
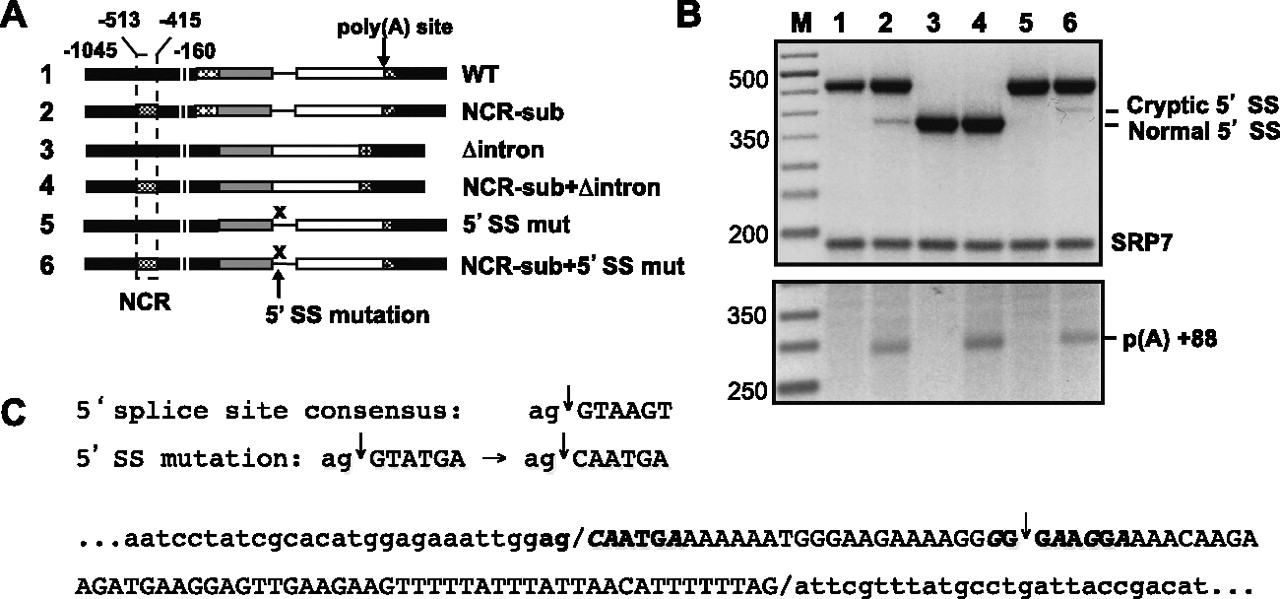
(A) Diagrams of chimeras with 5′ DNA derived solely from rem1 (odd numbers) or containing a replacement of the NCR with nmt81 sequences (even numbers). In constructs 1 and 2, the remainder of the gene is wild type. In constructs 3 and 4, the intron has been precisely deleted. In constructs 5 and 6, the nearly invariant GT at the 5′ end of the intron was mutated to CA, which generally abolishes use of a 5′ splice site (Alvarez and Wise 2001). (B) Splicing (top) and polyadenylation (bottom) assays on the chimeras diagrammed in panel A. (C) Identification of the cryptic 5′ splice site activated when the NCR is replaced in conjunction with mutating the natural exon/intron boundary. (Top) Transversions at the first two nucleotides of the intron were used to inactivate the natural 5′ splice site (Alvarez and Wise 2001). (Bottom) DNA sequence of the intron (uppercase) and surrounding exons (lowercase) with boundaries indicated by slashes. The cryptic 5′ junction, previously noted by Ayté and colleagues in an allele with nmt41 sequences replacing the entire upstream region (Moldon et al. 2008), is marked with an arrow. The octanucleotides encompassing both the natural and cryptic 5′ splice sites are highlighted in bold, with nucleotides that deviate from the fission yeast consensus (Wood et al. 2002) indicated by italics.








