
Multifaceted roles of RNA editing enzyme ADAR1 in innate immunity
- 1Department of Genetics, Stanford University, Stanford, California 94305, USA
- 2AIRNA Corporation, Cambridge, Massachusetts 02142, USA
- Corresponding authors: inga.jarmoskaite{at}airna.com, jin.billy.li{at}stanford.edu
Abstract
Innate immunity must be tightly regulated to enable sensitive pathogen detection while averting autoimmunity triggered by pathogen-like host molecules. A hallmark of viral infection, double-stranded RNAs (dsRNAs) are also abundantly encoded in mammalian genomes, necessitating surveillance mechanisms to distinguish “self” from “nonself.” ADAR1, an RNA editing enzyme, has emerged as an essential safeguard against dsRNA-induced autoimmunity. By converting adenosines to inosines (A-to-I) in long dsRNAs, ADAR1 covalently marks endogenous dsRNAs, thereby blocking the activation of the cytoplasmic dsRNA sensor MDA5. Moreover, beyond its editing function, ADAR1 binding to dsRNA impedes the activation of innate immune sensors PKR and ZBP1. Recent landmark studies underscore the utility of silencing ADAR1 for cancer immunotherapy, by exploiting the ADAR1-dependence developed by certain tumors to unleash an antitumor immune response. In this perspective, we summarize the genetic and mechanistic evidence for ADAR1's multipronged role in suppressing dsRNA-mediated autoimmunity and explore the evolving roles of ADAR1 as an immuno-oncology target.
Keywords
INTRODUCTION
Innate immune pathways that sense double-stranded RNA (dsRNA) form an essential layer of antiviral defense in mammals (Fig. 1; Hur 2019). However, besides signaling viral infection, dsRNAs can also form within endogenous transcripts, predominantly between the transposable repeat sequences that occupy nearly half of the human genome. Thus, the protective role of dsRNA sensors needs to be constantly balanced with the risk of erroneously launching an immune response against cellular RNAs (Chen and Hur 2022).
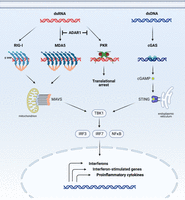
Major cytoplasmic nucleic sensing pathways in mammals. The RIG-I pathway (activated by 5′-triphosphorylated, blunt-ended dsRNA), MDA5 pathway (activated by long dsRNAs), and cGAS-STING pathway (activated by dsDNA) converge to induce interferon signaling. Meanwhile, PKR dimerization on dsRNA primarily leads to translational arrest and cell death. ADAR1 blocks the recognition of endogenous dsRNAs by MDA5 and PKR to prevent constitutive interferon activation and apoptosis.
Adenosine-to-inosine (A-to-I) RNA editing constitutes an essential mechanism for marking cellular dsRNAs as “self” and repressing autoimmunity (Quin et al. 2021). In mammals, A-to-I RNA editing is carried out by two catalytically active ADAR proteins, ADAR1 and ADAR2. A third homolog, ADAR3, lacks deaminase activity and instead functions as a negative regulator of editing in the brain (Fig. 2A; Nishikura 2016; Oakes et al. 2017; Tan et al. 2017). The ubiquitously expressed ADAR1 protein carries out most of the cellular editing, predominantly targeting long dsRNAs that form between the complementary sequences of transposable repeat elements (Tan et al. 2017). Meanwhile, ADAR2 primarily functions in the brain and is responsible for most A-to-I editing events in coding regions, including an essential recoding event in the neurotransmitter receptor GRIA2 (Sommer et al. 1991; Higuchi et al. 2000; Tan et al. 2017). ADAR1 comes in two isoforms that exhibit significant differences in molecular structure, subcellular localization, and function. The shorter p110 isoform is expressed from a constitutively active promoter and resides in the nucleus, whereas the longer p150 isoform, featuring an additional nucleic acid binding domain, initiates from an upstream interferon-inducible promoter and shuttles between the nucleus and cytoplasm (George and Samuel 1999; Sun et al. 2021). Unlike the site-specific editing events in coding sequences, typically confined to short and imperfect duplex regions, the long transposable repeat-derived dsRNAs tend to be edited by ADAR1 promiscuously, producing clusters of inosines that are referred to as hyper-editing (Porath et al. 2014). These clusters of editing events can markedly alter the structure of cellular dsRNAs by replacing canonical A-U base pairs with the less stable I-U wobbles and can transform dsRNAs–protein interactions, including the recognition by dsRNA sensors (Fig. 2B; Bass and Weintraub 1988; Strobel et al. 1994; Serra et al. 2004; Wright et al. 2007; Ahmad et al. 2018).
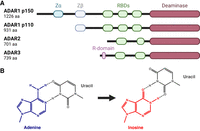
ADAR proteins and RNA editing. (A) Human ADAR proteins. (RBD) RNA-binding domain. (B) Deamination of adenine to inosine changes the base pair geometry and lowers the stability of base-pairing.
Our understanding of the physiological roles of ADAR1 has undergone a significant shift since the original discovery that ADAR1 was essential for survival. Initially centered on the biological functions of individual editing events, more recent research has established a global role of ADAR1 in regulating the cellular “dsRNAome.” Genetic studies of mouse models and human disease have identified ADAR1 as a pivotal regulator of dsRNA innate immunity, bearing implications for autoinflammatory diseases and cancer. Among the pathways responsible for sensing dsRNA, the predominant in vivo function of ADAR1 appears to be the antagonism of MDA5 through A-to-I editing. Nonetheless, recent findings highlight editing-independent mechanisms that also play a crucial role in maintaining cellular homeostasis. These mechanisms involve the competition for dsRNA binding between ADAR1 and PKR, and between ADAR1 and ZBP1. In the following sections, we present the evidence supporting ADAR1's role as a suppressor of dsRNA immunity, with a primary focus on the ADAR1–dsRNA–MDA5 axis. We conclude with a discussion of the emergent promise of targeting ADAR1 in cancer immunotherapy.
THE ADAR1–dsRNA–MDA5 AXIS OF LONG dsRNA-MEDIATED IMMUNITY
Genetic evidence for the ADAR1–dsRNA–MDA5 interplay in mouse
Genetic studies in mice were instrumental in establishing ADAR1's functions in dsRNA immunity. Both ADAR1 and ADAR2 are essential for survival in mice (Higuchi et al. 2000; Hartner et al. 2004; Wang et al. 2004). In the case of ADAR2, a single recoding event in the neurotransmitter receptor GRIA2 accounts for the lethal deletion phenotype (Higuchi et al. 2000). In contrast, the mechanism behind the lethal phenotype of ADAR1 deletion proved more complex and more challenging to delineate.
The knockout of ADAR1 in mice leads to embryonic lethality due to extensive liver damage (Hartner et al. 2004; Wang et al. 2004). At the molecular level, ADAR1-deficient mouse embryos display hallmarks of an activated innate immune response, characterized by excessive production of interferon and the induction of interferon-stimulated genes (ISGs) (Hartner et al. 2009). These observations strongly hinted at ADAR1's involvement in controlling innate immunity.
Nevertheless, the early attempts to identify the responsible molecular pathways were unsuccessful. Knockout of PKR, a dsRNA sensor that induces translational arrest and apoptosis, failed to rescue the lethal Adar1 null phenotype (Wang et al. 2004). The OAS-RNase L pathway, which likewise is triggered by cytoplasmic dsRNAs, also appeared to be unaffected in vivo (Wang et al. 2004). It was not until a decade later that the question of ADAR1's role in innate immunity was revisited with new mouse models, including mice lacking components of the long dsRNA-sensing MDA5–MAVS pathway (Mannion et al. 2014; Liddicoat et al. 2015, 2016; Pestal et al. 2015; Heraud-Farlow et al. 2017; Li et al. 2017; Costa Cruz et al. 2020). MDA5 had not yet been linked to dsRNA innate immunity at the time of the initial rescue studies (Andrejeva et al. 2004; Yoneyama et al. 2004). Yet, of all tested cytoplasmic nucleic acid sensors and interferon pathway components, only the knockout of the dsRNA sensor MDA5 or its downstream effector MAVS rescued embryonic death in mice lacking functional ADAR1 (Mannion et al. 2014; Liddicoat et al. 2015; Pestal et al. 2015; Heraud-Farlow et al. 2017; Li et al. 2017; Costa Cruz et al. 2020). Removal of the dsRNA sensor RIG-I, DNA sensing adaptor STING, interferon receptors (IFNαR, IFNγR), or the transcriptional activator of interferon signaling STAT1 provided no rescue (Mannion et al. 2014; Pestal et al. 2015; Liddicoat et al. 2016).
Mechanistically, it was the A-to-I editing activity of ADAR1 that obstructed MDA5 activation and the induction of ISGs. This was evidenced by the fact that a catalytically inactive ADAR1 point mutant produced an embryonically lethal phenotype similar to that observed in full ADAR1 knockout (Liddicoat et al. 2015). Notably, mice lacking the A-to-I editing function of ADAR1 were able to reach their full life span when MDA5 was eliminated, indicating tight coupling between ADAR1-mediated editing and dsRNA sensing by MDA5 (Liddicoat et al. 2015; Heraud-Farlow et al. 2017). Consistent with a role in suppressing cytoplasmic dsRNA sensing, the removal of ADAR1's cytoplasmic, interferon-inducible p150 isoform alone was sufficient to replicate the Adar1 null phenotype (Ward et al. 2011; Pestal et al. 2015). Collectively, these genetic studies pinpointed MDA5–MAVS as the primary pathway that is negatively regulated by ADAR1-mediated editing in vivo. This conclusion was later corroborated by a mouse model completely devoid of A-to-I editing by ADAR1 and ADAR2 (Chalk et al. 2019). A quadruple mutant mouse was generated by breeding ADAR1 editing-deficient mice that were rescued by MDA5 knockout with ADAR2-deficient mice that were rescued by an A-to-G substitution (mimicking A-to-I) in ADAR2's essential target GRIA2. The completely editing-deficient mice have normal life spans without apparent phenotypes, indicating that editing is only vitally important within MDA5's dsRNA targets and the GRIA2 mRNA. Together, the genetic studies in mouse models have established an essential function of ADAR1-mediated editing in suppressing spontaneous MDA5 activation by endogenous dsRNAs.
Clinical evidence for the ADAR1–dsRNA–MDA5 interplay in humans
The ADAR1–dsRNA–MDA5 axis is conserved from mouse to human, as revealed by genetic studies of human immune disorders. ADAR1 loss-of-function (LOF) and MDA5 gain-of-function (GOF) mutations give rise to an array of rare autoinflammatory diseases (i.e., conditions characterized by an overactive innate immune system) (Rice et al. 2012, 2014, 2020; Hayashi and Suzuki 2013; Oda et al. 2014; Crow et al. 2015; Crow and Stetson 2022). These so-called type-I interferonopathies range from relatively mild skin conditions in individuals with one LOF ADAR1 allele to lethal neurodevelopmental disorders in the presence of MDA5 GOF mutations or two LOF ADAR1 alleles (Crow et al. 2015; Rice et al. 2020). Despite differences in symptoms, all tested patients exhibit up-regulation of type-I interferon activity, mirroring the constitutive interferon signaling observed in ADAR1-deficient mice.
Both ADAR1 and MDA5 mutations have been linked to the rare neurodevelopmental disease Aicardi–Goutières syndrome (AGS), which can also be caused by mutations in other genes involved in nucleic acid sensing (Crow et al. 2015). AGS typically starts in childhood, manifesting with brain and skin abnormalities and often severe intellectual and physical disability (Crow and Manel 2015). Most AGS patients with ADAR1 mutations contain two mutated alleles, with at least one mutation in the deaminase domain, whereas the most common compound mutation is located in the p150 isoform-specific Zα domain (Rice et al. 2012). All disease-linked ADAR1 mutations are predicted to decrease the deaminase activity or weaken dsRNA binding (Rice et al. 2012; Matthews et al. 2016; Karki et al. 2024). In contrast, the MDA5 mutations behind AGS and other autoinflammatory disorders confer gain of function by stabilizing MDA5 filament formation on dsRNA (Rice et al. 2014, 2020; Yu et al. 2021). These pathogenic mutations typically cluster around the ATPase active site, disrupting the ATPase activity and consequently slowing the dissociation of MDA5 monomers from dsRNA (Peisley et al. 2012; Rice et al. 2014; Yu et al. 2018). The resulting stabilization of dsRNA binding makes MDA5 less sensitive to imperfections in dsRNA structure and A-to-I editing, leading to constitutive MDA5 activation by endogenous dsRNAs (Ahmad et al. 2018; Yu et al. 2018). Conversely, MDA5 LOF variants can be protective against certain autoimmune diseases, including type-I diabetes, psoriasis, psoriatic arthritis, vitiligo, and coronary artery disease, although excessive MDA5 LOF increases the susceptibility to certain viral infections (Nejentsev et al. 2009; Shigemoto et al. 2009; Li et al. 2010; Asgari et al. 2017; Budu-Aggrey et al. 2017; Dand et al. 2017; Jin et al. 2017; Lamborn et al. 2017; Zaki et al. 2017; Emdin et al. 2018; Cananzi et al. 2021; Chen et al. 2021a).
The clinical manifestation of ADAR1 and MDA5 variants, when considered alongside mouse genetic studies, suggests a precisely tuned balance between MDA5's sensing of endogenous dsRNAs and the counteracting editing activity of ADAR1 (Fig. 3). Disruption of this delicate balance through perturbations in any component—be it ADAR1 loss, heightened expression of endogenous dsRNA, or MDA5 gain of function—can escalate dsRNA sensing by MDA5, leading to autoinflammation. Adjusting each of the three components of the ADAR1–dsRNA–MDA5 axis holds the potential not only to address autoinflammatory diseases but also transform treatments that benefit from transient interferon induction, as epitomized by the emerging applications in cancer immunotherapy.
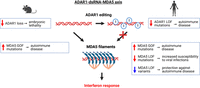
The ADAR1–dsRNA–MDA5 axis of innate immunity. In their unedited form, long cytoplasmic dsRNAs serve as substrates for MDA5 filament formation, leading to the induction of interferon response. By catalyzing A-to-I editing in endogenous dsRNAs, ADAR1 prevents MDA5 oligomerization and constitutive interferon activation. Boxes indicate the physiological consequences of genetic perturbations of ADAR1 and MDA5 activity in mice and humans.
Mapping immunogenic dsRNAs
The immunogenic “self” dsRNAs represent the most enigmatic and likely most complex component of the ADAR1–dsRNA–MDA5 axis. Initiating the MDA5–MAVS dsRNA-sensing pathway requires the formation of stable MDA5 filaments on long dsRNAs, typically comprising at least several hundred base pairs (Fig. 1; Cadena and Hur 2019). Interactions between adjacent dsRNA-bound MDA5 monomers stabilize dsRNA binding, counteracting the rapid ATP-dependent dissociation of individual monomers (Cadena and Hur 2019). The MDA5 filaments activate the downstream effector MAVS by bringing together MDA5's amino-terminal caspase activation and recruitment domains to form oligomers, which in turn nucleate filaments of MAVS CARD domains (Fig. 1; Wu and Hur 2015). Once in its active filamentous state, MAVS recruits TRAF molecules, triggering the interferon signaling cascade (Wu and Hur 2015).
MDA5 filaments are best sustained by long perfect dsRNAs, such as those arising from viral replication (Peisley et al. 2012). Nevertheless, the genetic studies discussed above demonstrated that cellular dsRNAs can sufficiently activate MDA5–MAVS to trigger lethal autoinflammatory responses, unless these dsRNAs are “neutralized” by ADAR1-mediated editing (Fig. 3). The vast majority of the endogenous dsRNAs are presumably formed by inverted repeats where most of ADAR1-mediated hyper-editing takes place (Tan et al. 2017; Reich and Bass 2019; Levanon et al. 2023). Most of these repeats are located in introns and intergenic regions, making them unlikely to be exposed to cytoplasmic dsRNA sensors like MDA5. Nevertheless, a small fraction of tandem inverted repeats occur in mature mRNA, particularly in the untranslated regions (UTRs), and are exported into the cytosol, where they become potential MDA5 ligands capable of inducing an innate immune response. Through A-to-I editing, the cytoplasmic ADAR1 p150 isoform destabilizes and reshapes the dsRNA structure, interfering with MDA5 filament formation and thereby mitigating the risk of inflammation (Fig. 3; Strobel et al. 1994; Serra et al. 2004; Wright et al. 2007; Ahmad et al. 2018).
The precise identity of cellular dsRNAs that require A-to-I editing to avoid MDA5 sensing has been a topic of extensive investigation and debate (Chen and Hur 2022; Levanon et al. 2023; Cottrell et al. 2024). One strategy for identifying potentially immunogenic cytoplasmic dsRNAs involves isolating the editing sites that are selectively introduced by the cytosolic, interferon-inducible ADAR1 p150 isoform, in contrast to nuclear ADARs (Kim et al. 2021; Sun et al. 2021, 2022; Kleinova et al. 2023). Studies using this approach have consistently highlighted an enrichment for UTR-localized inverted Alu repeats (IR Alus) among p150-selective sites and suggest that only a minuscule fraction of IR Alus may be immunogenic (Kim et al. 2021; Sun et al. 2022; Levanon et al. 2023). Intriguingly, some of the most potent immunogenic dsRNAs may not derive from Alu repeats at all but instead originate from cis-natural antisense transcripts (cis-NATs)—that is, complementary RNAs that are transcribed in opposite directions (Faghihi and Wahlestedt 2009; Li et al. 2022; Sun et al. 2022). Unlike IR Alus, which form relatively short, ∼300-bp imperfect duplexes, cis-NATs constitute perfectly complementary dsRNAs spanning hundreds to over 1000 bp, making them much more suitable ligands for stable MDA5 multimerization (Sun et al. 2022; Levanon et al. 2023; Cottrell et al. 2024). Although cis-NATs are rare compared to IR Alus, their ideal properties as MDA5 substrates suggest that they may carry disproportionate weight in the pool of immunogenic dsRNAs. Indeed, cis-NAT editing was implicated in a number of autoinflammatory and immune-related diseases based on their enrichment at the genomic loci defined by genome-wide association studies (Li et al. 2022). Moreover, cis-NATs but not the primate-specific IR Alus tend to be conserved from mice to humans and may have played a pivotal role in the conservation of the ADAR1–dsRNA–MDA5 axis (Sun et al. 2022).
Immunogenic dsRNAs comprise a dynamic category of cellular dsRNAs that is shaped by tissue-specific transcriptional programs, among other factors. For instance, neural progenitor cells (NPCs) stand out for their elevated burden of immunogenic dsRNA compared to other cell types (Chung et al. 2018; Sun et al. 2022; Dorrity et al. 2023). This increased burden results both from the distinct identities of RNAs expressed in NPCs and from cell-type specific variations in RNA expression levels. Moreover, the global lengthening of the 3′ UTRs of neuronal mRNAs contributes to the heightened dsRNA load in neurons (Dorrity et al. 2023). The overabundance of immunogenic dsRNAs in neuronal cells may explain why in the ADAR1-linked genetic disorder AGS, inflammation is largely confined to the brain. The cell-type specific immunogenic dsRNA burden may also account for inflammation patterns in other diseases, such as type-I diabetes (Knebel et al. 2024). This subject warrants further exploration across the spectrum of autoimmune and related conditions.
BEYOND A-TO-I EDITING: THE INVOLVEMENT OF ADAR1 IN PKR AND ZBP1 PATHWAYS OF dsRNA IMMUNITY
Competition between ADAR1 and PKR for dsRNA binding
The essential role of ADAR1-mediated dsRNA editing in mitigating MDA5-induced dsRNA autoimmunity is well-established. However, mouse and in vitro studies have also unveiled that ADAR1 juggles several functions across dsRNA-sensing pathways, some of which may not require the deaminase function at all.
In the landmark genetic studies in mice that elucidated the ADAR1–dsRNA–MDA5 axis, an initially puzzling observation arose from rescue experiments of mice with full ADAR1 knockout versus those with a deaminase-inactivating mutation. Concurrent MDA5 knockout provided full rescue only in mice expressing editing-deficient ADAR1 but not in those entirely lacking ADAR1 (Pestal et al. 2015). The latter were still rescued from embryonic lethality but only survived for two days postbirth. Knockout of MDA5's downstream adaptor MAVS yielded analogous partial rescue of Adar1 null mice (Mannion et al. 2014; Pestal et al. 2015). This implied that other, editing-independent functions of ADAR1 become important during later development (Mannion et al. 2014; Pestal et al. 2015). Two studies published in 2023 finally identified the dsRNA sensor PKR as the missing link, by demonstrating that knocking out PKR in addition to MDA5 (or its downstream adaptor MAVS) rescued ADAR1 knockout mice to adulthood (Hu et al. 2023; Sinigaglia et al. 2023). Previously, PKR knockout alone was known to be ineffective in rescuing the embryonic lethal phenotype of ADAR1 knockout, despite abundant evidence for ADAR1 suppressing PKR activation in vitro (Wang et al. 2004; Toth et al. 2009; Li et al. 2017; Chung et al. 2018; Corbet et al. 2021). The new in vivo studies reconcile these seemingly conflicting observations. The accompanying mechanistic studies revealed that ADAR1 operates through two distinct mechanisms: it prevents MDA5 activation through A-to-I editing of immunogenic dsRNAs, and it competes with PKR for dsRNA binding, thereby preventing PKR-induced translational arrest (Hu et al. 2023). Both pathways must be suppressed by ADAR1 to prevent fatal inflammation in mice, explaining why disabling MDA5/MAVS and PKR individually was insufficient to fully compensate for ADAR1 loss in earlier mouse studies.
From structural and biochemical perspectives, PKR and MDA5 form distinct interactions with dsRNA, contributing to the different modes of ADAR1 intervention. Whereas MDA5 requires long dsRNAs spanning hundreds of base pairs to form stable filaments, PKR's active form is a dsRNA-induced dimer, requiring as few as 33 bp of dsRNA (Hull and Bevilacqua 2016). Once dimerized, PKR undergoes autophosphorylation, generating the active pPKR form that phosphorylates the translation initiation factor eIF2α, leading to global translational shutdown and ultimately apoptosis (Hull and Bevilacqua 2016). Besides differences in the dsRNA footprint and downstream steps, PKR and MDA5 utilize different domains for dsRNA binding. Whereas MDA5 engages dsRNA through its helicase domains, PKR contains two dsRNA-binding domains (dsRBDs). dsRBDs represent a conserved class of RNA-binding motifs and are also present in ADAR proteins (Fig. 2A). In the case of ADAR1, three dsRBDs are located upstream of its deaminase domain. The shared cytoplasmic localization and similar RNA-binding domain composition can explain the ability of ADAR1 p150 to efficiently compete with PKR for binding cytosolic dsRNAs (Cottrell et al. 2024). Indeed, overexpression of ADAR1's dsRBD domains alone was sufficient to suppress PKR activation, as was cytoplasmic expression of dsRBDs from other dsRNA-binding proteins (Hu et al. 2023).
Although PKR's dsRNA targets may still be edited by ADAR1, the mismatches potentially introduced by A-to-I editing seem to have a much smaller role in preventing the binding of PKR compared to blocking MDA5 filaments. This observation can be rationalized by PKR only requiring short continuous dsRNA regions, which will still be present in most edited dsRNAs. On the other hand, even sparse mismatches introduced by ADAR1 may interfere with stable multimerization of MDA5 on long dsRNA duplexes. The distinct dsRNA length requirements of PKR and MDA5 also imply that the sets of dsRNA recognized by each sensor are not identical, even though for both IR Alus comprise a major subset of targets (Ahmad et al. 2018; Kim et al. 2018; Sun et al. 2022).
Competition between ADAR1 and ZBP1 and the roles of ADAR1's Zα domain in innate immunity
ADAR1 p150 stands out among mammalian ADARs in several ways: its cytoplasmic localization, expression from an IFN-inducible promoter, and the possession of a unique amino-terminal Zα domain (Fig. 2A). As discussed earlier, the former two features clearly position ADAR1 p150 to interact with cytosolic dsRNA-sensing pathways. Recent investigations now show that the third distinguishing feature, a Z-form RNA and DNA binding domain, also contributes to the role of ADAR1 p150 in curbing dsRNA immunity.
Although the majority of AGS-causing ADAR1 mutations reside in the deaminase domain, surprisingly, the most common ADAR1 compound mutation in AGS patients, Pro193Ala, is located in the Zα domain (Crow and Manel 2015). Found in only one other human protein, ZBP1, this domain recognizes the Z-conformation of dsDNA and dsRNA, which can be adopted by purine–pyrimidine repeat sequences (Herbert et al. 1997; Schwartz et al. 1999; Brown et al. 2000; Placido et al. 2007). Consistent with Z-RNA binding by ADAR1, purine–pyrimidine tracts can accelerate the editing of RNA substrates in vitro and may enhance the editing of certain substrates in vivo (Koeris et al. 2005). In a crystal structure of the ADAR1 Zα domain complexed with Z-form DNA, Pro193 (the residue mutated in AGS) contacts the Z-DNA backbone, suggesting that the alanine substitution may weaken the binding of certain dsRNAs; alternatively, the structural role of Pro193 may result in misfolding of the Zα domain upon mutation (Schwartz et al. 1999; de Reuver et al. 2021). The importance of Pro193 for RNA binding and editing is further supported by the decreased editing of a representative substrate in HEK293 cells expressing the Pro193Ala mutant (Mannion et al. 2014).
Several studies recently zeroed in on the role of the Zα domain in innate immune regulation in vivo (de Reuver et al. 2021, 2022; Maurano et al. 2021; Nakahama et al. 2021; Tang et al. 2021; Jiao et al. 2022). Mice with mutations in the Zα domain typically exhibit mild or no defects and show subtle changes in editing (de Reuver et al. 2021; Maurano et al. 2021; Nakahama et al. 2021; Tang et al. 2021; Liang et al. 2023). Severe detrimental effects are observed only when the AGS-linked Pro193Ala mutation is introduced in the absence of a second Adar1 p150 allele, resulting in a shortened life span for a subset of animals (Maurano et al. 2021; Liang et al. 2023). In all mouse models, the observed interferon up-regulation was MDA5–MAVS dependent, with additional involvement of the dsRNA sensors PKR and LGP2. Thus, the Zα domain exerts a modest effect on ADAR1 p150's ability to modulate dsRNA immunity, but this role may be exacerbated in certain contexts.
ADAR1 p150 shares the Zα domain with only one other mammalian protein, ZBP1, which is likewise implicated in innate immunity through as yet incompletely understood mechanisms (de Reuver and Maelfait 2023). The potential for competition between the two proteins for binding the same Z-form prone RNA targets spawned a collection of recent studies focusing on ZBP1 (de Reuver et al. 2022; Hubbard et al. 2022; Jiao et al. 2022). These investigations uncovered the involvement of ZBP1 in triggering the interferon response in ADAR1-deficient mice and found that ZBP1 knockout could extend the survival of ADAR1 knockout mice, although to much shorter life spans than seen in the complete rescue by MDA5 and PKR knockout (de Reuver et al. 2022; Hubbard et al. 2022; Jiao et al. 2022; Hu et al. 2023; Sinigaglia et al. 2023). These studies paint a nuanced picture of the importance of ADAR1's Zα domain while providing support for a model of competition between ADAR1 p150 and ZBP1 (de Reuver and Maelfait 2023).
Taken together, the interactions of ADAR1 with the MDA5, PKR, and ZBP1 dsRNA-sensing pathways demonstrate how ADAR1 utilizes its versatile domain architecture to safeguard the cell from dsRNA autoimmunity on multiple fronts. The A-to-I editing activity of the deaminase domain is responsible for preventing MDA5 multimerization, the dsRBDs compete with PKR for dsRNA binding; and the Zα domain confers the unique ability to compete with ZBP1 for binding of select dsRNAs. This spectrum of activities by ADAR1 epitomizes the intricate competitive landscape of endogenous dsRNAs and their interaction partners (Cottrell et al. 2024). With the key players and pathways now identified, a deeper mechanistic understanding of the involved pathways will be a key priority, along with therapeutic development guided by such understanding.
ADAR1 IN CANCER
Evidence for ADAR1's role as a checkpoint of anticancer immunity
The neutralizing role of ADAR1 in long dsRNA-mediated immunity can be exploited by cancer cells to promote tumor growth and evade immune recognition (Bhate et al. 2019). Here, we briefly discuss the potential for targeting ADAR1 in cancer immunotherapy and highlight the promise of activating the MDA5, PKR, and/or ZBP1 dsRNA-sensing pathways through ADAR1 inhibition.
Targeting the innate immune system presents a potent antitumor strategy, as has been successfully demonstrated for the cGAS-STING and RIG-I pathways, which run parallel to the MDA5 pathway (Fig. 1; Kasumba and Grandvaux 2019; Reisländer et al. 2020; Cao et al. 2022). In preclinical settings, durable tumor regression can be achieved by treating tumors with synthetic cGAS-STING and RIG-I agonists in combination with immune checkpoint inhibitors or, in some cases, as standalone treatments (Kasumba and Grandvaux 2019; Reisländer et al. 2020). The analogous promise of activating MDA5 to combat cancer is supported by a variety of experimental evidence. Similarly to RIG-I and STING agonists, synthetic MDA5 ligand Poly(IC:LC)—a dsRNA derivative—enhances the potency of a cancer vaccine (Kasumba and Grandvaux 2019). Moreover, the efficacy of epigenetic cancer therapy was attributed to MDA5 activation by endogenous dsRNAs that become up-regulated in cancer cells upon treatment with DNA methyltransferase inhibitors or inhibitors of other epigenetic processes (Chiappinelli et al. 2015; Roulois et al. 2015; Brocks et al. 2017; Cuellar et al. 2017; Sheng et al. 2018; Zhang et al. 2018; Mehdipour et al. 2020; for review, see Chen et al. 2021b). Similarly, treating mouse tumors with spliceosome-targeted therapies leads to cytoplasmic accumulation of misspliced dsRNAs and activation of dsRNA innate immune pathways, including MDA5 (Bowling et al. 2021). Despite the clear therapeutic potential of MDA5 activation in cancer, specific MDA5 agonists without severe toxicities are yet to be identified (Kasumba and Grandvaux 2019). Potentially circumventing this challenge, the ADAR1–MDA5 antagonism provides a unique opportunity to indirectly activate MDA5 through ADAR1 inhibition.
ADAR1 is overexpressed in various cancers, and its pro-oncogenic effects have been compellingly linked to the silencing of endogenous dsRNA-sensing pathways. In the pivotal studies that first identified ADAR1 as an immuno-oncology target, ADAR1 consistently emerged as a top suppressor of anticancer immunity (Manguso et al. 2017; Gannon et al. 2018; Ishizuka et al. 2019; Liu et al. 2019). Deletion of ADAR1 sensitized certain tumors to checkpoint inhibitor therapy, resulting in reduced tumor growth. This effect was associated with the activation of interferon signaling in cancer cells and the inflammation of the tumor microenvironment, dependent on the MDA5–MAVS axis (Gannon et al. 2018; Ishizuka et al. 2019; Liu et al. 2019). Furthermore, the ADAR1 knockout triggered tumor cell growth arrest and apoptosis through the PKR pathway (Gannon et al. 2018; Ishizuka et al. 2019; Liu et al. 2019). Consistent with our current understanding of ADAR1's distinct modes of interference with MDA5 and PKR sensing, ADAR1 impeded both pathways albeit through different mechanisms (Gannon et al. 2018). Although in both cases dependent on the cytoplasmic p150 isoform, MDA5 but not PKR suppression required editing (Gannon et al. 2018). In vivo, the MDA5 and the PKR pathways each were sufficient to prime Adar1 null tumors for immune checkpoint inhibitor (anti-PD1) blockade (Ishizuka et al. 2019). Thus, ADAR1-dependent suppression of the MDA5 and PKR dsRNA-sensing pathways is essential for tumor growth in a subset of cancers, making tumors vulnerable to ADAR1 loss and, prospectively, pharmacological ADAR1 inhibition (Fig. 4). In line with these results, ADAR1 knockdown also enhances the effects of epigenetic therapy, boosting MDA5 activation by DNMTi-induced endogenous dsRNA (Mehdipour et al. 2020). More recently, ADAR1's role in blocking ZBP1-dependent cell death was also implicated in cancer, indicating additional opportunities for intervention in the realm of cancer immunotherapy (Karki et al. 2021; Zhang et al. 2022; de Reuver and Maelfait 2023).
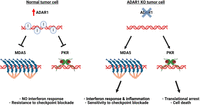
ADAR1 promotes tumor growth by suppressing dsRNA sensing. (Left) Increased ADAR1 expression in tumors (red arrow) leads to efficient editing of endogenous dsRNAs, including abnormal dsRNAs that may accumulate in cancer cells. Through editing dependent and independent mechanisms, ADAR1 inhibits spontaneous activation of MDA5 and PKR, thus promoting tumor growth and preventing immune recognition, even in the presence of checkpoint inhibitors. (Right) ADAR1 deletion unleashes the innate immune response against cellular dsRNAs in cancer cells, leading to inflammation and enhanced sensitivity to checkpoint inhibitors (via the MDA5 pathway) and triggering apoptosis (via the PKR pathway).
Toward ADAR1-based cancer therapies
Taken together, accumulating evidence suggests that perturbations in the core components of the ADAR1–dsRNA–MDA5 axis, along with parallel pathways involving PKR and ZBP1, could be leveraged for therapeutic benefit in cancer. A crucial consideration is establishing tolerance levels for ADAR1 inhibition or activation of MDA5, PKR, and ZBP1 in patients. The severity of certain ADAR1 and MDA5-linked autoinflammatory diseases warrants caution, as does the potential for cell death beyond the tumor. Nonetheless, the elevated ADAR1 expression observed in some cancers suggests a therapeutic window for the specific activation of dsRNA sensors in tumors.
The recent mechanistic insights offer many opportunities for fine-tuning the potency and specificity of ADAR1's antitumor effects. For example, selectively inhibiting the deaminase activity should induce MDA5-dependent interferon up-regulation without triggering PKR-dependent apoptosis or ZBP1-dependent cell death. Meanwhile, targeting the RNA-binding activity of ADAR1 would likely induce several dsRNA-sensing pathways, including PKR. Each of these strategies may be advantageous in different scenarios.
Research on ADAR1's roles in innate immunity and cancer alike underscores the presence of a pool of endogenous dsRNAs in mammalian cells that must be edited by ADAR1 to prevent constitutive MDA5 activation. A better understanding of the nature of these immunogenic dsRNAs may enable their manipulation as specific MDA5 agonists in cancer therapies and beyond. Epigenetic therapy seems to trigger innate immune signaling by increasing the expression of some immunogenic RNAs (Chen et al. 2021b). Alternative strategies to induce the expression of dsRNAs, for example, by inhibiting RNA methylation, or more targeted approaches may also help achieve therapeutically appropriate levels of MDA5 activation (Liu et al. 2020, 2021; Chelmicki et al. 2021).
CONCLUDING REMARKS AND FUTURE DIRECTIONS
Genetic studies in human patients and mouse models have firmly established the central role of ADAR1 in regulating dsRNA-mediated innate immunity. It is now clear that this same role is exploited by cancer cells to promote tumor growth and evade immune detection. Promisingly, targeted inhibition of ADAR1 in tumors, once therapeutically possible, may provide a more specific and safer approach to triggering an interferon response in tumors than, for example, global DNA demethylation in epigenetic therapy. To harness the full potential of ADAR1 and dsRNA sensing as therapeutic targets in cancer, several key questions need to be addressed.
First, specific ADAR1 inhibitors and MDA5 or PKR agonists are yet to be identified. Progress in this area will depend on advances in structural and mechanistic understanding of ADAR1 and the innate immunity pathways it interacts with. In particular, the efforts to inhibit ADAR1 would greatly benefit from high-resolution structural data. At a broader level, determining the most effective types of perturbations (ADAR1 repression, activation of dsRNA sensors, or increasing the tumor's dsRNA burden), or combinations thereof, is essential to achieve potent antitumor effects with minimal toxicity.
Additionally, the design of specific MDA5 agonists would benefit from an improved understanding of the identities and features of cellular immunogenic dsRNAs. Despite recent advances, studies of cellular dsRNAs and their immunogenic potential remain an active research frontier. Intriguingly, rather than comprising a defined set of RNAs, the cellular repertoire of immunogenic dsRNAs appears to be dynamic and vary between tissues, developmental stages, and disease contexts. This dynamic nature underscores the importance of studying immunogenic dsRNAs in physiologically relevant model systems that accurately reproduce in vivo complexity.
The surge in interest in ADAR1 biology promises a wealth of new insights in the coming years. We envision that the greatest advances will arise from the synthesis of continued fundamental studies with new ventures into clinical applications, together paving the way for ADAR1-based therapies in cancer and beyond.
COMPETING INTEREST STATEMENT
J.B.L. is a cofounder of AIRNA and a consultant for Risen Pharma. I.J. is an employee of AIRNA Corporation.
ACKNOWLEDGMENTS
Work in the Li Lab is funded in part by National Institutes of Health grants R35GM144100, R01 GM102484, R01 GM124215, R01 MH115080, and R01AI182437. Figures were created with BioRender.com.
Footnotes
-
Article is online at http://www.rnajournal.org/cgi/doi/10.1261/rna.079953.124.
-
Freely available online through the RNA Open Access option.
This article, published in RNA, is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








