
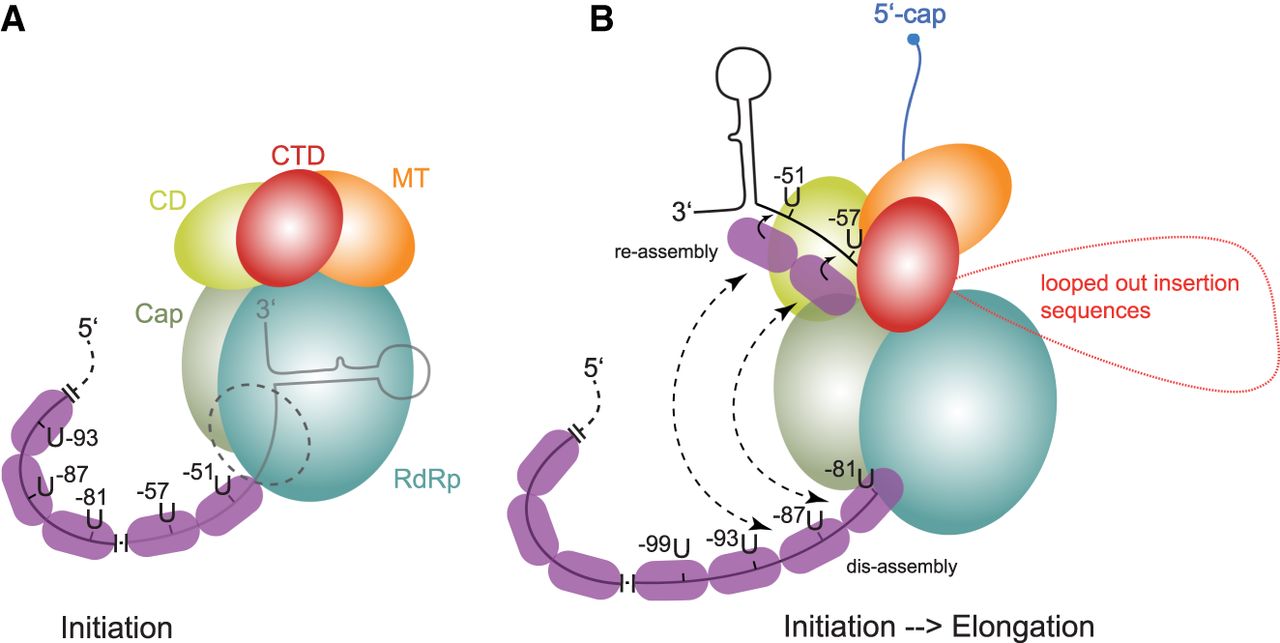
Models to explain the hexamer phasing, suggesting a key role for NP which binds 6 nt per monomer. The sketches are drawn according to the cryo-EM structure of the VSV polymerase (Liang et al. 2015). (A) This model assumes that there is essentially no replication or transcription activity (not even abortive transcription) of the EBOV polymerase when hexamer phasing is abolished. Here, the polymerase binds the ∼55 3′-terminal nucleotides (PE1) of the genome. Proper positioning of the enzyme on the template strand is only achieved when NP is assembled in the correct register, directed by the UN5 hexamers, explaining why the spacer can be expanded by multiples of 6. (B) This model, which is mechanistically largely speculative, may reflect the transition from the initiation to the elongation mode of the polymerase. It assumes that the enzyme starts to polymerize upon positioning of the genome 3′-end in the active site; when the genome 3′-end leaves the template exit channel, the enzyme may hold on to the first UN5 hexamers (U-51, U-57); as soon as UN5 hexamers enter the template entry path, they will be bound as well to trigger communication between the entry and exit path; hexamer periodicity may synchronize reassembly at the exit site and dis-assembly at the entry site; expansion sequences in the spacer region that may loop out are shown in red. The nascent mRNA transcript is shown in blue; in the VSV system, the nascent mRNA transcript is capped when reaching a length of 31 nt (Tekes et al. 2011). For more details, see text.








