
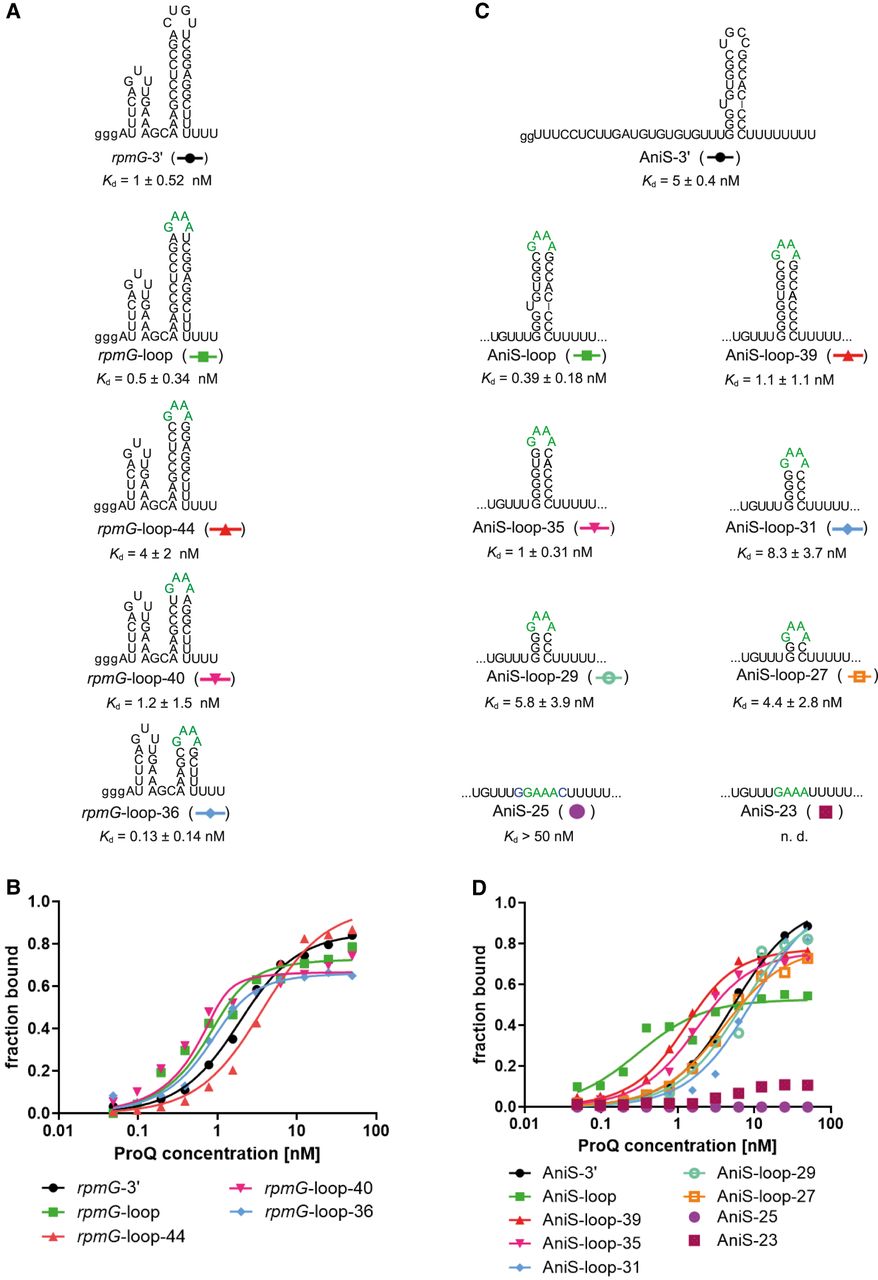
The lower part of the terminator hairpin is involved in rpmG-3′ and AniS-3′ binding to the N. meningitidis ProQ protein. (A) rpmG-3′ mutants with shorter terminator stems were constructed by replacement of the native apical loop CUGU with the tetraloop GAAA and gradual removal of base pairs from the top of the terminator stem. (B) The fitting of the ProQ binding data using the quadratic equation provided Kd values of 1.2 nM for rpmG-3′, 0.3 nM for rpmG-loop, 3.2 nM for rpmG-loop-44, 0.6 nM for rpmG-loop-40, and 0.3 nM for rpmG-loop-36. (C) AniS-3′ mutants with shorter terminator stems were constructed by replacement of the native apical loop UGCC with the tetraloop GAAA, removing the bulge, and gradual removal of base pairs from the top of the terminator stem. (D) The fitting of the ProQ binding data using the quadratic equation provided Kd values of 4.9 nM for AniS-3′, 0.7 nM for AniS-loop-39, 1.3 nM for AniS-loop-35, 9.2 nM for AniS-loop-31, 6.2 nM for AniS-loop-29, 3.5 nM for AniS-loop-27, while the binding for AniS-25 was barely detected, and AniS-23 was essentially undetected up to 50 nM concentration of the ProQ. The fitting of data for AniS-loop RNA to the equation for one site-specific binding with the Hill slope model provided Kd value of 0.1 nM. The data in the plots for rpmG-3′ and AniS-3′ binding to ProQ are the same as in Figure 1. The lower case g denotes guanosine residue added on 5′ end to enable T7 RNA polymerase transcription. Green font indicates GAAA tetraloop. Gels corresponding to the data in the plots are shown in Supplemental Figure S9. The RNA secondary structure predictions were performed in the ViennaRNA program (Lorenz et al. 2011). The average equilibrium dissociation constant (Kd) values and maximum RNA fraction bound calculated from at least three independent experiments are shown in Table 4.








