
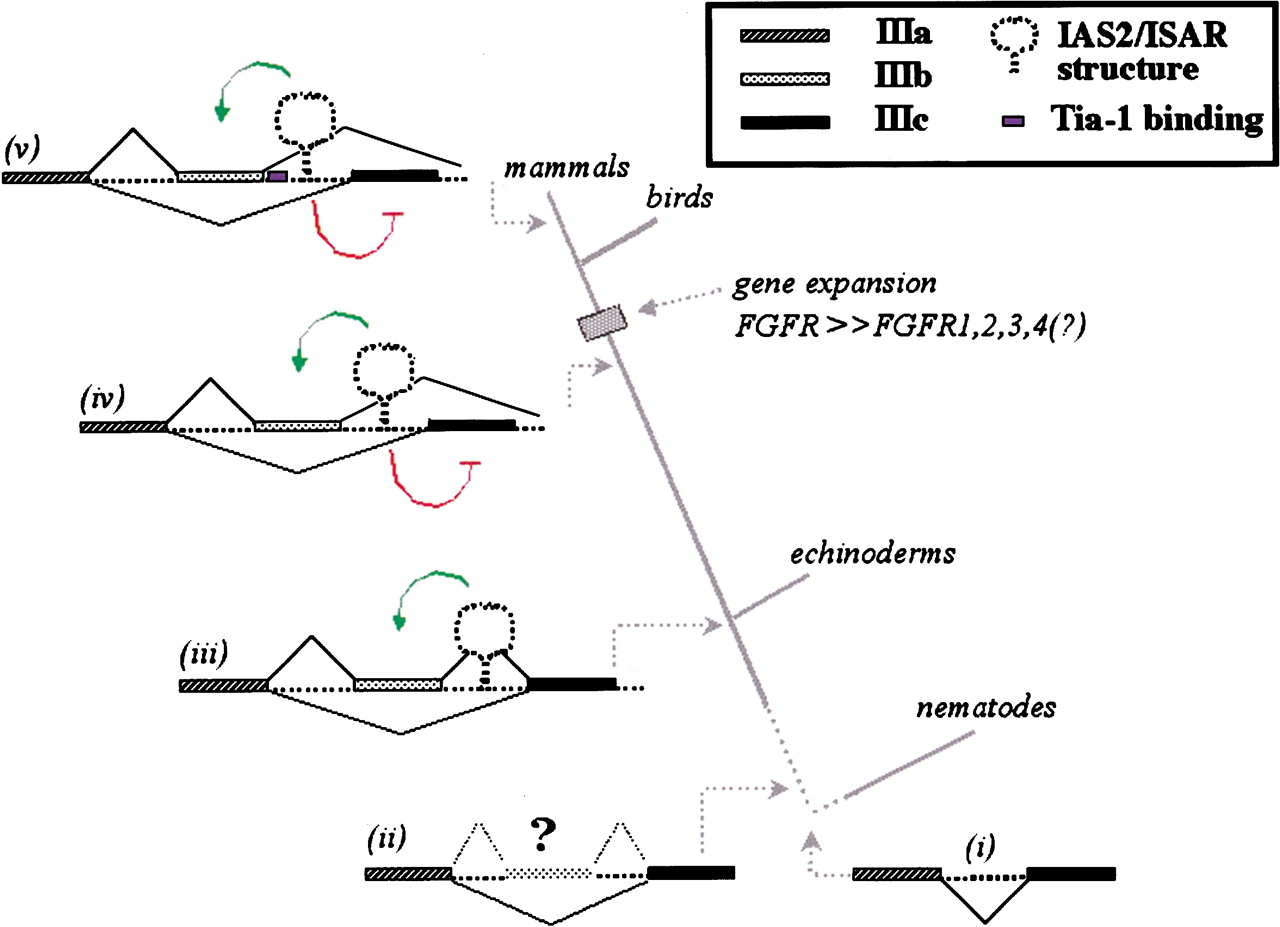
The evolution of the FGF-R IgIII alternative splicing unit. Three FGF-R genes from protostomes had been sequenced previously, the Dfr1 (X74030) and Dfr2 (X74031) in the fruit fly Drosophila melanogaster and the egl-15 gene in the nematode C. elegans (U39761). The D. melanogaster FGF-R Ig domains are all encoded within one exon. The IgIII domain is encoded by two exons in egl-15 and the position of the intron relative to the protein domain has been precisely conserved from vertebrates to C. elegans (i) (De Vore et al. 1995). The amino-terminal half of the domain is encoded by exon 8 (ceIIIa), which is homologous to vertebrate exon IIIa. The carboxy-terminal half of the IgIII domain is encoded by exon 9 (ceIIIc), which is more homologous to IIIc exon of vertebrates than to the IIIb exon. Intron 8 is only 46 nucleotides in length and does not have sequences that resemble IAS2 and ISAR. Given the conserved position of the intron in the C. elegans FGF-R gene and in deuterostomes, we suggest that the ancestral FGF-R gene contained an intron splitting the IgIII domain (i). A weak proto-exon IIIb evolved and was sometimes included in FGF-R transcripts (ii). Our data indicate that the common ancestor of echinoderms and vertebrates evolved the regulation of this proto-exon IIIb and developed an FGF-R IgIII alternative splicing unit with IAS2 and ISAR elements, which activate the proto-exon (indicated in green) (iii). We believe that before the evolution of vertebrates, an organism evolved the mutually exclusive use of exon IIIb or exon IIIc (iv). This unit must have evolved the capacity to repress exon IIIc inclusion (indicated in red). The IgIII alternative splicing unit of FGF-R2 has evolved further and among mammals we observed the IAS1 element immediately downstream of exon IIIb (indicated in blue)(v).








