
An siRNA ribonucleoprotein is found associated with polyribosomes in Trypanosoma brucei
Abstract
Genetic interference by double-stranded RNA (dsRNA) or RNA interference (RNAi) triggers the sequence-specific degradation of cellular transcripts. The mediators of mRNA degradation are small interfering RNAs (siRNAs). Here, we report that in Trypanosoma brucei, 10%–20% of siRNAs cosediment with polyribosomes. Preventing the assembly of ribosomes on mRNAs results in a concomitant decrease of siRNAs sedimenting as high-molecular-weight complexes. We further provide evidence that siRNAs are associated with translating ribosomes and that this association is mediated by a 70-kD ribonucleoprotein complex.
Keywords
INTRODUCTION
RNA interference (RNAi) triggered by double-stranded RNA (dsRNA) is an evolutionarily conserved process and exists in a wide variety of organisms, throughout the range of protozoa, plants, fungi, and animals (Sharp 2001; Zamore 2001; Hannon 2002; Hutvagner and Zamore 2002b). In the current model for RNAi, the first step is recognition and cleavage of dsRNA by an RNase III-like nuclease termed Dicer (Bernstein et al. 2001). dsRNA is either derived from cellular sources like viruses, transgenes, or transposons, or exogenously introduced into cells. By the action of Dicer, dsRNA is converted into small 21–25-nt-long fragments, known as small interfering RNAs or siRNAs (Hamilton and Baulcombe 1999; Zamore et al. 2000; Elbashir et al. 2001). After cleavage by Dicer, siRNAs join a multicomponent nuclease complex, termed RISC (RNA-induced-silencing complex), that uses siRNAs as guides to select target mRNAs for degradation (Hammond et al. 2000; Martinez et al. 2002; Schwarz et al. 2002).
The development of cell-free systems recapitulating RNAi has been instrumental in gaining an insight into the mechanistic details of siRNA-mediated mRNA degradation (Hannon 2002). Similarly, these extracts provided an entry point into the characterization of components of the RNAi machinery (Hannon 2002). Initial experiments using Drosophila S2 cell extracts revealed that the RISC complex pelleted at 200,000g, which represents a fraction enriched in polyribosomes, and that this complex appeared to have a molecular mass ranging in size from 230 to 500 kD (Hammond et al. 2000, 2001; Nykanen et al. 2001). On the other hand, more recent experiments with affinity-purified human and Drosophila RISC reported a size of ~90–160 kD (Martinez et al. 2002) and the identification of eIF2C1 and eIF2C2 as major components of the complex. Biochemical purification of the Drosophila RISC first identified Argonaute-2 (Ago-2) as a component of this complex (Hammond et al. 2001), and, more recently, a Drosophila fragile X-related protein (dFMR1) and VIG were also found to be associated with RISC (Caudy et al. 2002). Furthermore, Hutvagner and Zamore (2002a) presented data that the human RISC contained not only eIF2C, but also the putative helicase Gem3 and a novel protein Gem 4. In an independent study, the isolation of a Drosophila FMR1-associated complex revealed Ago-2, a Drosophila homolog of p68 helicase, and two ribosomal proteins, L5 and L11, as well as 5S rRNA (Ishizuka et al. 2002). Considering that dFMR1 appears to be important for the translation of some specific mRNAs, although the mechanism remains unknown, these findings point to a possible intersection of components of the RNAi and translational machineries. Furthermore, Carthew and colleagues have recently shown that mutations in aubergine and spindle-E block RNAi activation and perturb translational control during Drosophila oocyte development, indicating a connection between RNAi and translation in the oocyte (Kennerdell et al. 2002).
We previously reported that similar to Drosophila, a proportion of siRNAs sedimented as large complexes at 100,000g in the protozoan parasite Trypanosoma brucei (Djikeng et al. 2001). Here, we show that in steady-state conditions, ~10%–20% of siRNAs are found associated with polyribosomes, that this association is sensitive to inhibition of translation initiation, and that the ribosome-associated siRNAs can be released by salt extraction as a ribonucleoprotein complex with an apparent molecular mass of 70 kD.
RESULTS
A portion of siRNAs sediment as high-molecular-mass complexes in sucrose density gradients
We have previously shown that in trypanosomes ~10% of siRNAs, derived either from actin dsRNA expressed from a transgene or from endogenous retroposon transcripts, sedimented at 100,000g, where polyribosomes are found (Djikeng et al. 2001). An analysis of two additional cell lines expressing dsRNA corresponding to an RNA methyltransferase (MT40) or the green fluorescent protein (gfp) gave similar results, with up to 20% of the siRNAs fractionating in the 100,000g pellet (data not shown). To further analyze these siRNA-containing complexes, cytoplasmic extracts from trypanosomes expressing actin, MT40, or gfp dsRNAs were fractionated by centrifugation through 15%–50% sucrose density gradients. Cycloheximide (100 μg/mL) was added to the cells just before lysis to “freeze” polyribosomes. As can be appreciated from the A254 profiles (Fig. 1A), the centrifugation conditions used separated polysomes from 80S monosomes, whereas soluble material and small particles were found near the top of the gradient. Total RNA isolated from each fraction was passed through a microcon 100 filtration unit, and the filtrate, enriched for low-molecular-weight RNAs, was analyzed by Northern blot hybridization with gene-specific sense riboprobes. In all three cases, the antisense siRNAs fractionated into two pools: the majority (~80%) of antisense actin (Fig. 1B), MT40 (Fig. 1C), or gfp (Fig. 1D) siRNAs sedimented near the top of the gradient and therefore was not present in high-molecular-weight complexes. In agreement with our previous results (Djikeng et al. 2001; data not shown), the remainder of antisense siRNAs (~20%) sedimented as high-molecular-mass complexes throughout the heavier fractions of the gradient. On one hand, the sedimentation profile could be interpreted to indicate that siRNAs exist as a heterogeneous collection of complexes spanning a wide range of molecular weights. On the other hand, one can envision that siRNAs might be associated with ribosomes and that the observed distribution correlated with the fractionation of polyribosomes.
To begin to address the latter scenario experimentally, we investigated whether a blockage of ribosome assembly on mRNAs had any effect on the sedimentation behavior of siRNAs. To this end, we took advantage of pactamycin, a protein-synthesis inhibitor that prevents translation initiation both in prokaryotic and eukaryotic organisms (Brodersen et al. 2000). Trypanosome cells expressing MT40 dsRNA were incubated for 30 min in the presence of 200 ng/mL pactamycin, a treatment that resulted in nearly complete inhibition of protein synthesis (data not shown). Cytoplasmic extracts from pactamycin-treated and control cells, incubated with cycloheximide, were then analyzed by sucrose density gradient centrifugation (Fig. 2). Compared with control cells, the amount of polyribosomes in pactamycin-treated cells was substantially reduced with the concomitant increase of 80S monosomes (cf. Fig. 2A and 2E). These 80S ribosomes represent nontranslating ribosomes, because they are dissociated into their component subunits by treatment with high salt (data not shown). To obtain a quantitative measurement of the distribution of polyribosomes in the two gradients, RNA was extracted from each fraction, and an aliquot was hybridized to a probe complementary to 5S rRNA, which is associated with the 60S subunit (Fig. 2B,F). After phosphorimager quantitation, the percent 5S rRNA in each gradient fraction was plotted as a function of the total 5S rRNA present in each gradient (Fig. 2I). By this analysis, it appeared that, relative to control cells, pactamycin treatment resulted in an approximately twofold decrease in the amount of ribosomes sedimenting in the polyribosome region of the gradient (fractions 5–12).
Next, we analyzed the distribution of antisense MT40 siRNAs throughout the gradients by Northern blot hybridization with a radiolabeled sense riboprobe (Fig. 2C,G). Similar to what is shown in Figure 1, in control cells ~20% of the antisense MT40 siRNAs were present in the heavier fractions of the gradient (Fig. 2C). In contrast, in pactamycin-treated cells, there was a substantial decrease in the amount of antisense siRNAs sedimenting in the heavier fractions of the gradient (Fig. 2G). Quantitation of the antisense siRNAs showed that pactamycin treatment resulted in a two- to threefold reduction in the proportion of siRNAs sedimenting in fractions 5–12, as compared with the control extract. Similar results were obtained when the Northern blots of Figure 2, C and G, were stripped and rehybridized with an antisense riboprobe that detects the sense strand of the siRNAs (Fig. 2D,H). Thus, we observed that inhibition of translation initiation, which prevents the assembly of polyribosomes, changed the sedimentation of siRNAs from the heavier to the lighter fractions of the gradient. Furthermore, the extent of blockage of polyribosome assembly correlated roughly with the decrease of siRNAs sedimenting as large complexes. Similar results were obtained with cells expressing actin siRNAs (data not shown). Taken together, the data presented so far indicate that siRNAs may be associated with polyribosomes.
A proportion of MT40 siRNAs is found associated with translating ribosomes
If MT40 siRNAs are, indeed, associated with polyribosomes, then they should cosediment with translating 80S ribosomes. To test this prediction, we generated 80S ribosomes from polyribosomes by treating a cytoplasmic extract from trypanosomes expressing MT40 dsRNA with micrococcal nuclease. It is known that under limiting digestion conditions, micrococcal nuclease cleaves the mRNA in between translating ribosomes and generates 80S particles, representing translating ribosomes. Preliminary experiments established that incubation of trypanosome extracts with 100 U/mL of micrococcal nuclease for 30 min at 4°C was sufficient to convert most polyribosomes into monosomes, and that under these conditions ~60% of the MT40 siRNAswere resistant to digestion (data not shown). Therefore, a mock-treated extract and a micrococcal nuclease-treated extract were fractionated on a 5%–45% sucrose gradient using centrifugation conditions to better separate the 80S ribosomes from polyribosomes (Fig. 3). Given that under these conditions large polyribosomes will pellet, the gradients were overlaid onto a 60% sucrose cushion to collect them (lane C). In the mock-treated extract, MT40 siRNAs were found at the top of the gradient (fractions 1–3), in fractions 9–12, corresponding to small polyribosomes with about three to six ribosomes, and in the fraction corresponding to the sucrose cushion (lane C), which is enriched in heavy polyribosomes. In the micrococcal nuclease-treated extract, siRNAs sedimented near the top of the gradient and in fractions 6–9, encompassing the 80S peak, plus a residual amount of polyribosomes (Fig. 3B). A very small proportion of siRNAs were detected in the sucrose cushion (lane C). Thus, the micrococcal nuclease treatment not only converted polyribosomes to 80S particles, it also resulted in a clear shift of the heavy-sedimenting siRNAs to fractions corresponding to 80S particles. This strongly implied that a proportion of MT40 siRNAs is associated with translating ribosomes.
A 70-kD complex containing siRNAs
Previous studies from Hannon and colleagues established that RISC and most siRNAs can be extracted from a 200,000g pellet from Drosophila S2 cell extracts with 400 mM potassium acetate (Hammond et al. 2000). We therefore wished to determine whether in trypanosomes the association of MT40 siRNAs with ribosomes could be disrupted by salt extraction. A postnuclear supernatant from cells expressing MT40 dsRNA was separated at 100,000g into a soluble and a pellet fraction, and the pellet fraction was extracted with 400 mM potassium acetate. siRNAs, initially present in the first high-speed pellet fraction (Fig. 4B, lane 3), were rendered soluble after salt extraction (lane 4), and only a small proportion of siRNAs remained in the second pellet fraction (lane 5). Next, we fractionated the salt-soluble material by gel filtration chromatography on Superdex-200, which revealed that the ribosome-derived MT40 siRNAs had an apparent molecular mass of ~70 kD (Fig. 4C). siRNAs homologous to the retroposon INGI cofractionated with MT40 siRNAs released from polyribosomes (Fig. 4D), indicating that both transgene-derived and endogenous siRNAs had identical fractionation properties with a relatively narrow molecular weight range. In contrast, siRNAs from the S100 fraction had a more heterogeneous fractionation profile (Fig. 4E). Because digestion of the salt-extracted material with proteinase-K and phenol extraction prior to chromatography resulted in a clear shift of siRNAs to a lower molecular weight (Fig. 5), this demonstrated that MT40 siRNAs were extracted from ribosomes as a ribonucleoprotein complex.
DISCUSSION
Here we presented evidence that in trypanosomes a portion of siRNAs, in the form of a 70-kD ribonucleoprotein particle, are associated with polyribosomes. In particular, we have shown that siRNAs generated from three different dsRNAs cosedimented in sucrose density gradients with polyribosomes. The portion of siRNAs sedimenting as high-molecular-mass complexes varied between 10% and 20%. By inhibiting the assembly of polyribosomes onto mRNA we observed a concomitant reduction in the amount of MT40 siRNAs sedimenting as high-molecular-mass complexes. However, in the pactamycin-treated extract there was no detectable accumulation of siRNAs with 80S nontranslating ribosomes, indicating that siRNAs did not unspecifically bind to ribosomes. In contrast, we found that siRNAs cosedimented with translating 80S monosomes generated by treatment of a cytoplasmic extract with micrococcal nuclease. In conclusion, our results are consistent with an interaction between the translation and RNAi machinery and provide a plausible explanation for the high sedimentation coefficient of a proportion of siRNAs, as it was originally reported in Drosophila S2 cell extracts (Hammond et al. 2000), as well as in trypanosome cells (Shi et al. 2000).
Salt extraction of a high-speed pellet (containing polyribosomes) from Drosophila S2 extracts released RISC (Hammond et al. 2000). By applying a similar procedure, we identified a protein–siRNA complex with an apparent molecular mass of ~70 kD, but at present we have no evidence that this trypanosome siRNP preparation has RISC activity. The 70-kD molecular mass estimate of the siRNP salt-extracted from polyribosomes is most likely an underestimate of its true mass, because we have recently determined that one of the components of trypanosome siRNPs is an Argonaute-like protein of ~100 kD (H. Shi, A. Djikeng, C. Tschudi, and E. Ullu, in prep.). It is interesting to note that depending on the procedure for extract preparation, as well as the methodology of biochemical purification, the apparent molecular weight of RISC is quite variable. Indeed, the size of the RISC complex initially identified in a Drosophila S2 cell extract was estimated to be ~500 kD (Hammond et al. 2000, 2001), and in Drosophila embryo lysates, there is evidence for RISC complexes smaller than 230 kD (Nykanen et al. 2001). However, more recently the size of affinity-purified human and Drosophila melanogaster RISC was reported as ~90–160 kD (Martinez et al. 2002).
What is the relationship between the siRNA complexes that are present in the S100 fraction and those that are found associated with polyribosomes? By cloning and sequencing actin siRNAs from the S100 and polyribosome fractions, we have previously shown that the two fractions contain a similar distribution of actin siRNAs, indicating that as far as actin siRNAs are concerned, the two populations have a similar informational content (Djikeng et al. 2001). However, the fractionation profile on Superdex-200 of siRNA complexes from the S100 fraction was considerably more heterogeneous than that of siRNA complexes recovered from polyribosomes, both in terms of the molecular weight of the complexes, as well as of the size distribution of the siRNAs. This may be an indication that there are differences in the composition of the S100-derived versus the polyribosome-derived siRNPs.
The findings that siRNA ribonucleoprotein complexes can be almost quantitatively extracted from polyribosomes with a salt concentration of 400 mM and that both sense and antisense siRNAs are found associated with polyribosomes indicate that in steady-state conditions, most polyribosome-associated siRNAs are probably not engaged in hydrogen bonding with the target mRNA. Our results point to the possibility that siRNPs, most likely as a RISC complex, interact directly with one or more components of the ribosome, similar to what has been described in Drosophila (Ishizuka et al. 2002), and that this association is not mediated by the interaction of siRNAs with their specific target mRNA and is thus most likely independent of the identity of the mRNA that is being translated. Furthermore, some components of the nonsense-mediated decay (NMD) pathway are found associated with polyribosomes in steady-state conditions (Atkin et al. 1997). We speculate that the siRNPs bound to translating ribosomes might operate, similarly to the NMD pathway, as a surveillance system to trigger degradation of mRNAs that are being translated.
What might be the role of siRNA complexes associated with polyribosomes? One possibility is that this interaction facilitates recognition between siRNAs and mRNA targets that are actively translated. Base-pairing between siRNAs and the target mRNA might then trigger degradation of mRNA by the RNAi endonuclease while the mRNA is still associated with ribosomes and/or might result in marking the mRNA for subsequent destruction in the cytoplasm. More insight into the significance of the association between the translation and RNAi machinery will require the characterization of the compartment where target mRNA fragmentation takes place in vivo, as well as the identification of the RNAi nuclease and its cellular distribution.
MATERIALS AND METHODS
Trypanosome cell lines
Procyclic T. brucei were transfected as previously described (Ngo et al. 1998). The pLEWMT40 vector expressing MT40 dsRNA was inserted at the rDNA spacer to generate the MT2 cell line. The TAD19-gfp cell line expresses a gfp transgene inserted at the tubulin locus of 29.13.6 cells expressing T7 RNA pol (Wirtz et al. 1999).
Polyribosome analysis
Cytoplasmic extracts were prepared as described previously (Djikeng et al. 2001) but using a modified polysome buffer containing 120 mM KCl, 2 mM MgCl2, 20 mM Tris (pH 7.5), 1 mM DTT, and 10 μg/mL leupeptin. Cycloheximide or pactamycin were added to cells prior to collecting the cells by centrifugation. The drugs were present in all buffers throughout the entire procedure. Postnuclear supernatants from 5 × 108 cells were layered onto 15%–50% sucrose gradients prepared in polysome buffer and centrifuged at 4°C for 2 h at 36,000 rpm in a Beckman SW41 rotor. Then 1-mL fractions were collected using the ISCO gradient fractionation system. The OD254 profile was recorded using the ISCO UA-6 detector. Each fraction was precipitated with 1 mL of isopropanol, and the material was collected by centrifugation. Pellets were resuspended in 0.3 mL of a solution containing 10 mM Tris-HCl (pH 7.5), 1 mM EDTA, 100 μg/mL proteinase-K, and 1% SDS and incubated at 65°C for 30 min. Solubilized and digested material was centrifuged through a microcon-100 filtration unit according to the manufacturer’s instructions (Millipore Corp.), and the filtrate was precipitated with 1 volume of isopropanol after addition of 20 μg of glycogen and NaCl to 600 mM. Samples were then processed for Northern blot analysis as described (Djikeng et al. 2001).
Micrococcal nuclease treatment
A postnuclear supernatant from 5 × 108 cells was digested with 100 U/mL of micrococcal nuclease in the presence of 2 mM CaCl2 for 30 min, and the reaction was stopped by adding EGTA to 25 mM. A mock-treated extract was used as a control. Extracts were overlaid onto a 5%–45% sucrose gradient prepared in polyribosome buffer containing 10 mM MgCl2. A 1-mL cushion containing 60% sucrose was underlaid beneath the gradients to allow recovery of large polyribosomes. The gradients were centrifuged at 4°C for 3 h at 36,000 rpm in a Beckman SW41 rotor. Fractions were collected and processed as described above.
Other procedures
RNA extraction and Northern blotting were performed as described (Djikeng et al. 2001). For gel filtration analysis of siRNA complexes, a 0.4-mL aliquot of S100 or of salt-extracted material from the ribosome pellet was chromatographed on a Superdex-200 column (Amersham Biosciences) equilibrated with polysome buffer. The column was calibrated with thyroglobulin (669 kD), bovine serum albumin (66 kD), and carbonic anhydrase (29 kD). Fractions of 0.5 mL were collected. Digestion of salt-extracted material with proteinase-K (100 μg/mL) was carried out at room temperature for 30 min. RNAs were recovered by phenol extraction and ethanol precipitation before loading onto the Superdex-200 column.
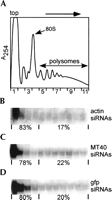
A proportion of siRNAs cosediment with polyribosomes. Sucrose density gradient analysis of cytoplasmic extracts from Trypanosoma brucei cells expressing dsRNA for actin, MT40, and gfp. (A) Representative absorbance profile at 254 nm; the positions of the 80S monosome and polyribosomes are indicated. The positions of polyribosomes, 80S monosomes, and the 40S subunit were verified by examining the distribution of 18S and 28S ribosomal RNAs in the gradient fractions by denaturing agarose gel electrophoresis (data not shown). (B) Northern blot of small RNAs from sucrose density gradient fractions probed with a riboprobe representing the sense strand of actin dsRNA. The quantitation of siRNAs was carried out by phosphorimager analysis. The area corresponding to siRNAs in each lane was separately quantitated, and background was subtracted from each value. An average background value was determined by measuring several equivalent areas of the membrane above and below the siRNA bands. (C) Northern blot with a probe representing the sense strand of MT40 dsRNA. (D) Northern blot with a probe representing the sense strand of gfp dsRNA.
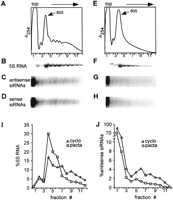
Distribution of MT40 siRNAs in cells treated with (C,D) cycloheximide or (G,H) pactamycin. (A,E) Absorbance profile at 254 nm; the position of the 80S monosome is indicated. (B,F) Northern blot with a probe detecting 5S rRNA. (I,J) Quantitation of 5S RNA and antisense MT40 siRNAs. (cyclo) Control cells treated with cycloheximide; (pacta) pactamycin-treated cells.
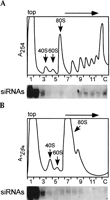
A postnuclear supernatant from cells expressing MT40 dsRNA was either (A) mock-treated or (B) incubated with 100 U/mL of micrococcal nuclease for 30 min on ice. Extracts were fractionated by sedimentation through 5%–45% sucrose density gradients. Below each absorbance profile is the corresponding Northern blot with an MT40 sense riboprobe. The positions of the 40S and 60S subunits and of the 80S monosomes are indicated.
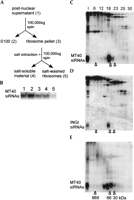
Salt extraction of polyribosomes releases siRNAs as a 70-kD ribonucleoprotein. (A) Outline of the cell fractionation procedure. Numbers in parentheses refer to lane numbers in panel B. (B) Northern blot with the MT40 sense riboprobe of small RNAs from equivalent amounts of postnuclear supernatant (lane 1), S100 fraction (lane 2), ribosomal pellet (lane 3), soluble fraction from the ribosomal pellet extracted with 400 mM potassium acetate after spinning at 100,000g for 1 h (lane 4), and from the salt-washed ribosomal pellet (lane 5). (C) The soluble salt-extracted material from the ribosome pellet shown in lane 4 of panel B was loaded on a Superdex-200 column, and selected fractions were Northern-blotted with a MT40 sense riboprobe. (D) The membrane shown in C was stripped and rehybridized with a sense INGI riboprobe. (E) An aliquot of the S100 fraction was loaded on a Superdex-200 column, and RNA from selected fractions was Northern-blotted with a MT40 sense riboprobe. The elution positions of carbonic anhydrase (30 kD), bovine serum albumin (66 kD), and thyroglobulin (669 kD) are indicated.
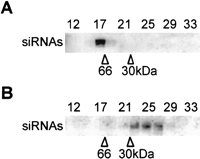
(A) The salt-extracted material from the ribosome pellet was loaded on a Superdex-200 column, and selected fractions were Northern-blotted with a MT40 sense riboprobe. (B) An equivalent amount of the salt-extracted fraction that was analyzed in A was digested with proteinase-K and phenol-extracted prior to chromatography on Superdex-200. The elution positions of carbonic anhydrase (30 kD), bovine serum albumin (66 kD), and thyroglobulin (669 kD) are indicated.
Acknowledgments
We thank Amersham Biosciences for providing pactamycin. Special thanks to Sandra Wolin for her interest and input in this project. We thank Susan Baserga, Shulamit Michaeli, Peter Walter, and Sandy Wolin for critical comments on the manuscript. This work was supported by NIH grant AI28798 to E.U. A.D. was supported in part by a James Hudson Brown-Alexander Brown Coxe Fellowship, and C.T. is the recipient of a Burroughs Wellcome Fund New Investigator Award in Molecular Parasitology.
The publication costs of this article were defrayed in part by payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 USC section 1734 solely to indicate this fact.
Footnotes
-
Article and publication are at http://www.rnajournal.org/cgi/doi/10.1261/rna.5270203.
-
- Accepted March 28, 2003.
- Received February 12, 2003.
- Copyright 2003 by RNA Society








