
Human T-cell leukemia virus type 1 uses a specific tRNAPro isodecoder to prime reverse transcription
- Department of Chemistry and Biochemistry, Center for RNA Biology, and Center for Retrovirus Research, The Ohio State University, Columbus, Ohio 43210, USA
- Corresponding author: musier-forsyth.1{at}osu.edu
Abstract
Human T-cell leukemia virus type 1 (HTLV-1) is the only oncogenic human retrovirus discovered to date. All retroviruses are believed to use a host cell tRNA to prime reverse transcription (RT). In HTLV-1, the primer-binding site (PBS) in the genomic RNA is complementary to the 3′ 18 nucleotides (nt) of human tRNAPro. The human genome encodes 20 cytoplasmic tRNAPro genes representing seven isodecoders, all of which share the same 3′ 18 nt sequence but vary elsewhere. Whether all tRNAPro isodecoders are used to prime RT in cells is unknown. A previous study showed that a 3′ 18 nt tRNAPro-derived fragment (tRFPro) is packaged into HTLV-1 particles and can serve as an RT primer in vitro. The role of this tRNA fragment in the viral life cycle is unclear. In retroviruses, N1-methylation of the tRNA primer at position A58 (m1A) is essential for successful plus-strand transfer. Using primer-extension assays performed in chronically HTLV-1-infected cells, we found that A58 of tRNAPro is m1A-modified, implying that full-length tRNAPro is capable of facilitating successful plus-strand transfer. Analysis of HTLV-1 RT primer extension products indicated that full-length tRNAPro is likely to be the primer. To determine which tRNAPro isodecoder is used as the RT primer, we sequenced the minus-strand strong-stop RT product containing the intact tRNA primer and established that HTLV-1 primes RT using a specific tRNAPro UGG isodecoder. Further studies are required to understand how this primer is annealed to the highly structured HTLV-1 PBS and to investigate the role of tRFPro in the viral life cycle.
Keywords
INTRODUCTION
Human T-cell leukemia virus type 1 (HTLV-1) belongs to the genus Deltaretrovirus of the family Retroviridae. This virus is the causative agent of adult T-cell leukemia/lymphoma (ATLL), an aggressive malignancy of the peripheral CD4+ T lymphocytes (Hinuma et al. 1981), and HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP), an inflammatory neurodegenerative disease (Gessain et al. 1985; Osame et al. 1986). Besides human immunodeficiency virus type 1 (HIV-1) and HIV-2, HTLV-1 is the only other species of the Retroviridae family to conclusively cause human disease. An estimated 5–10 million people are infected with HTLV-1 throughout the world, and it is endemic in the Caribbean, South America, the Middle East, sub-Saharan Africa, southern Japan, Papua New Guinea, and central Australia (Gessain and Cassar 2012). About 5% of HTLV-1-infected people develop either ATLL or HAM/TSP; higher proviral load is associated with a higher lifetime risk of developing these diseases (Nagai et al. 1998; Meekings et al. 2008; Akbarin et al. 2013). The current treatment for ATLL includes chemotherapy, biological agents, allogeneic hematopoietic stem cell transplantation (alloHSCT), and antiretroviral therapy (ART) (Panfil et al. 2016), using a combination of interferon-α and zidovudine (AZT or 3′-azido-3′-deoxythymidine) (Hermine et al. 2002; O'Donnell et al. 2024). AZT is a nucleoside analog reverse transcriptase inhibitor (NRTI), suggesting inhibiting this enzyme and virus replication could mitigate ATLL disease progression. However, many mechanistic details of reverse transcription (RT) in HTLV-1 are still unknown.
All retroviruses are believed to use host cell tRNAs as primers to initiate RT (Marquet et al. 1995). The 3′ 18 nucleotides (nt) of the tRNAs that serve as primers are perfectly complementary to a region in the 5′ UTR of
the genomic RNA (gRNA) known as the primer-binding site (PBS). For example, HIV-1, HIV-2, simian immunodeficiency virus (SIV),
feline immunodeficiency virus (FIV), mouse mammary tumor virus (MMTV), and equine infectious anemia virus (EIAV) use 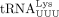 (Peters and Glover 1980; Kawakami et al. 1987; Talbott et al. 1989; Jiang et al. 1993); Mason–Pfizer monkey virus (MPMV) and human foamy virus (HFV) use
(Peters and Glover 1980; Kawakami et al. 1987; Talbott et al. 1989; Jiang et al. 1993); Mason–Pfizer monkey virus (MPMV) and human foamy virus (HFV) use 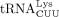 (Harrison et al. 1995; Baldwin and Linial 1999); Moloney murine leukemia virus (MoMLV), feline leukemia virus (FLV), spleen necrosis virus (SNV), simian sarcoma virus (SSV),
and bovine leukemia virus (BLV) use
(Harrison et al. 1995; Baldwin and Linial 1999); Moloney murine leukemia virus (MoMLV), feline leukemia virus (FLV), spleen necrosis virus (SNV), simian sarcoma virus (SSV),
and bovine leukemia virus (BLV) use 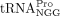 (Harada et al. 1979; Shimotohno et al. 1980; Devare et al. 1983; Laprevotte et al. 1984; Sagata et al. 1984); and Rous sarcoma virus (RSV) and avian sarcoma leukosis virus (ASLV) use
(Harada et al. 1979; Shimotohno et al. 1980; Devare et al. 1983; Laprevotte et al. 1984; Sagata et al. 1984); and Rous sarcoma virus (RSV) and avian sarcoma leukosis virus (ASLV) use 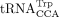 (Harada et al. 1975; Haseltine et al. 1977). In many of these cases, the identity of the tRNA RT primer was inferred from the sequence of the PBS based on complementarity
to the corresponding tRNA. In the case of RSV, MoMLV, MMTV, and HIV-1, the primer was identified via sequencing of the small
RNA released from the viral gRNA and/or other biochemical analyses, such as aminoacylation assays to measure specific amino
acid acceptor activity (Faras et al. 1974; Harada et al. 1975, 1979; Peters and Glover 1980; Jiang et al. 1993).
(Harada et al. 1975; Haseltine et al. 1977). In many of these cases, the identity of the tRNA RT primer was inferred from the sequence of the PBS based on complementarity
to the corresponding tRNA. In the case of RSV, MoMLV, MMTV, and HIV-1, the primer was identified via sequencing of the small
RNA released from the viral gRNA and/or other biochemical analyses, such as aminoacylation assays to measure specific amino
acid acceptor activity (Faras et al. 1974; Harada et al. 1975, 1979; Peters and Glover 1980; Jiang et al. 1993).
The mechanism for 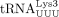 primer selection, packaging, and annealing to the HIV-1 PBS has been extensively studied (Jiang et al. 1993; Hargittai et al. 2004; Kleiman et al. 2010; Jin and Musier-Forsyth 2019). When human
primer selection, packaging, and annealing to the HIV-1 PBS has been extensively studied (Jiang et al. 1993; Hargittai et al. 2004; Kleiman et al. 2010; Jin and Musier-Forsyth 2019). When human 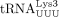 was initially identified as the HIV-1 RT primer, the convention for numbering tRNAs varied. For example,
was initially identified as the HIV-1 RT primer, the convention for numbering tRNAs varied. For example, 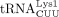 ,
, 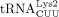 , and
, and 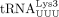 were numbered according to the order they eluted from an anion exchange column (Raba et al. 1979).
were numbered according to the order they eluted from an anion exchange column (Raba et al. 1979). 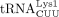 and
and 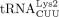 differ in a single base pair in the anticodon stem. Thus, they are often designated as
differ in a single base pair in the anticodon stem. Thus, they are often designated as 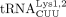 , and these two tRNA species belong to the same isoacceptor group because they share the same anticodon. The HIV-1 primer,
, and these two tRNA species belong to the same isoacceptor group because they share the same anticodon. The HIV-1 primer,
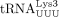 , belongs to a second isoacceptor group. Two human
, belongs to a second isoacceptor group. Two human 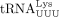 sequences encoded by six distinct genes share the same 3′ 18 nt but differ in one position (A50 or G50) (Supplemental Fig. S2B). To our knowledge whether one or both of these tRNAs serves as the HIV-1 primer has not been established. In HTLV-1 and
HTLV-2, the PBS is complementary to the 3′ 18 nt of human tRNAPro (Seiki et al. 1982). However, both full-length tRNAPro and a 3′ 18 nt tRNAPro-derived tRNA fragment (tRFPro) are enriched in HTLV-1 particles (Ruggero et al. 2014). Both RNAs contain regions perfectly complementary to the 18 nt HTLV-1 PBS and are capable of priming RT in vitro (Ruggero et al. 2014). Previous RNA-structure probing data showed that the HTLV-1 PBS is embedded in a highly stable hairpin containing 10 Watson–Crick
base pairs (Wu et al. 2018), in contrast to the HIV-1 PBS, which is much less structured (Sleiman et al. 2013). The highly structured nature of the HTLV-1 PBS raises the possibility that the less structured tRFPro primes RT in this system.
sequences encoded by six distinct genes share the same 3′ 18 nt but differ in one position (A50 or G50) (Supplemental Fig. S2B). To our knowledge whether one or both of these tRNAs serves as the HIV-1 primer has not been established. In HTLV-1 and
HTLV-2, the PBS is complementary to the 3′ 18 nt of human tRNAPro (Seiki et al. 1982). However, both full-length tRNAPro and a 3′ 18 nt tRNAPro-derived tRNA fragment (tRFPro) are enriched in HTLV-1 particles (Ruggero et al. 2014). Both RNAs contain regions perfectly complementary to the 18 nt HTLV-1 PBS and are capable of priming RT in vitro (Ruggero et al. 2014). Previous RNA-structure probing data showed that the HTLV-1 PBS is embedded in a highly stable hairpin containing 10 Watson–Crick
base pairs (Wu et al. 2018), in contrast to the HIV-1 PBS, which is much less structured (Sleiman et al. 2013). The highly structured nature of the HTLV-1 PBS raises the possibility that the less structured tRFPro primes RT in this system.
Although 20 amino acids are specified by 61 codons, more than 450 tRNA genes are encoded in the human genome (Goodenbour and Pan 2006; Chan et al. 2021). tRNAs aminoacylated with a particular amino acid are grouped into isoacceptor families by anticodon sequence; tRNAs with
the same anticodon (in the same isoacceptor family) but different body sequences are called isodecoders. According to the
genomic tRNA database (GtRNAdb), each tRNA gene is assigned a unique identification in the format: [tRNA]-[three-letter amino
acid code]-[anticodon]-[transcript ID]-[gene locus ID] (Seal et al. 2020). tRNA genes with the same transcript ID but different locus ID have the same sequence but are distributed in different genomic
locations. Each isodecoder has a unique transcript ID, with smaller numbers reflecting greater confidence that this gene encodes
a functional tRNA. Isodecoders are differentially transcribed by RNA polymerase III, resulting in varying tRNA abundance across
tissues, as well as different aminoacylation efficiencies and translation rates (Dittmar et al. 2005; Dale et al. 2009; Geslain and Pan 2010; Zhang et al. 2011; Pinkard et al. 2020). How sequence differences among a tRNA isoacceptor family contribute to their distinct functions is still not well understood.
In one recent study, a specific 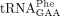 isodecoder encoded by the tRNA-Phe-GAA-1-1 gene was found to be essential for mouse brain development and embryo viability.
Loss of this isodecoder led to compensatory elevation of other
isodecoder encoded by the tRNA-Phe-GAA-1-1 gene was found to be essential for mouse brain development and embryo viability.
Loss of this isodecoder led to compensatory elevation of other 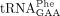 isodecoders and more misincorporation of Phe in proteins with high Phe content in mouse brain (Hughes et al. 2023).
isodecoders and more misincorporation of Phe in proteins with high Phe content in mouse brain (Hughes et al. 2023).
In this study, our main goal was to identify the specific RNA that HTLV-1 uses as an RT primer. Twenty tRNAPro genes are encoded in the human genome and represent three isoacceptor families (UGG, CGG, and AGG) and seven distinct isodecoders. All isodecoders possess identical 3′ 18 nt sequence and could, in principle, serve as the RT primer. We probed the size and sequence of the HTLV-1 RT primer in virions by primer-tagging assays and Sanger sequencing. Our results support the use of a full-length tRNA as an RT primer and revealed the specific tRNAPro isodecoder used by HTLV-1 to prime RT. We also identified the exact tRNALys isodecoder used by HIV-1. Our results suggest that both HIV-1 and HTLV-1 select specific tRNA isodecoders as their RT primers.
RESULTS
Full-length tRNAPro is the HTLV-1 RT primer
To identify the authentic RT primer used by human retroviruses, we purified the primer-PBS binary complex from HTLV-1 and
HIV-1 virions and performed primer-tagging assays to determine the length of the extended RNA. After the total viral RNA was
extracted, the endogenous primer-PBS binary complexes were formed by cofolding, as described in the Materials and Methods,
extended in the presence of [α-32P] dCTP (dC*), and terminated by ddTTP. The samples were resolved on a sequencing gel along with length standards (Supplemental Fig. S1A,B). As expected, the results obtained for the HIV-1 sample show RT primer extension products of 77 nt (+1 nt, dC*) and 78 nt
(+2 nt, dC*, and ddT), consistent with the extension of the HIV-1 RT primer, 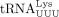 (Supplemental Fig. S1B, lane 10). Lane 11 shows the results obtained with the HTLV-1 virion samples; 77 nt (+2 nt, dA, and dC*) and 78 nt (+3 nt,
dA, dC*, and ddT) RT primer extension products were detected, consistent with full-length tRNAPro serving as the RT primer in HTLV-1 virions. No product was detected at 21 nt, which suggests that 18 nt tRFPro is not the major RT primer.
(Supplemental Fig. S1B, lane 10). Lane 11 shows the results obtained with the HTLV-1 virion samples; 77 nt (+2 nt, dA, and dC*) and 78 nt (+3 nt,
dA, dC*, and ddT) RT primer extension products were detected, consistent with full-length tRNAPro serving as the RT primer in HTLV-1 virions. No product was detected at 21 nt, which suggests that 18 nt tRFPro is not the major RT primer.
Full-length tRNAPro contains the m1A58 modification
In HIV-1, N1-methylation of A58 (m1A58) of 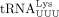 is required for the proper termination of plus-strand strong-stop DNA ([+] ssDNA) synthesis and successful plus-strand transfer
(Fig. 1A; Renda et al. 2001, 2004). Assuming a similar mechanism of RT in HTLV-1, m1A at position 58 of tRNAPro would, therefore, be required. To test whether m1A58 is indeed present in tRNAPro in HTLV-1-infected cells, total RNA was extracted from chronically HTLV-1-infected MT-2 cells, and primer extension was performed
using 5′ 32P-labeled 16 nt DNA primers complementary to the 3′ end of tRNAPro or
is required for the proper termination of plus-strand strong-stop DNA ([+] ssDNA) synthesis and successful plus-strand transfer
(Fig. 1A; Renda et al. 2001, 2004). Assuming a similar mechanism of RT in HTLV-1, m1A at position 58 of tRNAPro would, therefore, be required. To test whether m1A58 is indeed present in tRNAPro in HTLV-1-infected cells, total RNA was extracted from chronically HTLV-1-infected MT-2 cells, and primer extension was performed
using 5′ 32P-labeled 16 nt DNA primers complementary to the 3′ end of tRNAPro or 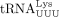 (Fig. 1B, lanes 1 and 5). The 32P-labeled primers were annealed to in vitro transcribed
(Fig. 1B, lanes 1 and 5). The 32P-labeled primers were annealed to in vitro transcribed 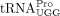 or
or 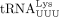 ; extension of these primers terminated at C56 as expected, based on the absence of posttranscriptional modifications (Fig. 1B, lanes 2 and 6). When primer extension was performed in the presence of cellular tRNAs, the reactions terminated at A59
(Fig. 1B, lanes 3 and 7). These data support the presence of m1A58 in both tRNAs. To confirm that the early stop at A59 was due to methylation at A58, total RNA extracted from MT-2 cells
was treated with α-ketoglutarate-dependent dioxygenase B (AlkB), a demethylase that removes the methyl group at N1 of A and
N3 of C (Zheng et al. 2015). As shown in lanes 4 and 8, the primers annealed to demethylated cellular tRNAs were extended beyond A58 and terminated
at C56. These results suggest that the early stops at A59 in lanes 3 and 7 were indeed due to the presence of m1A58 in tRNAPro and
; extension of these primers terminated at C56 as expected, based on the absence of posttranscriptional modifications (Fig. 1B, lanes 2 and 6). When primer extension was performed in the presence of cellular tRNAs, the reactions terminated at A59
(Fig. 1B, lanes 3 and 7). These data support the presence of m1A58 in both tRNAs. To confirm that the early stop at A59 was due to methylation at A58, total RNA extracted from MT-2 cells
was treated with α-ketoglutarate-dependent dioxygenase B (AlkB), a demethylase that removes the methyl group at N1 of A and
N3 of C (Zheng et al. 2015). As shown in lanes 4 and 8, the primers annealed to demethylated cellular tRNAs were extended beyond A58 and terminated
at C56. These results suggest that the early stops at A59 in lanes 3 and 7 were indeed due to the presence of m1A58 in tRNAPro and 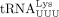 , respectively. Taken together, these results show that the A58 residue of tRNAPro in MT-2 cells is N1-methylated and suggest that tRNAPro can facilitate the proper termination of (+) ssDNA synthesis and successful plus-strand transfer.
, respectively. Taken together, these results show that the A58 residue of tRNAPro in MT-2 cells is N1-methylated and suggest that tRNAPro can facilitate the proper termination of (+) ssDNA synthesis and successful plus-strand transfer.
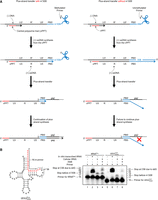
Primer extension assays to probe the methylation state of A58 of tRNAPro in MT-2 cells. (A) Plus-strand strong-stop DNA ([+] ssDNA) synthesis terminates 1 nt before A58 due to N1 methylation (m1A58); this ensures proper completion of plus-strand synthesis (left). In the absence of m1A58, (+) ssDNA synthesis terminates downstream from A58, and the product contains sequences not present in the gRNA, resulting
in failed plus-strand synthesis (right). (B) (Left) Sequence of unmodified human 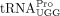 (isodecoder identified in this work as the HTLV-1 RT primer). (Right) Denaturing polyacrylamide gel showing migration of 32P-labeled DNA primers and extension products. (Lanes 1 and 5) The 5′ 32P-labeled 16 nt primers complementary to the 3′ end of tRNAPro or
(isodecoder identified in this work as the HTLV-1 RT primer). (Right) Denaturing polyacrylamide gel showing migration of 32P-labeled DNA primers and extension products. (Lanes 1 and 5) The 5′ 32P-labeled 16 nt primers complementary to the 3′ end of tRNAPro or 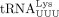 , respectively. (Lanes 2 and 6) Primer extension products with in vitro transcribed
, respectively. (Lanes 2 and 6) Primer extension products with in vitro transcribed  or
or 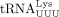 . (Lanes 3 and 7) Primer extension products with cellular tRNAPro or
. (Lanes 3 and 7) Primer extension products with cellular tRNAPro or 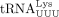 . (Lanes 4 and 8) Primer extension products using cellular tRNAPro or
. (Lanes 4 and 8) Primer extension products using cellular tRNAPro or 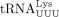 following demethylation with AlkB.
following demethylation with AlkB.
Specific 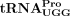 isodecoder is used as the RT primer in HTLV-1 virions
isodecoder is used as the RT primer in HTLV-1 virions
Humans encode 20 cytoplasmic tRNAPro genes that encompass three isoacceptor families, including three 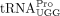 , two
, two 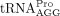 , and two
, and two 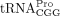 isodecoders. All cytoplasmic tRNAPro sequences share the same acceptor stem and TΨC arm, including the 3′ 18 nt that are complementary to the HTLV-1 PBS (Supplemental Fig. S2A). Thus, it is not clear based on sequence alone, whether a specific tRNA isodecoder serves as the RT primer. To answer this
question for both human retroviruses, we purified HTLV-1 and HIV-1 RNAs from culture supernatants of MT-2 cells and HEK293T
cells transfected with pNL4-3, respectively. Following two rounds of RT, as described in the Materials and Methods, the RT
product containing the primer sequence was amplified by PCR (Fig. 2A). Bands of the expected size were excised and eluted from an agarose gel, and submitted for Sanger sequencing (Fig. 2B,C). In three independent experiments, the sequence of the RT primer used by HTLV-1 aligned to tRNA-Pro-TGG-3-X (where X
is 1–5, depending on the gene locus) except at position 34, the wobble position (Fig. 2D, indicated by a red arrow). The nt at position 34 is a U in this isodecoder (http://gtrnadb.ucsc.edu/) (Chan et al. 2021), and instead of the expected A, we observed base-pairing primarily with C or with A or G at a lower frequency during the
second round RT reaction. U34 is likely modified to 5-carbamoylmethyluridine (ncm5U34) (Keith et al. 1990). This highly modified base has been observed to base pair with C through two hydrogen bonds formed between the N3 hydrogen
in the modified U and the N3 nitrogen in C, and between the 4-carbonyl oxygen in the modified U and the N4 hydrogen in C (Supplemental Fig. S3; Plant et al. 2007; Tükenmez et al. 2015).
isodecoders. All cytoplasmic tRNAPro sequences share the same acceptor stem and TΨC arm, including the 3′ 18 nt that are complementary to the HTLV-1 PBS (Supplemental Fig. S2A). Thus, it is not clear based on sequence alone, whether a specific tRNA isodecoder serves as the RT primer. To answer this
question for both human retroviruses, we purified HTLV-1 and HIV-1 RNAs from culture supernatants of MT-2 cells and HEK293T
cells transfected with pNL4-3, respectively. Following two rounds of RT, as described in the Materials and Methods, the RT
product containing the primer sequence was amplified by PCR (Fig. 2A). Bands of the expected size were excised and eluted from an agarose gel, and submitted for Sanger sequencing (Fig. 2B,C). In three independent experiments, the sequence of the RT primer used by HTLV-1 aligned to tRNA-Pro-TGG-3-X (where X
is 1–5, depending on the gene locus) except at position 34, the wobble position (Fig. 2D, indicated by a red arrow). The nt at position 34 is a U in this isodecoder (http://gtrnadb.ucsc.edu/) (Chan et al. 2021), and instead of the expected A, we observed base-pairing primarily with C or with A or G at a lower frequency during the
second round RT reaction. U34 is likely modified to 5-carbamoylmethyluridine (ncm5U34) (Keith et al. 1990). This highly modified base has been observed to base pair with C through two hydrogen bonds formed between the N3 hydrogen
in the modified U and the N3 nitrogen in C, and between the 4-carbonyl oxygen in the modified U and the N4 hydrogen in C (Supplemental Fig. S3; Plant et al. 2007; Tükenmez et al. 2015).
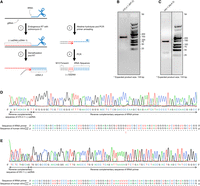
Sequencing minus-strand strong-stop DNA ([−] ssDNA) to identify sequence of HTLV-1 and HIV-1 RT primers. (A) Flowchart of method used to amplify sequence containing the HIV-1 and HTLV-1 RT primer with a short RT extension. Endogenous
RT was performed to synthesize the (−) ssDNA (step 1). Following demethylation, a second round of RT was performed to synthesize
the antisense strand of the tRNA RT primer (step 2). The product was PCR-amplified (steps 3 and 4). (B,C) The PCR-amplified products were run on 2% agarose gels. PCR products with the expected size (boxed with red lines, 135 bp
for HTLV-1, B; 133 bp for HIV-1, C) were excised and eluted from the gel and submitted for Sanger sequencing. (D,E) Chromatograph of Sanger sequencing results for the (−) ssDNA containing RT primer and sequence alignments to 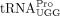 (for HTLV-1, D) or
(for HTLV-1, D) or 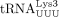 (for HIV-1, E).
(for HIV-1, E).
We used similar methods to confirm the sequence of the RT primer used by HIV-1. Humans encode 27 cytoplasmic tRNALys genes, which are members of two isoacceptor families, including eight 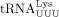 and 10
and 10 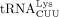 isodecoders. Among them, only tRNA-Lys-TTT-3-X (where X is 1–5) and tRNA-Lys-TTT-5-1 genes share the same 3′ 18 nt sequence
that is complementary to the HIV-1 PBS. The only difference between these two isodecoders is at position 50, which is an A
in tRNA-Lys-TTT-3-X and a G in tRNA-Lys-TTT-3-X (Supplemental Fig. S2B). We found that HIV-1 uses the specific isodecoder encoded by tRNA-Lys-TTT-3-X as the RT primer (Fig. 2E). In this case, heterogeneous base-pairing with U34 was not observed and U34 was unambiguously identified. Uridine at the
wobble position in
isodecoders. Among them, only tRNA-Lys-TTT-3-X (where X is 1–5) and tRNA-Lys-TTT-5-1 genes share the same 3′ 18 nt sequence
that is complementary to the HIV-1 PBS. The only difference between these two isodecoders is at position 50, which is an A
in tRNA-Lys-TTT-3-X and a G in tRNA-Lys-TTT-3-X (Supplemental Fig. S2B). We found that HIV-1 uses the specific isodecoder encoded by tRNA-Lys-TTT-3-X as the RT primer (Fig. 2E). In this case, heterogeneous base-pairing with U34 was not observed and U34 was unambiguously identified. Uridine at the
wobble position in 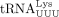 is modified to 5-methoxycarbonylmethyl-2-thiouridine (mcm5s2U34) (Björk et al. 2007) and is critical for mRNA decoding and translation (Johansson et al. 2008; Klassen et al. 2015). The terminal methyl group in mcm5U34 may be removed by AlkB treatment before the second round of RT, resulting in 5-carboxymethyl-2-thiouridine (cm5s2U34) (Supplemental Fig. S3B). The less bulky nature of this modification relative to ncm5U may prevent mispairing.
is modified to 5-methoxycarbonylmethyl-2-thiouridine (mcm5s2U34) (Björk et al. 2007) and is critical for mRNA decoding and translation (Johansson et al. 2008; Klassen et al. 2015). The terminal methyl group in mcm5U34 may be removed by AlkB treatment before the second round of RT, resulting in 5-carboxymethyl-2-thiouridine (cm5s2U34) (Supplemental Fig. S3B). The less bulky nature of this modification relative to ncm5U may prevent mispairing.
DISCUSSION
In a previous study, both full-length tRNAPro and an 18 nt 3′ fragment of tRNAPro (tRFPro) were shown to be packaged into HTLV-1 virions and both were capable of priming RT in vitro (Ruggero et al. 2014). In this work, full-length 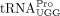 was identified as the RT primer, in agreement with other retroviruses that also use full-length tRNAs as RT primers (Marquet et al. 1995). The HTLV-1 PBS forms part of a stable hairpin containing 10 bp (Wu et al. 2018), which is predicted to be more stable than the HIV-1 PBS hairpin with only 3 bp (Sleiman et al. 2013). Although annealing of tRNAPro is thermodynamically disfavored compared to tRFPro, annealing reactions requiring higher activation energy and multiple steps could be more tightly regulated than reactions
with lower activation energy. In HIV-1, besides the base-pairing between the PBS and the 3′ 18 nt of
was identified as the RT primer, in agreement with other retroviruses that also use full-length tRNAs as RT primers (Marquet et al. 1995). The HTLV-1 PBS forms part of a stable hairpin containing 10 bp (Wu et al. 2018), which is predicted to be more stable than the HIV-1 PBS hairpin with only 3 bp (Sleiman et al. 2013). Although annealing of tRNAPro is thermodynamically disfavored compared to tRFPro, annealing reactions requiring higher activation energy and multiple steps could be more tightly regulated than reactions
with lower activation energy. In HIV-1, besides the base-pairing between the PBS and the 3′ 18 nt of 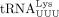 , efficient initiation of RT has been reported to require a loop–loop interaction between the A-rich loop upstream of the
PBS (MAL isolate) and the U-rich anticodon loop of
, efficient initiation of RT has been reported to require a loop–loop interaction between the A-rich loop upstream of the
PBS (MAL isolate) and the U-rich anticodon loop of 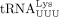 (Isel et al. 1993), an interaction between a C-rich region upstream of the A-rich loop (NL4-3 isolate) and the G-rich 3′ anticodon stem and
variable loop of
(Isel et al. 1993), an interaction between a C-rich region upstream of the A-rich loop (NL4-3 isolate) and the G-rich 3′ anticodon stem and
variable loop of 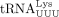 (Iwatani et al. 2003), and an interaction between the primer activation signal (PAS) upstream of the C-rich region (LAI isolate) and the 5′ TψC
arm of
(Iwatani et al. 2003), and an interaction between the primer activation signal (PAS) upstream of the C-rich region (LAI isolate) and the 5′ TψC
arm of 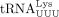 (Beerens and Berkhout 2002b; Sleiman et al. 2012). In HTLV-1, a PAS sequence 10 nt upstream of the PBS has been predicted (Beerens and Berkhout 2002a). Whether this PAS sequence plays a role in RT and whether there are other interactions between
(Beerens and Berkhout 2002b; Sleiman et al. 2012). In HTLV-1, a PAS sequence 10 nt upstream of the PBS has been predicted (Beerens and Berkhout 2002a). Whether this PAS sequence plays a role in RT and whether there are other interactions between 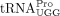 and the HTLV-1 5′ UTR outside the 18 nt PBS domain are unknown.
and the HTLV-1 5′ UTR outside the 18 nt PBS domain are unknown.
In primer extension assays, m1A58 was detected in tRNAPro isolated from MT-2 cells (Fig. 1B), consistent with the capability of tRNAPro to facilitate plus-strand transfer during RT. In addition to m1A58, other posttranscriptional modifications in 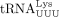 are required for optimal RT in HIV-1, such as mcm5s2U34 at the wobble position and 5,2′-O-dimethyluridine at U54 (m5Um54) (Isel et al. 1993; Fukuda et al. 2021). Modification of mcm5s2 at U34 maintains the canonical U-turn anticodon-loop structure and enhances the interaction between the A-rich loop in HIV-1
gRNA and the U-rich anticodon loop of
are required for optimal RT in HIV-1, such as mcm5s2U34 at the wobble position and 5,2′-O-dimethyluridine at U54 (m5Um54) (Isel et al. 1993; Fukuda et al. 2021). Modification of mcm5s2 at U34 maintains the canonical U-turn anticodon-loop structure and enhances the interaction between the A-rich loop in HIV-1
gRNA and the U-rich anticodon loop of 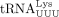 (Isel et al. 1993; Bénas et al. 2000; Sundaram et al. 2000). Whereas m1A58 acts as the first termination site for (+) ssDNA synthesis (Renda et al. 2001, 2004), m5Um54 serves as an alternative termination site when m1A58 modification is absent (Fukuda et al. 2021). Whether posttranscriptional modifications other than m1A58 in tRNAPro are essential for primer selectivity and successful RT and whether HTLV-1 infection induces tRNAPro modifications required for optimal tRNAPro-HTLV-1 gRNA interaction requires further investigation.
(Isel et al. 1993; Bénas et al. 2000; Sundaram et al. 2000). Whereas m1A58 acts as the first termination site for (+) ssDNA synthesis (Renda et al. 2001, 2004), m5Um54 serves as an alternative termination site when m1A58 modification is absent (Fukuda et al. 2021). Whether posttranscriptional modifications other than m1A58 in tRNAPro are essential for primer selectivity and successful RT and whether HTLV-1 infection induces tRNAPro modifications required for optimal tRNAPro-HTLV-1 gRNA interaction requires further investigation.
In this study, we sequenced HTLV-1 and HIV-1 RT primers following extension of primer/template complexes isolated from virions.
We found that 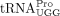 encoded by tRNA-Pro-TGG-3 serves as the HTLV-1 RT primer. The other six tRNAPro isodecoders with identical 3′ sequences complementary to the PBS were not identified in this analysis. Whereas a specific
isodecoder appears to serve as the primer in HTLV-1, tRNAPro also primes MoMLV RT, and two isoacceptors were identified to serve as primers of this simple retrovirus (Harada et al. 1979). We also found that
encoded by tRNA-Pro-TGG-3 serves as the HTLV-1 RT primer. The other six tRNAPro isodecoders with identical 3′ sequences complementary to the PBS were not identified in this analysis. Whereas a specific
isodecoder appears to serve as the primer in HTLV-1, tRNAPro also primes MoMLV RT, and two isoacceptors were identified to serve as primers of this simple retrovirus (Harada et al. 1979). We also found that 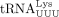 encoded by tRNA-Lys-TTT-3 serves as the HIV-1 RT primer. We did not detect the tRNALys encoded by tRNA-Lys-TTT-5 as a primer despite the fact that this tRNA is identical to tRNA-Lys-TTT-3 except at position 50
(Supplemental Fig. S2B). The sequence of tRNA-Lys-TTT-3 is the same as the previously defined tRNALys3 (Raba et al. 1979). Coincidentally, both are designated by the number three, but the meaning of this number is different. Historically, tRNALys1,2,3 were identified as three major tRNALys species in rabbit liver, where the numbering indicates the order of elution from a DEAE-Sephadex A-50 column (Raba et al. 1979). In the new gene designation, the number reflects the transcript ID (Chan et al. 2021).
encoded by tRNA-Lys-TTT-3 serves as the HIV-1 RT primer. We did not detect the tRNALys encoded by tRNA-Lys-TTT-5 as a primer despite the fact that this tRNA is identical to tRNA-Lys-TTT-3 except at position 50
(Supplemental Fig. S2B). The sequence of tRNA-Lys-TTT-3 is the same as the previously defined tRNALys3 (Raba et al. 1979). Coincidentally, both are designated by the number three, but the meaning of this number is different. Historically, tRNALys1,2,3 were identified as three major tRNALys species in rabbit liver, where the numbering indicates the order of elution from a DEAE-Sephadex A-50 column (Raba et al. 1979). In the new gene designation, the number reflects the transcript ID (Chan et al. 2021).
Previous studies showed that HIV-1 has a strong preference for maintaining the PBS sequence complementary to 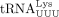 (Das et al. 1995). Mismatches of a few nucleotides are tolerated by reverse transcriptase; therefore, a misaligned tRNA primer can still be
extended (Chan and Musier-Forsyth 1997). For example, a mutant PBS sequence complementary to tRNAPro is primed by tRNAIle despite some mismatches and reverts to the tRNAIle-specific PBS after one round of HIV-1 replication (Das et al. 1995). In subsequent rounds, the tRNAIle-specific PBS sequence is primed by
(Das et al. 1995). Mismatches of a few nucleotides are tolerated by reverse transcriptase; therefore, a misaligned tRNA primer can still be
extended (Chan and Musier-Forsyth 1997). For example, a mutant PBS sequence complementary to tRNAPro is primed by tRNAIle despite some mismatches and reverts to the tRNAIle-specific PBS after one round of HIV-1 replication (Das et al. 1995). In subsequent rounds, the tRNAIle-specific PBS sequence is primed by 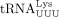 and finally reverts back to the
and finally reverts back to the 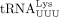 -specific PBS, which is stably maintained. This suggests that specific isodecoder tRNA selection is not just based on sequence
complementarity to the PBS; specific viral and host factors are also involved in selection. For example, human lysyl-tRNA
synthetase (LysRS), which binds specifically to all tRNALys isoacceptors, is packaged into HIV-1 virions, and tryptophanyl-tRNA synthetase (TrpRS) is packaged into RSV virions, which
use
-specific PBS, which is stably maintained. This suggests that specific isodecoder tRNA selection is not just based on sequence
complementarity to the PBS; specific viral and host factors are also involved in selection. For example, human lysyl-tRNA
synthetase (LysRS), which binds specifically to all tRNALys isoacceptors, is packaged into HIV-1 virions, and tryptophanyl-tRNA synthetase (TrpRS) is packaged into RSV virions, which
use 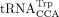 as the RT primer (Cen et al. 2002). In contrast, prolyl-tRNA synthetase (ProRS) is not packaged into MLV virions (Cen et al. 2002), and MLV has lower tRNA primer specificity than HIV-1 or RSV. When the wild-type (WT) tRNAPro-specific PBS is mutated to be complementary to tRNALys3 or tRNAGln1,2, MLV uses tRNALys3 and tRNAGln1,2 as RT primers with comparable efficiency as the use of tRNAPro by WT MLV (Colicelli and Goff 1986; Lund et al. 1993). How
as the RT primer (Cen et al. 2002). In contrast, prolyl-tRNA synthetase (ProRS) is not packaged into MLV virions (Cen et al. 2002), and MLV has lower tRNA primer specificity than HIV-1 or RSV. When the wild-type (WT) tRNAPro-specific PBS is mutated to be complementary to tRNALys3 or tRNAGln1,2, MLV uses tRNALys3 and tRNAGln1,2 as RT primers with comparable efficiency as the use of tRNAPro by WT MLV (Colicelli and Goff 1986; Lund et al. 1993). How 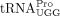 is selected as an HTLV-1 RT primer is an open question. However, selection may in part be based on tRNA abundance. Among
all three tRNAPro isoacceptor families expressed in HEK293T cells, 87.7% are
is selected as an HTLV-1 RT primer is an open question. However, selection may in part be based on tRNA abundance. Among
all three tRNAPro isoacceptor families expressed in HEK293T cells, 87.7% are 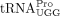 , with the major sequence (81%) being the isodecoder identified as the HTLV-1 primer in this study. Although
, with the major sequence (81%) being the isodecoder identified as the HTLV-1 primer in this study. Although 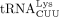 is the most abundant tRNALys isoacceptor family, among
is the most abundant tRNALys isoacceptor family, among 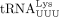 isoacceptors, 70% are
isoacceptors, 70% are 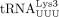 , the HIV-1 RT primer (Gogakos et al. 2017; Shigematsu et al. 2017).
, the HIV-1 RT primer (Gogakos et al. 2017; Shigematsu et al. 2017).
Our data suggest that the packaged tRFPro does not serve as the primer in HTLV-1. In HIV-1-infected cells, the tRNA/PBS duplex was proposed to be capable of being
recognized and cleaved by Dicer to produce 18 nt 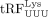 ; this tRF can be bound by Argonaute 2 (AGO2), targeting HIV-1 gRNA for degradation resulting in decreased viral replication
(Yeung et al. 2009). In mouse endogenous retroviruses (ERVs), 18 nt
; this tRF can be bound by Argonaute 2 (AGO2), targeting HIV-1 gRNA for degradation resulting in decreased viral replication
(Yeung et al. 2009). In mouse endogenous retroviruses (ERVs), 18 nt 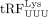 inhibits RT by competing with full-length
inhibits RT by competing with full-length 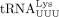 for binding to the PBS independent of AGOs, which results in restricted mobility of the transposable element (TE). This is
proposed to serve as a defense mechanism to maintain genome integrity in cells that are unable to silence the TE by epigenetic
modification (Schorn et al. 2017). Whether tRFPro serves as a facilitator or inhibitor of HTLV-1 replication remains to be investigated.
for binding to the PBS independent of AGOs, which results in restricted mobility of the transposable element (TE). This is
proposed to serve as a defense mechanism to maintain genome integrity in cells that are unable to silence the TE by epigenetic
modification (Schorn et al. 2017). Whether tRFPro serves as a facilitator or inhibitor of HTLV-1 replication remains to be investigated.
Our finding that full-length 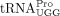 serves as the HTLV-1 RT primer raises additional questions. Because the HTLV-1 PBS forms a highly structured GC-rich hairpin
(Wu et al. 2018), a strong chaperone protein would be required to facilitate tRNA annealing to the PBS. In HIV-1, the NC domain of Gag is
a robust nucleic acid chaperone protein capable of facilitating human
serves as the HTLV-1 RT primer raises additional questions. Because the HTLV-1 PBS forms a highly structured GC-rich hairpin
(Wu et al. 2018), a strong chaperone protein would be required to facilitate tRNA annealing to the PBS. In HIV-1, the NC domain of Gag is
a robust nucleic acid chaperone protein capable of facilitating human 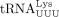 annealing to the HIV-1 PBS in the absence of any other cofactors (Hargittai et al. 2004). In contrast, HTLV-1 NC is a relatively weak chaperone protein because of its unique highly acidic C-terminal extension
(Stewart-Maynard et al. 2008; Qualley et al. 2010). The MA domain has been shown to possess more robust chaperone activity than NC in deltaretroviruses, such as HTLV-1, HTLV-2,
and BLV (Katoh et al. 1991; Sun et al. 2014; Wu et al. 2018). Future studies are required to establish whether a viral or host factor facilitates primer
annealing to the HIV-1 PBS in the absence of any other cofactors (Hargittai et al. 2004). In contrast, HTLV-1 NC is a relatively weak chaperone protein because of its unique highly acidic C-terminal extension
(Stewart-Maynard et al. 2008; Qualley et al. 2010). The MA domain has been shown to possess more robust chaperone activity than NC in deltaretroviruses, such as HTLV-1, HTLV-2,
and BLV (Katoh et al. 1991; Sun et al. 2014; Wu et al. 2018). Future studies are required to establish whether a viral or host factor facilitates primer 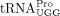 annealing onto the PBS. Taken together, the identification of the specific RT primer used by HTLV-1 may lead to future therapeutic
strategies aimed at blocking tRNA primer function and HTLV-1 RT.
annealing onto the PBS. Taken together, the identification of the specific RT primer used by HTLV-1 may lead to future therapeutic
strategies aimed at blocking tRNA primer function and HTLV-1 RT.
MATERIALS AND METHODS
Plasmids
To prepare the plasmid template for transcribing a 95 nt portion (nt 362–456) of the HTLV-1 5′ UTR containing the PBS sequence,
PCR amplification was carried out using a forward primer encoding a KpnI site and the T7 promoter and a reverse primer encoding
a PstI site (forward primer: 5′-CGT GGT ACC TAA TAC GAC TCA CTA TAG GGC CCA TCC TAT AGC ACT CTC CAG-3′; reverse primer: 5′-ATG GTC CTG CAG GGA TGT CGA GTA ACG GGC CCA TTG CCT AGG GAA TAA AGG-3′). The product was digested with KpnI and PstI and cloned into pUC19
(New England Biolabs). Plasmids for in vitro transcribing human 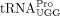 (pUC119 tRNAPro) and
(pUC119 tRNAPro) and 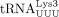 (pIDTSmart tRNALys) were generated previously (An et al. 2008; Cantara et al. 2022). Bacterial expression plasmids for purifying Escherichia coli AlkB, pET30a WT AlkB, and pET30a D135S AlkB, were gifts from Dr. Tao Pan (University of Chicago) (Zheng et al. 2015).
(pIDTSmart tRNALys) were generated previously (An et al. 2008; Cantara et al. 2022). Bacterial expression plasmids for purifying Escherichia coli AlkB, pET30a WT AlkB, and pET30a D135S AlkB, were gifts from Dr. Tao Pan (University of Chicago) (Zheng et al. 2015).
Cell culture and virus production
Human embryonic kidney cells (HEK293T) were maintained in Dulbecco's modified Eagle medium (DMEM) supplemented with 10% (v/v) fetal bovine serum (FBS), 100 unit/mL penicillin, 100 μg/mL streptomycin, and 1× nonessential amino acids solution (complete DMEM). MT-2 cells were grown in Roswell Park Memorial Institute (RPMI) 1640 medium supplemented with 10% FBS, 100 unit/mL penicillin, and 100 μg/mL streptomycin.
For these studies, strategies that allowed the preparation of large amounts of viral particles were needed. Many chronically HTLV-1-infected cell lines exist that closely mimic the natural infection. We chose MT-2 to perform our experiments, because MT-2 cells produce the highest number of virions compared to other chronically HTLV-1-infected cell lines. Chronically infected HIV-1 cell lines do not exist; therefore, we chose HEK293T cells transfected with an HIV molecular clone.
To produce HIV-1 virions, HEK293T producer cells were cotransfected with pNL4-3 E−R+Luc+ (NIH AIDS reagent program) and pMD2.G (encoding VSV-G) by the polyethylenimine (PEI) method (Longo et al. 2013). The medium was replaced with fresh complete DMEM 6 h posttransfection. Two days posttransfection, viral supernatant was collected and filtered through 0.45 μm-pore-size filters. Filtered supernatant (10 mL) layered on top of 1 mL of 25% sucrose was concentrated by centrifugation at 90,000g for 1.5 h at 4°C in a Sorvall SW41 swinging bucket rotor (sucrose cushion). The supernatant was discarded and the viral particle-containing pellet was resuspended in 500 μL of 1× STE buffer (10 mM Tris-HCl, pH 8.0, 100 mM NaCl, and 1 mM ethylenediaminetetraacetic acid [EDTA]) by gentle shaking overnight at 4°C. Resuspended viruses were further purified via an OptiPrep density gradient (Sigma-Aldrich) ranging from 10% to 40% with five steps in 7.5% increments and centrifuged at 250,000g for 3 h at 4°C in a Sorvall SW41 swinging bucket rotor. The gradient fraction (500 μL) at ∼20% was collected and concentrated using a sucrose cushion, as described above. To purify HTLV-1 virions, the MT-2 cell culture supernatant was collected, and virions were concentrated by a sucrose cushion and a 10%–40% OptiPrep density gradient using the same procedure as for purifying HIV-1 virions.
RNA preparation
Human tRFPro was purchased from Dharmacon. All other RNAs (sequences shown in Supplemental Table S1) were in vitro transcribed from linearized T7 promoter–containing plasmid templates using T7 RNA polymerase, as previously
described (Milligan et al. 1987). Briefly, in vitro transcribed RNAs were resolved on 10% polyacrylamide/8 M urea gels, eluted from the gels overnight in
RNA elution buffer (0.5 mM NH4OAc and 1 mM EDTA, pH 8.0), followed by butanol extraction to reduce the volume and ethanol precipitation. The concentration
of RNAs was determined by measuring the absorbance at 260 nm using the following molar extinction coefficients: HTLV-1 PBS,
870,278 M−1cm−1; tRFPro, 122,678 M−1cm−1; 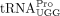 , 655,343 M−1cm−1;
, 655,343 M−1cm−1; 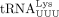 , 664,688 M−1cm−1.
, 664,688 M−1cm−1.
All RNAs were folded before use in 50 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), pH 7.5, by heating for 2 min at 80°C, for 2 min at 60°C, adding 1 mM of MgCl2, followed by incubation for 30 min at 37°C, and finally cooling on ice for at least 30 min.
5′ 32P RNA or DNA labeling
Before labeling, 500 pmol of in vitro transcribed RNA was treated with 5 units of calf intestinal alkaline phosphatase (CIP) (New England Biolabs) to remove the 5′ phosphate. After phenol–chloroform extraction and ethanol precipitation, samples were incubated with 50 μCi of [γ-32P] ATP (PerkinElmer) and 10 units of T4 polynucleotide kinase (New England Biolabs) to phosphorylate the 5′ end. Free 32P-ATP was removed using a Sephadex G-25 spin column (Roche) and samples were phenol–chloroform extracted, followed by ethanol precipitation. The CIP step was omitted for commercially synthesized nucleic acids.
Primer extension to probe the presence of m1A58 in human tRNAPro and 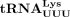
Total RNA from MT-2 cells was extracted by TRIzol (Invitrogen) and loaded onto a 12% polyacrylamide/8 M urea denaturing gel.
RNAs in the length range of 70–80 nt were excised from the gel and eluted by the crush and soak method (Milligan et al. 1987). The AlkB demethylation reaction was performed as described previously with some alterations (Zheng et al. 2015). Briefly, 1 μg of gel-extracted total tRNA was treated with 160 pmol of WT AlkB and 200 pmol of D135S AlkB, and incubated
in 50 mM MES, 300 mM KCl, 2 mM MgCl2, 50 μM of (NH4)2Fe(SO4)2·6H2O, 300 μM 2-ketoglutarate, 2 mM l-ascorbic acid, and 50 μg/mL bovine serum albumin, pH 5.0, for 2 h at room temperature. After the demethylation reaction,
tRNAs were recovered by phenol–chloroform extraction and ethanol precipitation. For primer extension reactions, 5′ 32P-labeled primers (for tRNAPro, 5′-TGG GGG CTC GTC CGG G-3′; for 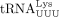 , 5′-TGG CGC CCG AAC AGG G-3′) were annealed to the 3′ 16 nt of in vitro transcribed
, 5′-TGG CGC CCG AAC AGG G-3′) were annealed to the 3′ 16 nt of in vitro transcribed 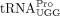 or
or 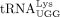 or cellular tRNA with or without AlkB treatment, and extended by incubation with d/ddNTP mix (500 nM of each dATP, dTTP,
dCTP, and ddGTP), 10 mM dithiothreitol (DTT), 1× first-strand buffer (50 mM Tri-HCl, pH 8.3, 75 mM KCl, and 3 mM MgCl2), and SuperScript III reverse transcriptase (Invitrogen) for 1 h at 50°C, followed by 15 min of incubation at 70°C to inactivate
the enzyme. After primer extension, tRNAs were removed by alkaline hydrolysis in 200 nM NaOH for 3 min at 95°C. All samples
were mixed with loading dye containing 50% formamide and run on 20% polyacrylamide/8 M urea gels. The gels were exposed to
phosphor screens overnight, and bands were visualized by phosphoimaging on a Typhoon FLA9500 (Cytiva).
or cellular tRNA with or without AlkB treatment, and extended by incubation with d/ddNTP mix (500 nM of each dATP, dTTP,
dCTP, and ddGTP), 10 mM dithiothreitol (DTT), 1× first-strand buffer (50 mM Tri-HCl, pH 8.3, 75 mM KCl, and 3 mM MgCl2), and SuperScript III reverse transcriptase (Invitrogen) for 1 h at 50°C, followed by 15 min of incubation at 70°C to inactivate
the enzyme. After primer extension, tRNAs were removed by alkaline hydrolysis in 200 nM NaOH for 3 min at 95°C. All samples
were mixed with loading dye containing 50% formamide and run on 20% polyacrylamide/8 M urea gels. The gels were exposed to
phosphor screens overnight, and bands were visualized by phosphoimaging on a Typhoon FLA9500 (Cytiva).
Sequencing the minus-strand strong-stop RT product
After purifying viruses from 8 mL culture supernatant by sucrose cushion and OptiPrep density gradient centrifugation, as described above, pelleted viruses were suspended in 40 μL of TNEP buffer (100 mM Tris-HCl, pH 8.3, 100 mM NaCl, 1 mM EDTA, and 0.01% NP-40), and endogenous RT reactions to synthesize the (−) ssDNA were initiated by adding 100 μL of 50 mM Tri-HCl, pH 8.3, 50 mM NaCl, 6 mM MgCl2, 0.01% NP-40, 1 mM DTT, 1 mM dNTP, and 100 μg/mL actinomycin D. The reaction mixtures were incubated for 30 min at 37°C and terminated by adding 20 μL of 10% sodium dodecyl sulfate (SDS) and 4 μL of 0.5 M EDTA (Tanese et al. 1991). Following phenol–chloroform extraction and ethanol precipitation, the AlkB demethylation reaction was performed as described previously (Zheng et al. 2015). To synthesize the plus-strand containing the antisense sequence of the tRNA primer, the (−) ss RT products underwent another round of RT using SuperScript III reverse transcriptase and an RT primer complementary to the 5′ end of the (−) ssDNA and containing an M13 sequence overhang (Supplemental Table S2). RNAs were removed by treatment with 200 nM NaOH at 95°C for 3 min (alkaline hydrolysis). PCR was performed using a forward primer with the same sequence as the plus-strand RT primer and a reverse primer complementary to the 5′ end of tRNA (Fig. 2A; Supplemental Table S2). PCR products were loaded onto 2% agarose gels, and bands with the expected size were excised from the gel, eluted by QIAquick gel extraction kit (QIAGEN), and submitted for Sanger sequencing at the Genomics Shared Resource Facility (Ohio State University). All human tRNA sequences used in the sequence alignments are from the GtRNAdb (http://gtrnadb.ucsc.edu/) (Chan et al. 2021).
SUPPLEMENTAL MATERIAL
Supplemental material is available for this article.
ACKNOWLEDGMENTS
We thank Dr. Patrick L. Green and Dr. Amanda R. Panfil (Ohio State University) for providing MT-2 cells and sharing space to perform cell culture work. Research reported in this manuscript was supported by the National Institute of Allergy and Infectious Disease of the National Institutes of Health (NIH) under award numbers R01AI150493 and U54AI170855. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Footnotes
-
Handling editor: Eric Westhof
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.080006.124.
-
Freely available online through the RNA Open Access option.
- Received February 24, 2024.
- Accepted April 8, 2024.
This article, published in RNA, is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








