
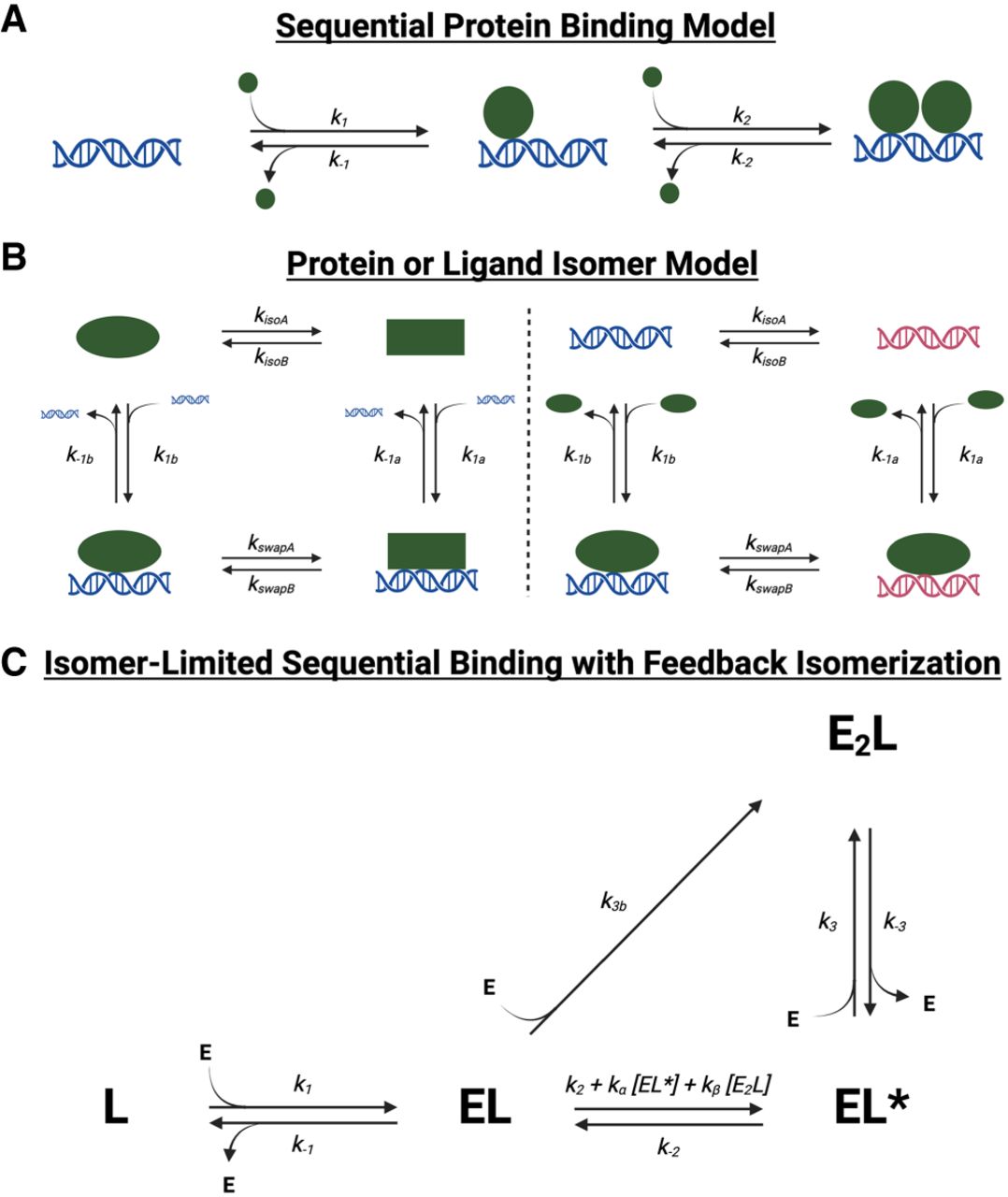
Various reaction schemes that predict biphasic protein–ligand dissociation. (A) Sequential protein-binding model. After initial protein–ligand association, additional protein can associate with the complex to form higher stoichiometry complexes. If the complex states with differing stoichiometries also have differing stabilities, multiphasic dissociation kinetics can be produced. If protein associates at differing rates with ligand versus existing complex, multiphasic association kinetics can be produced. Based on this model, rate constants for Sox2HMG CBS dsDNA binding are inferred to be k1,2 ≈ 8 × 105 M−1 sec−1, k−1 ≈ 1 × 10−3 sec−1, and k−2 ≈ 5 × 10−2 sec−1. The rate constants for Sox2HMG G4 RNA binding are inferred to be k1 ≈ 1 × 106 M−1 sec−1, k2 ≈ 7 × 104 M−1 sec−1, k−1 ≈ 1 × 10−3 sec−1, and k−2 ≈ 5 × 10−2 sec−1. (B) Protein or ligand isomer model. The protein (left) or ligand (right) may isomerize to an alternative state, which produces different protein–ligand association and/or dissociation rates. (C) Isomer-limited sequential binding with feedback isomerization. After initial protein–ligand association to form a stable complex (EL), protein can inefficiently associate with the initial complex to form a higher-order stoichiometry complex (E2L), or the initial complex can isomerize to an alternative complex state (EL*) that can more readily accommodate additional protein monomers. Complex isomerization is intrinsically slow but may be accelerated by “feedback” from isomerized complex or higher-order stoichiometry complex. Such a reaction scheme could produce monophasic, biphasic, or “lagged” triphasic association and monophasic, biphasic, or triphasic dissociation, depending on specific values of rate constants. E is a monomer of free protein, and L is free ligand.








