
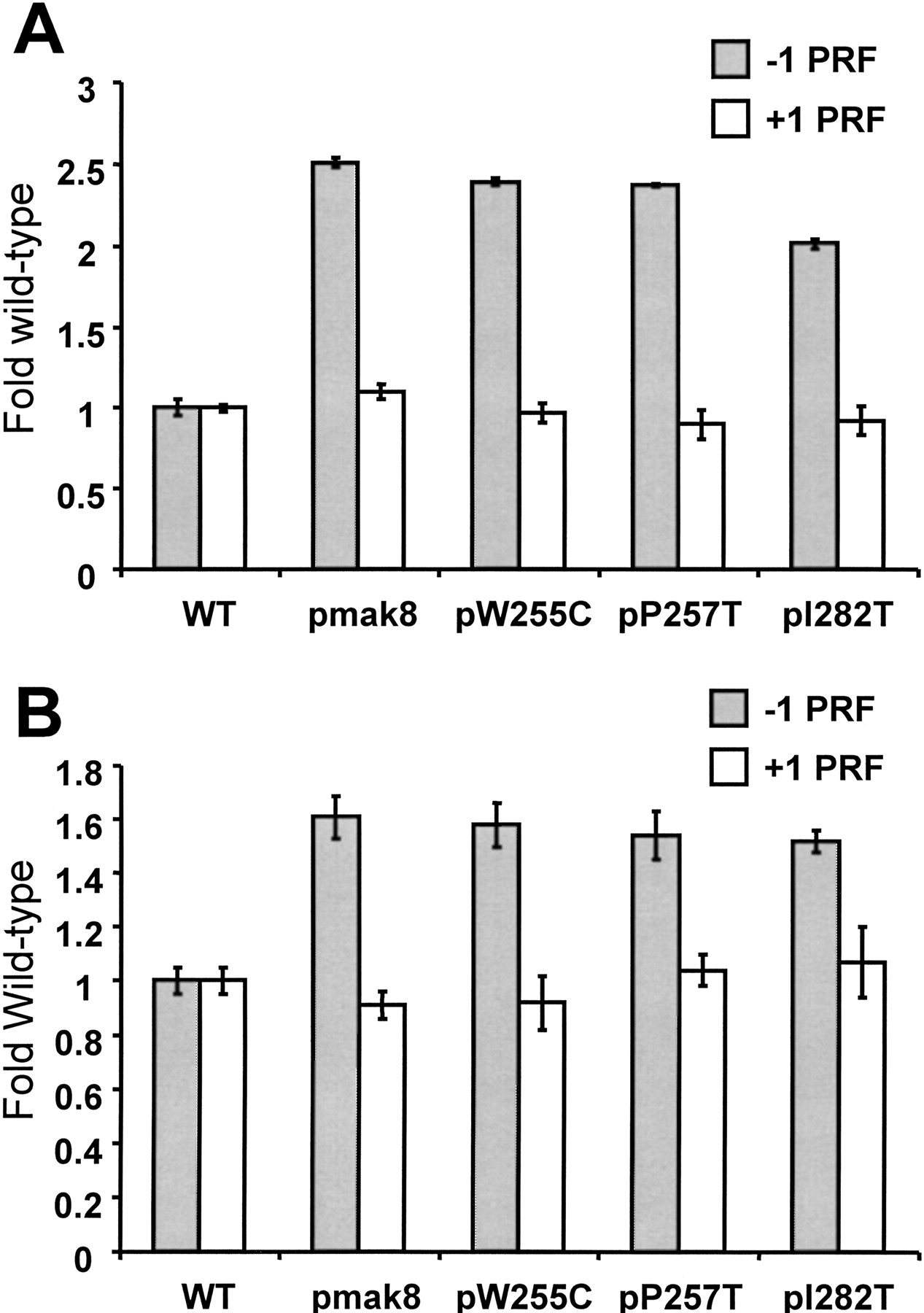
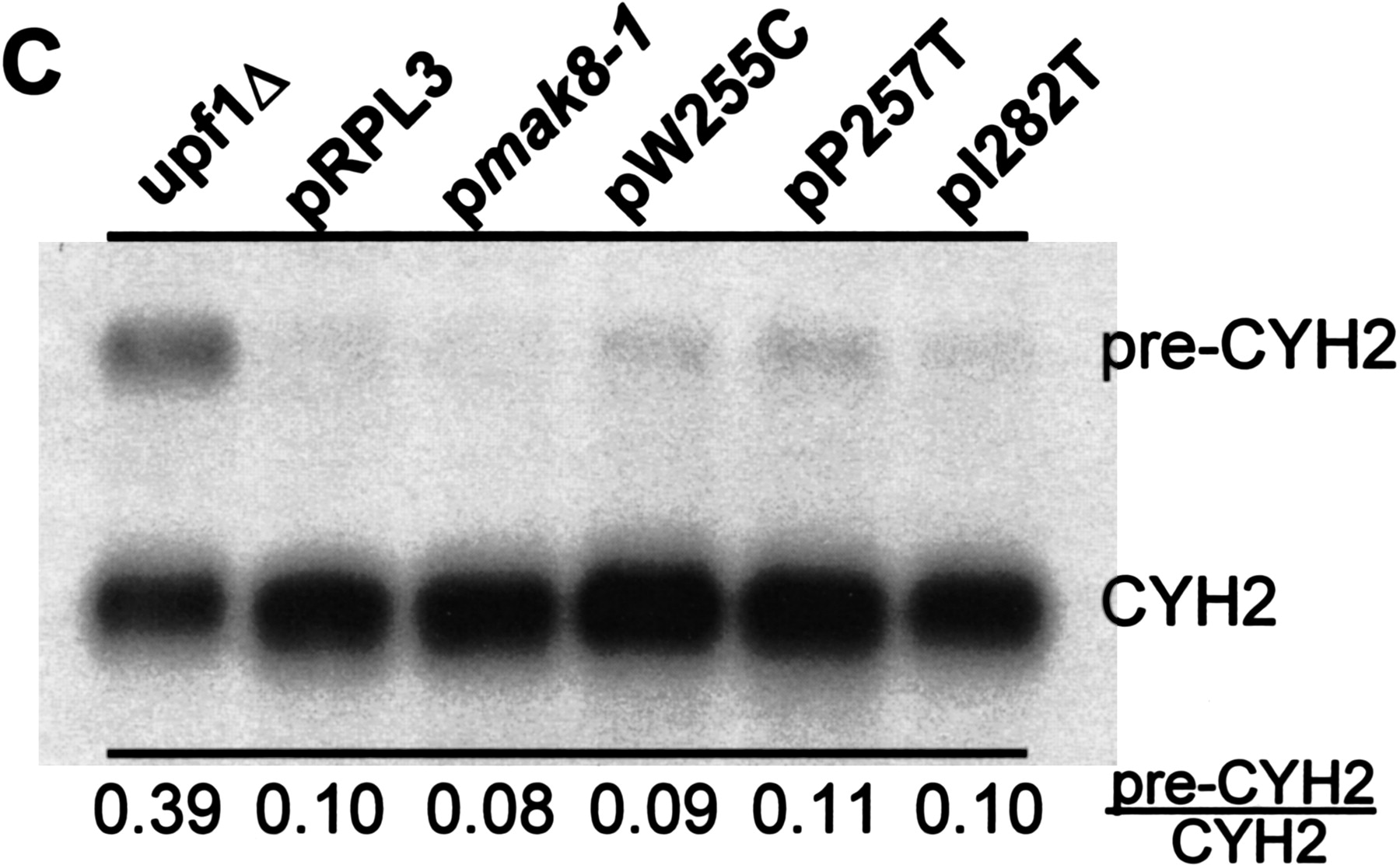
Elevated efficiencies of −1 programmed ribosomal frameshifting in cells expressing mutant rpl3. (A) Monscistronic PRF reporter plasmid system. Isogenic strains harboring the TRP1–CEN6-based RPL3 alleles were cotransformed with p0 or p−1 (LEU2–CEN6-based vectors), or p0 or p+1 (URA3–CEN6) and programmed ribosomal frameshifting efficiencies were determined as described (Dinman et al. 1991; Peltz et al. 1999). Changes in frameshift efficiencies are depicted in terms of fold wild-type (see Table 1). Each dataset represents the averages of three individual experiments repeated in triplicate. Error bars denote standard deviations from the means. (B) Bicistronic PRF reporter plasmid system. Isogenic strains harboring the TRP1–CEN6-based RPL3 alleles were cotransformed with bicistronic URA3–CEN6 based Renilla–firefly luciferase 0-frame, −1 (L-A derived), or +1 (Ty1-derived) reporter vectors. Renilla and firefly luciferase activities of clarified cell lysates were determined using Dual-Luciferase Assay Reagents (Promega) and quantitated using a TD20/20 lumineter (Turner designs). Frameshifting efficiencies were calculated by dividing the firefly/Renilla luminescence ratios from the −1 and +1 programmed frameshift test reporters by the 0-frame control reporter. Each dataset represents the averages of three individual experiments repeated in triplicate. Changes in frameshift efficiencies are depicted in terms of fold wild-type (see Table 1). Error bars denote standard deviations from the means. (C) Expression of mutant forms of L3 do not affect the nonsense mediated mRNA decay (NMD) pathway. Total cellular mRNA was extracted from mid-logarithmically growing rpl3Δ cells harboring plasmids encoding the wild-type or mutant alleles of RPL3 as indicated. mRNA from PLY36 cells harboring the upf1Δ|mBhisG allele (upf1Δ) was used as a control. Total RNA from each sample was separated through a 1.0% agarose formaldehyde denaturing gel and transferred to a nylon membrane. The RNA blot was subsequently hybridized with a radiolabeled CYH2 probe as previously described (Cui et al. 1996). The image was visualized, and band intensities were quantitiated using a Molecular Dyanmics PhosphorImager. The locations of the NMD-sensitive CYH2-precursor mRNA (pre-CYH2) and of the NMD-insensitive mature CYH2 mRNA are indicated, as are the pre-CYH2/CYH2 signal intensity ratios.








