basu et al. 2014). The other nucleotides, dG and dA, were rarely chosen, Thus, to direct site-specific initiation at a coligo stem–loop junction, a single dC or dT should be embedded among dA and dG nucleotides in the 5 nt spanning the junction. When there is only one pyrimidine (C or T) in this 5 nt window, then that pyrimidine has a secondary structure-based preference to be a strong transcription initiation site. If there is more than one pyrimidine within the 5 nt window, each can template initiation, but initiation is strongly preferred at pyrimidines (C > T) followed by a purine. If more than one pyrimidine in the window is followed by a purine, then a pyrimidine sandwiched between two purines is the stronger initiation site. These trends are exemplified by the following start sites (underlined, and see Fig. 3): 15a-3 5′GCA > TTG; mut2B GCG > ATA; 122 ATA > TTG > TTT; 221 CTG > TTG > GTT; 143-2 ACA > TAA; Dcr3 GCA > CAA. The stem should also have a small symmetrical internal loop within 8 bp of the junction (Lama et al. 2014), as in 15a-3, mut2B, 122 and 143-2; the larger loop should be ≥10–11 nt; and the smaller loop should not exceed ∼5–6 nt (Patel et al. 2016).
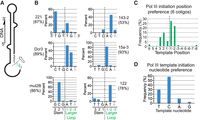
Pol III transcription initiation preferences on hairpin-encoding DNA coligos. (A) Schematic depiction of a coligo stem-larger loop junction with numbering to distinguish stem and loop nucleotide positions. The dotted line marks the junction location here and in panels B and C. The 5 nt junction window described in the text is from nt 3 to −2. (B) Pol III transcription initiation position preference for each of the six coligos studied. The percentage of reads initiating within the 5 nt junction window is summed and shown in parentheses under the coligo name. Coligo 122 percentage includes initiations with one or two untemplated adenylates (see text). The average percentage of reads initiating within the 5 nt window for the six coligos was 81%. (C) Pol III transcription initiation positions in relation to the stem-larger loop junction for the six coligos studied. Initiation events were concentrated in the 5 nt window from 3 to −2. (D) Transcription initiation nucleotide preference for the six coligos studied. Initiation events took place mainly on template dC and dT positions.
Termination determinants: junction secondary structure
The stem-larger loop junction region, including the larger loop, also holds the main determinants for region-specific termination. Almost all termination occurred in the larger loop within 11 nt following the ds stem (Fig. 4A,B). The five coligos sequenced here have larger loops of 11 or 12 nt, and so it initially appeared that completing transcription of the larger loop, and reapproaching the initiatior, led to termination, since no reads with 3′ ends at the initiation region or in the stem beyond it were found in the RNA excised for sequencing. However, coligo Dcr3 (Lama et al. 2019) has a 24 nt loop, and in this case termination also began and concluded within the first 11 nt following the stem-larger loop junction. Based on this evidence, the coligo feature correlating with termination appears to be the stem–loop junction followed by a terminal loop of at least 10 to 11 ss nucleotides. The 5- to 6-nt size of the smaller loops did not support termination. We speculate that once the polymerase finishes transcribing the coligo stem, the stem base-pairing register and stem–loop junction reform, slowing further entry of the coligo as a single strand into the Pol III active site. Here, the polymerase may slow or pause, allowing the weakly bound ternary complex to dissociate. We have not been able to detect a complex between Pol III and a radiolabeled coligo (not shown), or the nascent transcript (Lama et al. 2014). This scenario is distinct from oligo dA-dependent yeast Pol III termination at ds genes, which involves the nontemplate strand (Arimbasseri and Maraia 2015), a feature missing in the ss coligo. In vitro, Pol III preparation details can influence how Pol III stops transcription (Nielsen et al. 2013; Nielsen and Zenkin 2014), and one alternative mode of termination in yeast Pol III appears to depend on hairpin RNA formation upstream of the termination site (Nielsen et al. 2013; Arimbasseri et al. 2014).

Pol III transcription termination preferences on hairpin-encoding DNA coligos. (A) Coligo schematic with numbering at the 3′ end of the stem-larger loop junction to distinguish stem and loop positions. (B) Chart showing the frequency of Pol III transcription termination in relation to the stem-larger loop junction for all six coligos studied. (C) As in panel B, but only for the three coligos having a larger loop site resembling the canonical oligo dA Pol III termination sequence. (D) As in panel B, but for only the three coligos lacking a Pol III termination sequence. (E) Diagram illustrating the defining features of (right, green) the coligo initiator structure: a 5-nt stretch spanning the stem-larger loop junction, with preference for initiation at a pyrimidine (dC or dT) followed by a purine; and (left, red) the termination region: the 10 to 11 ss nt in the larger loop following the stem. (Tss) transcription start site.
No two coligos had identical termination patterns, but when we compared the termination position for coligos with and without a Pol III oligo dA terminator sequence in the larger loop (Fig. 4C,D), we found that termination in both groups was distributed among the first 10 to 11 ss loop nucleotides. Coligo 221's asymmetric bulge was the only exception, but structurally this may resemble the start of a larger loop, causing some early termination. We also point out that coligo Dcr3 has three separate oligo dA terminator sites, two in the larger loop and one in the stem (Lama et al. 2019), yet termination in the sequenced transcripts only occurred at the site situated in the first 11 nt of the larger loop (Lama et al. 2019). Based on our results, and with regard to the predictive determinants of coligo transcription, termination appears to depend on the DNA coligo stem–loop junction secondary structure, followed by a ss stretch of about 11 nt. While we can't rule out some contribution from the oligo dA sites in 15a-3, 143-2, and Dcr3, we consider termination near their oligo dA more likely coincidental.
The probability of terminating after one circumtranscription was high, as described by the termination efficiency of the coligos, defined as the percentage of n = 1 terminated transcripts per summed n = 1, 2, 3, 4 transcripts (Table 1). This percentage was on average 82% (range: 65% to 91%). The small percentage of transcripts that escaped termination after the initial circumtranscription had a similarly high probability of terminating after the second turn around the coligo, and so forth, until after n = 4 their abundance approached zero. We point out that five out of the six coligo larger loops were A/T-rich (Table 1), a contributing factor in gene-context Pol III termination regions (Mishra and Maraia 2019). Within the range of A/T-richness in the coligos studied here (50%–75%, Table 1), there was no obvious correlation between termination efficiency and larger loop A/T percentage.
A coligo transcription model
Based on these and prior results, our overall model for a coligo transcription event is as follows: first, the coligo associates transiently with Pol III (a steady state level has not been detected by gel shift or pull-down experiments) by virtue of its physical resemblance to the natural promoter-triggered open complex, with the larger loop resembling the transcription bubble and the coligo stem presumably located in the enzyme's ds DNA binding cleft (Hoffmann et al. 2015; Patel et al. 2016). The 5 nt initiator window is positioned in the active site, and in some fraction of the complexes a burst of transcription one time around the coligo occurs as the coligo stem is melted and the coligo is reeled through the active site. Following circumtranscription, when the larger loop enters the active site, the stem base-pairing register can reform, possibly reestablishing the coligo secondary structure. In contrast to most RNA made from tailed Pol III templates (Campbell and Setzer 1992), we found that most of the transcripts do not remain bound to the coligo template following transcription (Seidl et al. 2013; Lama et al. 2014), probably because both transcript and coligo have strong intramolecular secondary structure. We therefore speculate that reforming the secondary structure slows transcription, and the weakness of the complex causes it to dissociate. A small percentage of complexes persist long enough to begin a second circumtranscription before once again encountering a high probability of termination, and so forth, leading to a small amount of rapidly diminishing tandem multimer transcripts. We cannot rule out a dissociative mechanism, wherein some RNA:DNA hybrids form and rebind the enzyme, leading to the tandem multimers, but this scenario seems less likely given the steady-state weakness of binding between DNA, polymerase and RNA.
Conclusion
Coligos are small RNA expression vectors that can encode the fundamental and ubiquitous RNA secondary structure motif of the imperfectly base-paired stem–loop hairpin. Coligo-seq has here enabled us to identify unequivocally the coligo stem-larger loop junction as a site-specific, secondary structure-based Pol III transcription initiator with very good initiation site predictability. A single pyrimidine, sandwiched between purines within a 5 nt window at the junction, yields a predictable transcription start site, and most transcripts will terminate with variable 3′ end heterogeneity within the first 11 nt of the larger ss loop after circumtranscription. Prior results suggest that these findings will apply to coligos transcribed by Pol III in human cells. Thus, coligos potentially provide a chemically synthesized, promoter-independent alternative to plasmid and virus-based small RNA expression in human cells.
MATERIALS AND METHODS
Coligo preparation
Linear coligo precursor oligonucleotides were purchased from Integrated DNA Technologies (IDT, Ultramer oligos) with 5′ phosphates. Their sequences are as follows: 5′pTTGCCTAGCAGTAGCTATTTAGTGTGATAATGGCGTTTGATAGTTTAGACACAAACACCATTGTCACACTCCACAGCTCTGCTAAGGAA (122), 5′pGAAGGAAAAATCAGTTTTGCATAGATTTGCACAACTACATTCTTCTTGTAGTGCAACTATGCAAAACTGCAAACAAAAACTA (Dcr3), 5′pGACACGCCTGAAGTCTCTGATTAATAAATTGTATTAATCAGAAACTTCAAGCGATAGTTTA (mut2B), 5′pACGAACACAGAAATCTACATTGTATGCCAGGTTCATGAAACCCAGCAGACAATGTAGCTGTTGCCTA (221), 5′pAGACCGAGATGCAGCACTGCACCTCTTCCTGAGCTACAGTGCTTCATCTCACAAAA (143-2), and 5′pATGACACAAACCATTATGTGCTGCTACTTTGCAGCACAATATGGCTTGCAAAA (15a-3). Coligo and coligo precursor secondary structure was predicted using the Mfold Web Server or the RNA fold Vienna package (Zuker 2003). Coligo precursors, discontinuous in regions predicted to be single-stranded (ss), were circularized using the TS2126 RNA ligase I (Rnl1), gel purified and isolated as previously described (Seidl and Ryan 2011; Seidl et al. 2013).
Adapter and primer oligonucleotides
Custom sequencing adapters and primer oligonucleotides used for the preparation of coligo transcript libraries were ordered from IDT and purified using denaturing polyacrylamide gel electrophoresis (DPAGE). Adapters and primer sequences for library preparation and sequencing on Illumina MiSeq were based on the NEBNext Small RNA Library Preparation Kit (NEB, E7330S) as previously described (Lama et al. 2019) but with the following reverse PCR primers containing different index sequences, N6: 5′CAAGCAGAAGACGGCATACGAGATN6GTGACTGGAGTTCAGACGTGTGCTCTTCCGATCT, where N6 are as follows: 122: ATTGGC; Dcr3: GATCTG; mut2B: TACAAG; 221: CGTGAT; 15a-3: ACATCG; 143-2: TGGTCA.
In vitro transcription
FLAG-Pol III was immunoprecipitated from whole cell extract (WCE) made from a HEK293/POLR3F stable cell line (Chiu et al. 2009), as previously described (Lama et al. 2014), using the FLAGIPT1 kit (Sigma–Aldrich). Briefly, 40 µL of drained anti-Flag M2-Agarose Affinity Gel was used with the whole cell extract from three 10 cm plates of cells, which typically yielded 1.4 mL of whole cell extract at 3–4 µg/µL total protein. The washed beads were used as the source of Pol III in transcription reactions. In vitro transcription reactions were carried out as previously described (Lama et al. 2019). The (100 µL) transcription reactions contained Pol III beads (20% of one entire three plate Pol III immunoprecipitation preparation), 80 U RNasin RNase inhibitor (Promega), 0.6 mM each ATP, GTP, CTP, 0.2 mM UTP, ∼2 µCi [α-32P]-UTP, 40 mM Tris-HCl pH 7.9 (at 25°C), 6 mM MgCl2, 10 mM DTT, 2 mM spermidine, and 100 nM DNA coligo. The typical concentration range for UTP in eukaryotic cells is 0.23 to 0.69 mM (Traut 1994). In order to increase the specific activity of the incorporated 32P label during transcription, we lowered the total UTP concentration in the transcription reactions to 0.2 mM. We verified that the transcript patterns did not change significantly between the two concentrations (not shown).
Coligo-seq and data analysis
Coligo-seq was carried out exactly as described in detail elsewhere (Lama et al. 2019). Since each library contains a unique barcode sequence, the libraries were pooled and sequenced together on an Illumina MiSeq instrument (110 cycles) at the Personalized Genomic Medicine Laboratory, Department of Pathology and Cell Biology, Columbia University Medical Center. Bioinformatics analyses for all the coligo libraries were also done following the protocol as described for coligo Dcr3 (Lama et al. 2019). The raw Coligo-seq data have been deposited to the Gene Expression Omnibus (SRA accession number: SAMN07414566).
ACKNOWLEDGMENTS
This work was supported by the National Institutes of Health (5SC1GM083754 to K.R.). Additional infrastructure support at the City College of New York was provided by grant 5G12RR003060 from the National Center for Research Resources and grant 8G12MD7603 from the National Institute on Minority Health and Health Disparities. We thank Dr. Peter Nagy of the Personalized Genomic Medicine Laboratory, Department of Pathology and Cell Biology, Columbia University Medical Center, for Illumina sequencing.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.078981.121.
- Received September 13, 2021.
- Accepted July 6, 2022.
This article is distributed exclusively by the RNA Society for the first 12 months after the full-issue publication date (see http://rnajournal.cshlp.org/site/misc/terms.xhtml). After 12 months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.
REFERENCES
MEET THE FIRST AUTHOR

Meet the First Author(s) is a new editorial feature within RNA, in which the first author(s) of research-based papers in each issue have the opportunity to introduce themselves and their work to readers of RNA and the RNA research community. Lodoe Lama is the first author of this paper, “Starting and stopping RNA polymerase III transcription on single-stranded DNA oligonucleotides.” This work was conducted in Dr. Kevin Ryan's laboratory at the City College of New York while Lodoe was a graduate student in the City University of New York Biochemistry Ph.D. program. Lodoe is now a postdoctoral associate in Dr. Thomas Tuschl's laboratory at Rockefeller University.
What are the major results described in your paper and how do they impact this branch of the field?
Circularized DNA oligonucleotides, or coligos, provide an alternative approach to expressing small RNA in human cells. These chemically synthesized templates are transcribed by human RNA polymerase III, but for these easily made templates to become generally useful, that is, to encode small RNA with defined 5′ and 3′ ends, I wanted to investigate what Pol III looks for in the template to initiate and terminate transcription. We used immunoprecipitated Pol III to transcribe a group of coligo templates and then exhaustively cataloged the end heterogeneity by a modified small RNA-seq method that we call coligo-seq. We found that Pol III begins transcription exclusively at the junction where a DNA helix transitions to a terminal single-stranded loop of about 11 or more nucleotides, and transcription always proceeds into the helix, never into the loop, so like a promoter it is directional. The sequence at the initiation site is also important: a single pyrimidine sandwiched between purines within a 5-nt window at the junction yields a predictable transcription start site. Termination is also predictable but occurs over a stretch of single-stranded loop nucleotides, leading to greater 3′ than 5′ end heterogeneity. When Pol III reads through this site, it has a second high probability chance at termination the next time around the coligo, preventing the “rolling” transcription observed with bacteriophage RNA polymerases.
What led you to study RNA or this aspect of RNA science?
Small RNAs (sRNA) like microRNA (miRNA), small interfering RNA (siRNA), CRISPR and other types of RNAs, have an interesting function and a promising therapeutic potential. However, successful translation of their therapeutic potential has been hindered by the fragility and expense of unmodified RNA, and in some cases the nature of the vectors used to express them, like viruses and plasmids. We all now know that RNA itself can be a valuable medicine, but I envision that synthetic DNA can provide a more stable and less expensive way to introduce RNA sequence into cells. I wanted to develop sRNA expression templates that leave the difficult job of making the RNA to the cells and leverage the relative stability and economy of synthetic DNA. How far can we go with these templates? That's what I want to know.
During the course of these experiments, were there any surprising results or particular difficulties that altered your thinking and subsequent focus?
One of the major difficulties encountered while performing these experiments was the inability of the commonly used two adapter ligation method to capture hairpin RNA for cDNA library synthesis (mainly due to poor 5′ adapter ligation when the 5′ end is close to the end of a helix). This forced me to pause my research while I stopped to adapt small RNA-seq to RNA hairpin substrates. Later, I learned this was a general problem but was rarely talked about. Along the way, I learned that if you don't check each step in a multistep protocol, it's hard to be confident of the end result! I also continue to be surprised at our progress in directing site-specific transcription initiation on coligo templates. We're not as fast as promoter-based transcription, but we are approaching homogeneous initiation.
If you were able to give one piece of advice to your younger self, what would that be?
Be courageous and ask scientifically naive questions. And as my graduate advisor told me, “Don't ask me if you can try it; if it's not dangerous, just do it and tell me about it later.”








